
Abstract
The chloroplast (cp) genome sequence of Mussaenda pubescens, a promising resource that is used as a traditional medicine and drink, is important for understanding the phylogenetic relationships among the Mussaenda family and genetic improvement and reservation. This research represented the first comprehensive description of the morphological characteristics of M. pubescens, as well as an analysis of the complete cp genome and phylogenetic relationship. The results indicated a close relationship between M. pubescens and M. hirsutula based on the morphological characteristics of the flower and leaves. The cp was sequenced using the Illumina NovaSeq 6000 platform. The results indicated the cp genome of M. pubescens spanned a total length of 155,122 bp, including a pair of inverted repeats (IRA and IRB) with a length of 25,871 bp for each region, as well as a large single-copy (LSC) region and a small single-copy (SSC) region with lengths of 85,370 bp and 18,010 bp, respectively. The results of phylogenetic analyses demonstrated that species within the same genus displayed a tendency to group closely together. It was suggested that Antirhea, Cinchona, Mitragyna, Neolamarckia, and Uncaria might have experienced an early divergence. Furthermore, M. hirsutula showed a close genetic connection to M. pubescens, with the two species having partially overlapping distributions in China. This study presents crucial findings regarding the identification, evolution, and phylogenetic research on Mussaenda plants, specifically targeting M. pubescens.
Similar content being viewed by others


Introduction
The Mussaenda genus contributes significantly to medical natural products, including flavonoids, triterpenes, saponins, and iridoids identified primarily from M. pubescens. The stems and leaves of M. pubescens with a sweet flavor possess the therapeutic effects of cooling and relieving summer heat, clearing heat, and dispersing wind, commonly used medicinally or dried as a tea substitute1. The whole plant of M. pubescens has been used in Chinese folk medicine against laryngopharyngitis, acute gastroenteritis, and dysentery2. Its extracts exhibit immunostimulant, hemolytic3, detoxifying4, analgesic5, and anti-RSV activities6. The genus Mussaenda L., which belongs to the family Rubiaceae belonging to Rubiaceae, consists of approximately 120 species primarily found in Africa, the Pacific islands, and Asia, including 31 species, 1 variety, and 1 variant in China. The morphology of trichomes and the shape of calyx and corolla are valuable characteristics for species identification7. The species of Mussaenda can be easily distinguishable from other genera due to their expanded petaloid calycophylls, terminal umbel inflorescence, and fleshy berries. M. pubescens, a liana-like shrub belonging to the genus Musaenda8, is categorized into two forms known as M. pubescens f. pubescens and M. pubescens f. clematidiflora, which are identified as the same species through ‘the Key to Dioecious Species of Musaenda in China’.
In general, the chloroplast is a semi-autonomous replication organelle9 and contains independent cpDNA10. Some species exhibit a circular structure consisting of one short single-copy (SSC) region, one large single-copy (LSC) region, and two inverted repeats (IR) regions that serve as a separator between the LSC and SSC regions9. The contraction and expansion of the IR, LSC, and SSC regions are common phenomena in the process of evolution11, which regions constitute the primary factors that contribute to the differences in size observed in cp genomes12. SSRs identified in cp genomes exhibit high conservation in terms of types and numbers, and the majority of mono-nucleotide repeats consist of base A or T, making them particularly well-suited as genetic markers for plant molecular studies13. The cp genome offers valuable insights for various scientific fields such as species identification, population genetics, phylogenetics, and genetic engineering research due to its similar structure14,15, large genetic information, highly conserved sequences16, high-valuable information17, and stable maternal inheritance18. Therefore, the cp genome has become an ideal resource for species and higher-order phylogenetic inference19, serving as a molecular marker in taxonomic research20. The complete chloroplast genomes of various plant species within the same genus were sequenced using the Illumina NovaSeq 6000 platform, thus providing valuable insights for species identification, evolutionary analysis, and phylogenetic studies of these plants21.
In the present study, the complete chloroplast genomes of M. pubescens were sequenced using Illumina technology and subsequently characterized their features. Our research aimed to investigate the molecular structures of M. pubescens cp genomes and analyze the phylogenetic relationships among Mussaenda species by utilizing their complete cp genome sequences, including the LSC, SSC, and IR regions. The current research will serve as a valuable resource for future studies on species identification, evolution, population genetics, and phylogenetic analyses of Mussaenda.
Material and methods
Plant material and morphological observations
All the materials were collected from the Third Botanical Resources Survey in Libo County (Guizhou, China, 26° 12′ N, 107° 30′ E) in September 2020. The species has been officially identified as ‘M. pubescens’ by a professor of plant taxonomy at Qiannan Normal University for Nationalities, which would be compared with the specimens of Chinese Mussaenda. The specimen (DZ20210505) was deposited in the herbarium of Qiannan Normal University for Nationalities in Duyun City, Guizhou Province, China. Morphological features of M. pubescens involved in branches, leaves, stipules, inflorescences, calyx lobes, corolla tubes, florescences, and fruits were observed to compare their differences (Fig. 1).

Morphological characters of M. pubescens. (A) Growth environment; (B) Fruit; (C) The longitudinal section of the fruit; (D) Transection of the fruit; (E) A fruiting branch (2 cm); (F) Enlarged Calyxlobes (2 cm); (G) Stipule (1 cm); (H) A long-styly branch (Scale bar: 5 cm); (I) Calyxlobes (1 cm); (J) A flowering short-styly branch (2 cm); (K) long-styly stigma (1 cm); (L) Long-styly corolla (1 cm); (M) Long-styly flower (1 cm).
Complete chloroplast genome sequence of M. pubescens
The leaves of M. pubescens were collected on September 15, 2020 (Fig. 1) and stored immediately at − 80 °C. The isolation of total genomic DNA was performed using a modified CTAB method22. Library construction and sequencing were conducted by Novogene Bioinformatics Technology Co. Ltd. (Guangzhou, China) using the Illumina NovaSeq 6000 platform with a Paired-End 150 (PE150) strategy. The FastQC software was employed to trim low-quality reads and adapters, and the genome was de novo assembled using SPAdes v3.923, and subsequently annotated using Plann software22. The accuracy of the preliminary annotation results was verified by comparing them with the proteins and rRNA sequences from previously reported cp genomes of relevant species using the methods of blastn and blastp. The complete chloroplast genome was submitted to the Database Resources of the National Genomics Data Center, China National Center for Bioinformatio (Genome Sequence Archive accession number: CRA013306; https://ngdc.cncb.ac.cn/gsa). The circular structure of the cp genome was visualized using CPGView (http://www.1kmpg.cn/cpgview/).
Phylogenetic analysis
A total of 28 complete chloroplast genome sequences spanning across 24 Rubiaceae species were acquired from the NCBI GenBank to ascertain the phylogenetic positions of M. pubescens within the Rubiaceae lineages. The Bayesian inference (BI) tree was constructed following standard protocols. The alignment of 28 cp genomic sequences was generated using the MAFFT online version24 with default parameters. The software tools MAFFT, Ultrafast bootstrap, IQ-TREE, ModelFinder, and MrBayes were employed within the PhyloSuite framework25. FigTree was employed to illustrate the phylogenetic relationships (http://tree.bio.ed.ac.uk/software/figtree/).
Ethics approval and consent to participate
Mussaenda pubescens is not endangered in China, and no specific permission was required for the collection. All Mussaenda pubescens materials in this study were collected in germplasm resource nursery of the College of Biological Science and Agriculture, Qiannan Normal University for Nationalities (QNUN) with the permission of the school. The current study complied with relevant institutional, national, and international guidelines and legislation.
Results and discussion
Morphological characteristics of M. pubescens
The collected specimens of M. pubescens were compared with Chinese Mussaenda specimens for this research study (Table 1). The morphological features of M. pubescens, including branches, leaves, stipules, inflorescences, calyx lobes, corolla tubes, florescences, and fruits, were presented in Fig. 1.
Type: China. Guizhou, Libo County, Maolan National Nature Reserve, 22° 5′ N, 107° 56′ E, ca. 980 m a.s.l., on forest edge, 26 Sep 2020, S. Wang 161915 (Holotype, IBSC!).
Distribution and habitat: Currently, M. pubescens is known from the provinces of Zhejiang, Hainan, Yunnan, Guizhou, Sichuan, Guangxi, Guangdong, Hubei, Fujian, Hunan, Jiangxi, and Taiwan in China. This plant species is commonly found in dense thickets located in ravines, on slopes of hills, and along village boundaries or roadsides, typically thriving at elevations ranging from 100 to 900 m a.s.l.
Phenology: Flowering from April to July, and fruiting from June to November.
Description: M. pubescens, climbing shrub, often extensively twining. Branchlets appressed pubescence, the stems are often accompanied by axillary short shoots that bear small leaves. Leaves with distinctly petiolate (3–15 mm), were ovate-oblong or ovate-lanceolate (5–8 × 2–5 cm), opposite or perhaps rarely whorled, membranous or thinly papery, densely strigillose. The stipules were triangular, densely strigillose, deeply 2-lobed, segments subulate (4–6 mm long). Inflorescences were cyme with sessile to subsessile (0.1–1.4 cm, though the fruit may be stipitate), primarily unbranched, densely strigillose to villosulous. Calyx lobes were non-leaflike, linear or triangular, less than 1.5 mm wide, longer than corolla tube twice, densely pilose on base, becoming sparser upward; 1 lobe on 1–3 flowers per inflorescence expanded into calycophyll, broadly elliptic (2.5–5 × 2–3.5 cm), narrow on base, strigillose on both surfaces, with 5–7 longitudinal veins. Corolla tubes were gyro-shaped (3–4 mm long), uniformly cylindrical, or inflated just below or at the throat with dense clavate pubescence; corolla lobes were oblong-lanceolate (4 mm long) and acuminate. The flower period was Apr-Jul. Fruit period was Jun to Dec, were berry subglobose, 8–10 × 6–7.5 mm in diameter, smooth, fleshy or stiffly papery, sparsely pilose; stipitate 4–5 mm; black after drying.
Similar species: The breeding system of M. pubescens is characterized by functional dioecy. A Key to Dioecious Species of Mussaenda in China is provided below. The forms of trichomes, as well as the morphology of the calyx and corolla, are important characteristics for identifying species7. M. pubescens is easily distinguished by its corolla tube 11–20 mm, with stems frequently having axillary short shoots containing small leaves26,27, flowers sessile to pedicellate and range in color from white to yellow, despite bearing morphological similarities to M. hirsutula28. Differences among these two species are provided in Table 1.
Etymology: The specific epithet is derived from the shape of the corolla.
Paratypes: China, Guangxi: Liu Jun 1235399 (PPBC), Zhang Dianxiang 0153214 (IBSC), Deng Liang 0659417 (IBSC), Rong County Census Team 0205196 (GXM); Guangdong: Li Guangmin 149045 (PPBC), Deng Xiaotong 012602 (SN), Liu Na 012311 (SN), Yang Yuying 010881 (SN); Hunan: Yu Xunlin 1236160 (PPBC), Ran Jing 0007240 (GNUG); Yunnan: Chen Yousheng 449219 (PPBC), Li Yongliang 1450785 (KUN), Yu Xunlin 064698 (CSFI); Fujian: Qu Hua 068014 (AU), Ching H.H. 0659418 (IBSC), Guo Zhizhao 076512 (AU); Jiangxi: Cao Lan 0001459 (JXCM), Chen Gongxi 00197444 (SYS); Gui zhou: Li Qirui 0101386 (GZTM), Chen Jianxiang 0099290 (GZTM), Wu Shiyan 0102818 (GZTM); Taiwan: Masakazu 0768530 (IBSC), Bartholomew B. 00793088 (PE); Hong Kong: Shiu Ying 00793108 (PE), Chen Huanyong 00793121 (PE); Sichuan: Zhu Dahai 02115800 (PE), Fang Wenpei 0472099 (IBSC); Hubei: Fu Guoxun 0472100 (IBSC); Zhejiang: Qin Renchang 0472098 (IBSC).
Key to dioecious species of Mussaenda in China
-
1a. Individual flowers with all calyx lobes enlarged into petaloid calycophylls……………M. anomala
-
1b. Individual flowers with only 1 calyx lobes enlarged into petaloid calycophylls, or without calycophylls………(2)
-
2a. Normal calyx lobes subleaflike, lanceolate, 1.5–5 mm wide…………………(3)
-
3a. Branches, calyx, and corolla, covered villosulous………………M. macrophylla
-
3b. Branches, calyx and corolla, appressed strigillose …………………M. esquirolii
-
2b. Normal calyx lobes, non-leaflike, linear or triangular, less than 1.5 mm wide……………(4)
-
4a. Leaves, sessile or subsessile, < 3 mm
-
4b. Leaves, distinctly petiolate, 3–15 mm………………………(5)
-
5a. Normal calyx lobes (at anthesis) 1–15 mm, shorter than calyx tube
-
5b. Normal calyx lobes (at anthesis) 25–30 mm, longer than calyx tube………………(6)
-
6a. Calyx lobes longer than calyx tube, but not more than twice………………(7)
-
7a. Cymes, dense
-
7b. Cymes, Loose…………………………(8)
-
8a. Calyx lobes, linear
-
8b. Calyx lobes, subulate………………………(9)
-
9a. Corolla lobes, triangular-ovate; leaves, elliptic………………M. elliptica
-
9b. Corolla lobes, ovate; leaves, elliptic or ovate-elliptic, with pubescent on abaxially……M. divaricata
-
6b. Calyx lobes longer than calyx tube, more than twice and more………………(10)
-
10a. Calyx lobes longer than calyx tube, more than twice
-
10b. Calyx lobes longer than corolla tube, twice …………………(11)
-
11a. Corolla tube, elliptic; leaves, elliptic or oblong, rarely obovate……………(12)
-
12a. Corolla lobes, triangular-ovate; calyx lobes, linear-lanceolate…………M. hainanensis
-
12b. Corolla lobes, elliptic; calyx lobes, linear ………………M. hirsutula
-
11b. Corolla tube, gyro-shaped, 3–4 mm long; leaves, ovate-oblong or ovate-lanceolate; corolla lobes, oblong-lanceolate, acuminate ……………………M. pubescens
Chloroplast genome features of M. pubescens
Chloroplast DNA sequences, characterized by their highly conserved structure, minimal recombination events, and predominantly uniparental inheritance, have traditionally served as preferred markers for reconstructing plant phylogeny. A robust phylogeny is essential to investigate the evolutionary patterns of traits across varying taxonomic levels. The chloroplast genome yields essential information for various scientific disciplines, including species identification, population genetics, phylogenetics, and genetic engineering research14,15,29. The filtering process obtained 20,928,581 paired-end reads from the Illumina NovaSeq platform, all meeting the required quality standards. The values for Q20 and Q30 were determined to be 97.53% and 92.98%, respectively. The complete cp genome sequence of M. pubescens was constructed de novo and subsequently submitted to the Database Resources of the National Genomics Data Center (GSA accession number: CRA013306).
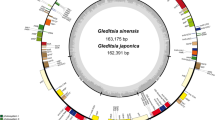
The chloroplast genomes of plants consisted of photosynthetic genes, genes related to chloroplast transcriptional expression, and additional protein-coding genes30. Expansion and contraction of the boundaries of the inverted region (IR) are the primary factors contributing to changes in the size of the cp genomes, playing a crucial role in species evolution31. As was common with other angiosperms, the cp genome of M. pubescens consisted of a circular genome measuring 155,122 bp in length and possessing the typical quadripartite structure. This structure included a pair of reverse repeats, namely IRA and IRB, which spanned 25,871 bp, a small single copy region (SSC) measuring 18,010 bp, and a large single copy region (LSC) spanning 85,370 bp (Table 2 and Fig. 2). The expansion and contraction observed in the IR regions might serve as the primary mechanism for generating variations in the length of the cp genomes in both M. pubescens and its closely related species9,32.

The chloroplast genome maps of M. pubescens. Genes situated on the inner side of the circle are transcribed in a clockwise direction, whereas those on the outer side are transcribed in a counter-clockwise direction. The inner circle of a darker gray represents the GC content, while the lighter gray indicates the AT content. Different colors represent different functional genes.
The GC content varied across different regions of the cp genomes, with the IR regions particularly exhibiting high GC content, likely due to the presence of rRNAs33. Our findings indicated that the GC contents in the IR and SSC regions were measured to be 43.17% and 31.89%, respectively, with the average GC content of the whole genome being 37.67% (Table 3). Notably, the GC content of DNA in the IR regions was higher than that in other regions (LSC, SSC) (Fig. 2), which aligned with similar patterns observed in other flowering plants34,35. Furthermore, the GC skewness has been identified as a crucial indicator of DNA leading chains, lagging chains, and replication origins and terminals, which in turn serves as an important determinant of species affinity36.
Gene functions were subsequently assigned to all the genes (Table 2), with these genes being classified into four types: genes related to self-replication, genes related to photosynthesis, unknown function genes, and specific genes, including maturase (matK), protease (clpP), and others. Out of the 128 identified genes, there were 86 protein-coding genes, 34 transfer RNA (tRNA) genes, and 8 rRNA genes. The observed results were similar to those found in other Mussaenda species37,38.
In total, 62 protein-coding genes and 22 tRNA genes were situated in the LSC region, while 11 protein-coding genes and 1 tRNA gene were assigned to the SSC region of the cp genome (Fig. 2). Additionally, a total of 20 intron-containing genes, among which there were 17 genes (e.g., ndhA, ndhB, petB, petD, atpF, rpl16, rpl2, and rps16) with 1 intron and only 3 genes (rps12, clpP1, and trnH-GUG) with 2 introns (Table 4). Among them, ndhA possessed the longest intron (1115 bp), while the shortest intron (16 bp) was observed in trnI GAU (Table 5). It was noteworthy that rps12 was classified as a trans-spliced gene consisting of two separated introns, with one exon positioned in the LSC region and the other two in the IR region (Fig. 2). Besides that, the rps12 gene of M. pubescens was comprised of an exon at the 5’-end located in the LSC region, while its 3’-end exons were positioned in the IR regions, as depicted in Fig. 2, which was consistent with that of the homologous species M. hirsutula38.
The codon usage bias in chloroplast genomes may arise from a combination of natural selection and genetic mutation, which is important to investigate this phenomenon as it provides insights into the evolutionary processes and functional constraints shaping the genetic code of chloroplasts39. The relative frequency of synonymous codons in the coding sequence of M. pubescens cp demonstrated that all genes were represented by 19,850 codons. The study identified the four most commonly utilized codons as ATT (Isoleucine), GAA (Glutamic acid), AAT (Asparagine), and AAA (Lysine), accounting for 843 (4.25%), 752 (3.79%), 723 (3.64%), and 715 (3.60%) codons, respectively. One of the most commonly used amino acids was leucine, with 2155 hits; another one, cysteine, had the lowest content, with only 277 hits. Additionally, codons ending with ‘A’ and ‘T’ accounted for 69.13% of all codons (Table 6), which aligned with previous studies on angiosperms40. The codon usage preferences encompassed within these features could actively contribute to a more in-depth understanding of exogenous gene expression and the mechanisms driving the evolution of the cp genome41,42.
Phylogenetic analysis
Chloroplast genomes are valuable sources of information for species identification and evolutionary analysis19. They serve as organelle-based “barcodes” to distinguish species and reveal interspecies phylogenetic relationships43. Furthermore, the progressive advancements in next-generation sequencing technology, specifically the implementation of second-generation technology, have facilitated the simplification of chloroplast genome sequencing. Thus, an accumulating body of studies has employed complete chloroplast genome sequences to examine phylogenetic relationships within angiosperms. The phylogenetic position of M. pubescens was analyzed by downloading 19 complete cp genomes from the GenBank database, all of which belonged to the Gramineae family. The genomes were aligned using the MAFFT24, and the phylogenetic tree was constructed with the Mega-X v10.0.5 software44 employing the maximum likelihood method and 1000 bootstrap replicates (Fig. 3). The observed cp genome sequences played a vital role in elucidating and comprehending the phylogenetic relationships among Mussaenda species.

The phylogenetic tree of M. pubescens.
The diversification of the species, M. pubescens, was attributed to the presence of a highly diverse set of genes, intermolecular recombination in the LSC region, and the co-occurrence of tandem repeats. The phylogenetic analyses yielded highly similar topologies across the complete cp genomes, LSC regions, and SSC regions (the complete cp genome displayed complete consistency with the LSC region), and every node in the phylogenetic trees exhibited high bootstrap support, with the exception of Antirhea chinensis and Cinchona officinalis (Fig. 3). Rubiaceae species, along with species in the same genus, showed a tendency to cluster together on a single large branch. The Rubiaceae branch was divided into two clades with Antirhea, Cinchona, Mitragyna, Neolamarckia, and Uncaria to other 20 genera. Mussaenda was found to be related to 19 other genera, and within these, it was determined that Mussaenda pubescens shared a close relationship with five other genera (Mussaenda hirsutula, Emmenopterys, Coffea, Diplospora, Lxora, and Trailliaedoxa). The phylogenetic tree demonstrated that Mussaenda was polyphyletic and originated from the intercalation of branches from the genera M. hirsutula. Moreover, M. pubescens and M. hirsutula were found to be closely related genera. The findings support the identification and taxonomic study of M. pubescens, enabling resource collection, cultivation, study of pharmacological activities, and development of functional products.
Conclusion
M. pubescens was compared with the specimens of Chinese Mussaenda, indicating that M. pubescens was closely related to M. hirsutula. However, M. pubescens was distinguished by its gyro-shaped corolla tubes measuring 3–4 mm in length, ovate oblong or ovate lanceolate leaves, and oblong lanceolate corolla lobes. Furthermore, this study determined the complete cp genomes of M. pubescens for the first time and revealed their basic structures, conservation, and variability. The complete cp genomes, including their LSC and SSC regions, were employed to investigate the robust phylogenetic relationships within and between genera. Additionally, the availability of this information would offer useful references for subsequent studies on taxonomic identification, phylogenetics, population structure, and biodiversity within the genus Mussaenda. Furthermore, the comparative analysis of cp genomics in the genus Mussaenda contributed to our comprehension of cp genome dynamics, complexity, and evolution within the Rubiaceae family. In conclusion, this study served as a valuable reference for future research on species identification, evolutionary relationships, and the development of genetic resources within the Rubiaceae family.
Data availability
The complete chloroplast genome has been submitted to the Database Resources of the National Genomics Data Center, China National Center for Bioinformatio (Genome Sequence Archive accession number: CRA013306; https://ngdc.cncb.ac.cn/gsa).
References
Li, D. et al. Ethnobotanical survey of herbal tea plants from the traditional markets in Chaoshan, China. J. Ethnopharmacol. 205, 195–206 (2017).
Chen, S., Luo, Z. & Zhang, D. Self-pollination in buds and homostyly in Mussaenda shikokiana (Rubiaceae), a monomorphic species in a distylous clade. J. Syst. Evol. 51, 731–742 (2013).
Xu, R., Zhao, W., Xu, J., Shao, B. & Qin, G. Studies on bioactive saponins from chinese medicinal plants. Adv. Exp. Med. Biol. 404, 371–382 (1996).
Xu, Y. et al. Effects of koumine, an alkaloid of Gelsemium elegans Benth., on inflammatory and neuropathic pain models and possible mechanism with allopregnanolone. Pharmacol. Biochem. Be. 101, 504–514 (2012).
Wang, Y., Wang, H., Wu, S., Li, D. & Chen, S. Effect of Gelsemium elegans and Mussaenda pubescens, the components of a detoxification herbal formula, on disturbance of the intestinal absorptions of indole alkaloids in Caco-2 cells. Evid.-Based Complement Altern. Med. 2017, 1–10 (2017).
Li, Y., Ooi, L. S. M., Wang, H., But, P. P. H. & Ooi, V. E. C. Antiviral activities of medicinal herbs traditionally used in southern mainland China. Phytother. Res. 18, 18–722 (2004).
Grecebio, J. D. A., Ulrich, M. & Sigrid, L. A taxonomic revision of Philippine Mussaenda (Rubiaceae, Mussaendeae). Ann. Mo. Bot. Gard. 101, 457–524 (2016).
Deng, X. & Zhang, D. Three new synonyms in Mussaenda (Rubiaceae) from China. J. Syst. Evol. 44, 608–611 (2006).
Xue, S. et al. Comparative analysis of the complete chloroplast genome among Prunus mume, P. armeniaca, and P. salicina. Hortic. Res. 6, 89 (2019).
Choi, K. S. & Park, S. The complete chloroplast genome sequence of Aster spathulifolius (Asteraceae); genomic features and relationship with asteraceae. Gene 572, 214–221 (2015).
Huang, H., Shi, C., Liu, Y., Mao, S. & Gao, L. Thirteen Camellia chloroplast genome sequences determined by high-throughput sequencing: genome structure and phylogenetic relationships. BMC Evol. Biol. 14, 151 (2014).
Yao, X., Tang, P. & Li, Z. The first complete chloroplast genome sequences in Actinidiaceae: genome structure and comparative analysis. Plos One 10, e129347 (2015).
Khan, A. et al. First complete chloroplast genomics and comparative phylogenetic analysis of Commiphora gileadensis and C. foliacea: Myrrh producing trees. Plos One 14, e208511 (2019).
Luo, C. et al. Comparative chloroplast genome analysis of Impatiens species (Balsaminaceae) in the Karst area of China: Insights Into genome evolution and phylogenomic implications. BMC Genom. 22, 1–18 (2021).
Wu, L. et al. Comparative and phylogenetic analyses of the chloroplast genomes of species of Paeoniaceae. Sci. Rep. 11, 14643 (2021).
Dobrogojski, J., Adamiec, M. & Ski, R. L. The chloroplast genome: A review. Acta Physiol. Plant. 42, 1–13 (2020).
Li, D., Zhao, C. & Liu, X. Complete chloroplast genome sequences of Kaempferia galanga and Kaempferia elegans: Molecular structures and comparative analysis. Molecules 24, 474 (2019).
Wu, L. et al. Comparative and phylogenetic analysis of the complete chloroplast genomes of three Paeonia section Moutan species (Paeoniaceae). Front. Genet. 11, 980 (2020).
Fan, R., Ma, W. & Liu, S. Integrated analysis of three newly sequenced fern chloroplast genomes: Genome structure and comparative analysis. Ecol. Evol. 11, 4550–4563 (2021).
Han, H., Qiu, R. & Liu, Y. Analysis of chloroplast genomes provides insights into the evolution of Agropyron. Front. Genet. 13, 832809 (2022).
Wang, N. A. et al. The complete chloroplast genomes of three Hamamelidaceae species: Comparative and phylogenetic analyses. Ecol. Evol. 12, 1–21 (2022).
Zhou, C. et al. Chloroplast genome structure and phylogenetic position of Lophatherum gracile. Mitochondrial DNA Part B, Resour. 6, 26–28 (2021).
Bankevich, A. A. et al. Spades: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 19, 455–477 (2012).
Katoh, K. & Standley, D. Mafft multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 30, 772–780 (2013).
Zhang, D. et al. Phylosuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 20, 348–355 (2020).
Luo, Z. et al. Reproductive isolation between sympatric sister species, Mussaenda kwangtungensis and M. pubescens var. Alba. J. Integr. Plant Biol. 57, 859–870 (2015).
Li, A., Wu, X., Zhang, D. & Barrett, S. C. H. Cryptic dioecy in Mussaenda pubescens (Rubiaceae): A species with stigma-height dimorphism. Ann. Bot. 106, 521–531 (2010).
Deng, X. & Zhang, D. Three new synonyms in Mussaenda (Ruhiaceae) from China. ACTA Phytotax. Sinica 44, 608–611 (2006).
Daniell, H., Lin, C., Yu, M. & Chang, W. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 17, 1–29 (2016).
Jansen, R. K. et al. Methods for obtaining and analyzing whole chloroplast genome sequences. Methods Enzymol. 395, 348–384 (2005).
Yen, L., Kousar, M. & Park, J. Comparative analysis of chloroplast genome of Desmodium stryacifolium with closely related legume genome from the phaseoloid clade. Int. J. Mol. Sci. 24, 6072 (2023).
Asaf, S., Khan, A. L. & Khan, A. R. Complete chloroplast genome of Nicotiana otophora and its comparison with related species. Front. Plant Sci. 7, 843 (2016).
Mehmood, F. et al. Chloroplast genome of Hibiscus Rosa-sinensis (Malvaceae): Comparative analyses and identification of mutational hotspots. Genomics 112, 581–591 (2020).
Hu, Y., Chen, X., Feng, X., Woeste, K. E. & Zhao, P. Characterization of the complete chloroplast genome of the endangered species Carya sinensis (Juglandaceae). Conserv. Genet. Resour. 8, 467–470 (2016).
Liu, L. et al. Chloroplast genome analyses and genomic resource development for epilithic sister genera Oresitrophe and Mukdenia (Saxifragaceae), using genome skimming data. BMC Genom. 19, 1–17 (2018).
Ulea, A. N. & Lobry, J. R. A new method for assessing the effect of replication on DNA base composition asymmetry. Mol. Biol. Evol. 24, 2169–2179 (2007).
Duan, R., Huang, M., Yang, L. & Liu, Z. Characterization of the complete chloroplast genome of Emmenopterys henryi (Gentianales: Rubiaceae), an endangered relict tree species endemic to China. Conserv. Genet. Resour. 9, 459–461 (2017).
Wang, H. et al. Complete plastome sequence of Mussaenda hirsutula Miq (Rubiaceae): An endemic shrub in South China. Mitochondrial DNA Part B: Resour. 4, 677–678 (2019).
Li, C. et al. Codon usage bias and genetic diversity in chloroplast genomes of Elaeagnus species (Myrtiflorae: Elaeagnaceae). Physiol. Mol. Biol. Plants 29, 239–251 (2023).
Gao, M. et al. Analysis of codon usage patterns in Bupleurum falcatum chloroplast genome. Chin. Herb. Med. 15, 284–290 (2023).
Duan, H. et al. Analysis of codon usage patterns of the chloroplast genome in Delphinium grandiflorum L. reveals a preference for at-ending codons as a result of major selection constraints. PeerJ. 9, e10787 (2021).
Huo, Z. et al. The complete chloroplast genome of Persicaria perfoliata and comparative analysis with four medicinal plants of Polygonaceae. Genome. 65, 377–389 (2022).
Yang, J., Yang, S., Li, H., Yang, J. & Li, D. Comparative chloroplast genomes of Camellia species. Plos One. 8, 1 (2013).
Kumar, S., Stecher, G. & Tamura, K. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 33, 1870–1874 (2016).
Funding
This research was funded by National Natural Science Foundation of China (32160727); Department of Education of Guizhou Province [Qianjiaoji(2023)027, QianjiaoheKYZi(2020)070, QianjiaoherencaiteamZi(2014)45]; and Qiannan Normal University for Nationalities (2024zdzk15).
Author information
Authors and Affiliations
Contributions
C.Z.: Methodology, Software, Writing-originaldraft. F.T.: Formal analysis, Investigation. X.W.: Data curation, Validation. R.L.: Software, Formalanalysis. X.Y.: Investigation, Conceptualization. L.X.: Supervision, Software. X.Z.: Su-pervision, Investigation, Writing-review & editing. T.G.: Validation, Visualization, Writing-review & editing.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhou, C., Tao, F., Long, R. et al. The complete chloroplast genome of Mussaenda pubescens and phylogenetic analysis. Sci Rep 14, 9131 (2024). https://doi.org/10.1038/s41598-024-55010-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-55010-y
Keywords
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
