
Phylogeny and Taxonomic Revision of the Genus Melanosciadium (Apiaceae), Based on Plastid Genomes and Morphological Evidence
 , and
, and
Abstract
:1. Introduction
2. Material and Methods
2.1. Taxon Sampling
2.2. Morphological Study
2.3. nrDNA Sequences and Plastid Genome Sequencing, Assembly, and Annotation
2.4. Plastid Genome Comparative Analyses
2.5. Phylogenetic Analysis
3. Result and Discussion
3.1. Morphological Result
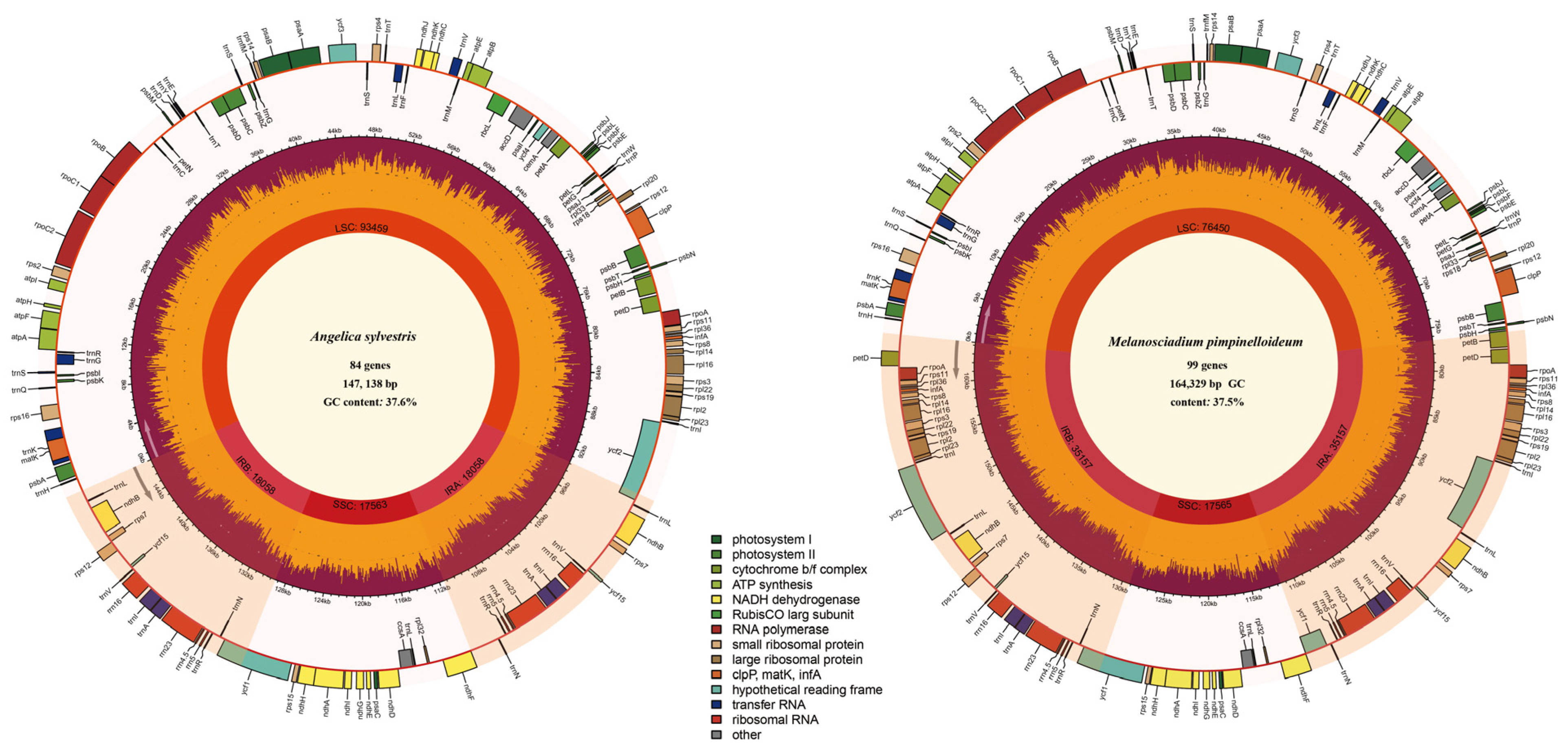
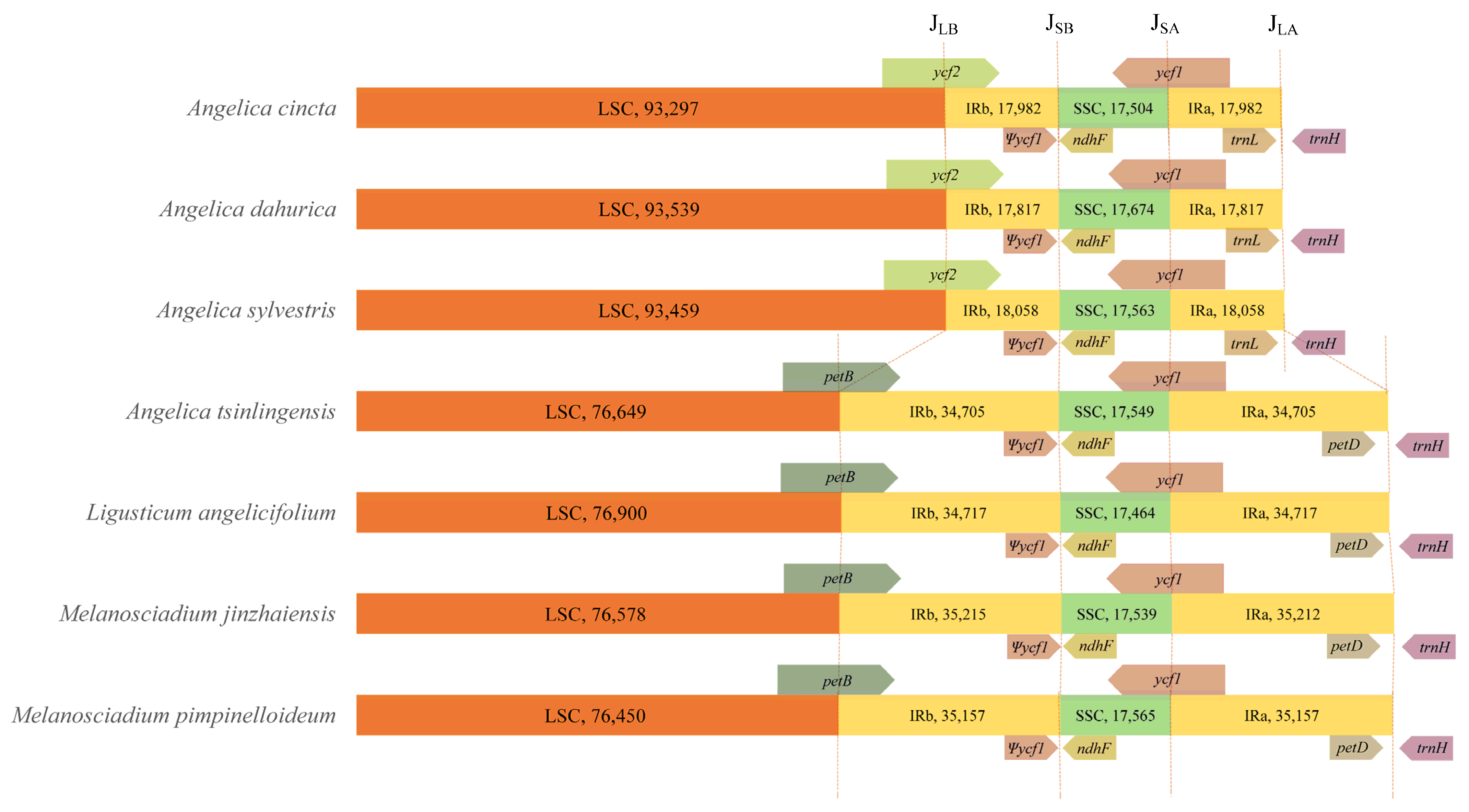
3.2. Plastid Genome Comparation Results
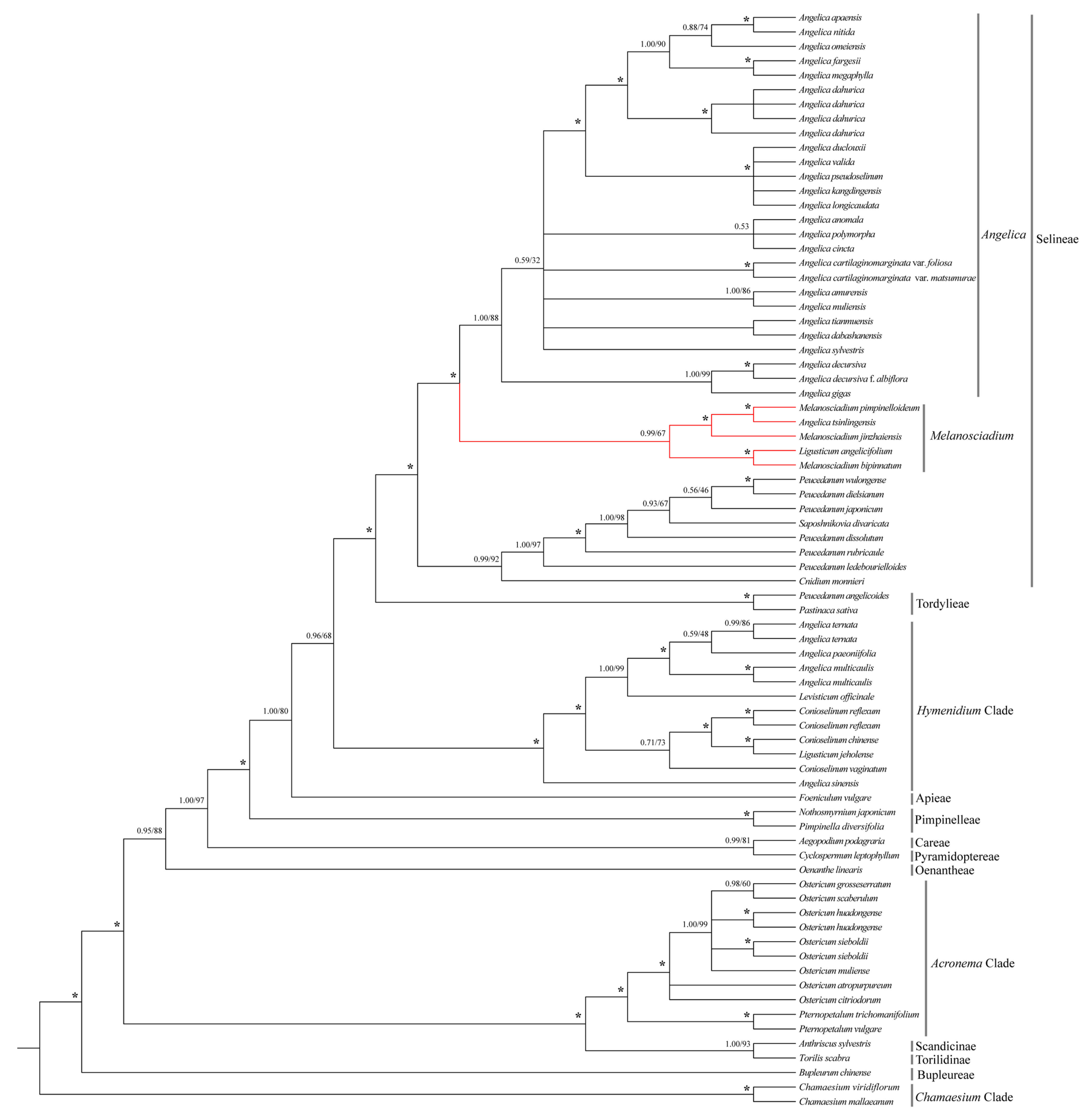
3.3. Phylogenetic Result
4. Taxonomic Treatment
- (1)
- (2)
- Melanosciadium tsinlingensis (K. T. Fu) Q. P. Jiang and X. J. He comb. nov.
- (3)
- Melanosciadium angelicifolium (Franch.) Q. P. Jiang and X. J. He comb. nov.
- 1a.
- Fruits are oblong-ovoid to subglobose, and all ribs are prominent, ridged.
- 2a.
- Petals purplish to purple; rays short, very unequal, rays about 5–14; anthers purple . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . M. pimpinelloideum
- 2b.
- Petals purplish; rays long, very unequal, rays about 14–30; anthers white. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . M. bipinnatum
- 1b.
- Fruits oblong-ovoid, oblong or suborbicular, dorsal ribs narrow-winged, lateral ribs winged.
- 3a.
- Rays fewer, less than 10.
- 4a.
- The rhachis of leaf blades refracted (bent back); rays unequal or subequal. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .M. genuflexum
- 4b.
- The rhachis of leaf blades not refracted; ray very unequal and short . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . M. jinzhaiensis
- 3b.
- Rays more, more than 10.
- 5a.
- Rays 10–20; petals white, the outer slightly enlarged . . . . . . M. tsinlingensis
- 5b.
- Rays 10–30; petals white to purple, the outer not enlarged . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .M. angelicifolium
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stevens, P.F. Onwards. “Apioideae”. Angiosperm Phylogeny Website. Version 14, February 2021. Available online: http://www.mobot.org/MOBOT/research/APweb/ (accessed on 3 January 2024).
- Calviño, C.I.; Teruel, F.E.; Downie, S.R. The role of the Southern Hemisphere in the evolutionary history of Apiaceae, a mostly north temperate plant family. J. Biogeogr. 2016, 43, 398–409. [Google Scholar] [CrossRef]
- Drude, C.G.O. Umbelliferae. In Die Natürlichen Pflanzenfamilien, Div.; Engler, A., Prantl, K., Eds.; Wilhelm Engelmann: Leipzig, Germany, 1898; Volume 3, p. 187. [Google Scholar]
- Pimenov, M.G.; Leonov, M.V. The Genera of the Umbelliferae. In A Nomenclator; Royal Botanical Gardens: Kew, UK, 1993. [Google Scholar]
- Downie, S.R.; Katz-Downie, D.S. A molecular phylogeny of Apiaceae subfamily Apioideae: Evience from nuclear ribosomal DNA internal transcribed spacer sequences. Am. J. Bot. 1996, 83, 234–251. [Google Scholar] [CrossRef]
- Downie, S.R.; Katz-Downie, D.S.; Cho, K.J. Phylogenetic analyses of Apiaceae subfamily Apioideae using nucleotide sequences from the chloroplast rpoC1 intron. Mol. Phylogenetics Evol. 1996, 6, 1–18. [Google Scholar] [CrossRef]
- Downie, S.R.; Ramanath, S.; Katz-Downie, D.S.; Llanas, E. Molecular systematics of Apiaceae subfamily Apioideae: Phylogenetic analyses of nuclear ribosomal DNA internal transcribed spacer and plastid rpoC1 intron sequences. Am. J. Bot. 1998, 85, 563–591. [Google Scholar] [CrossRef]
- Plunkett, G.M.; Downie, S.R. Major lineages within apiaceae subfamily apioideae: A comparison of chloroplast restriction site and dna sequence data. Am. J. Bot. 1999, 86, 1014–1026. [Google Scholar] [CrossRef]
- Downie, S.R.; Katz-Downie, D.S.; Watson, M.F. A phylogeny of the flowing plant family Apiaceae based on chloroplast DNA rpl16 and rpoC1 intron sequences: Towards a suprageneric classification of subfamily Apioideae. Am. J. Bot. 2000, 87, 273–292. [Google Scholar] [CrossRef]
- Downie, S.R.; Watson, M.F.; Spalik, K.; Katz-Downie, D.S. Molecular systematics of Old World Apioideae (Apiaceae): Relationships among some members of tribe Peucedaneae sensu lato, the placement of several island-endemic species, and resolution within the apioid superclade. Can. J. Bot. 2000, 78, 506–528. [Google Scholar]
- Downie, S.R.; Plunkett, G.M.; Watson, M.F.; Spalik, K.; Katz-Downie, D.S.; Valiejo-Roman, C.M.; Terentieva, E.I.; Troitsky, A.V.; Lee, B.-Y.; Lahham, J.; et al. Tribes and clades within Apiaceae subfamily Apioideae: The contribution of molecular data. Edinb. J. Bot. 2001, 58, 301–330. [Google Scholar] [CrossRef]
- Spalik, K.; Reduron, J.P.; Downie, S.R. The phylogenetic position of Peucedanum sensu lato and allied genera and their placement in tribe Selineae (Apiaceae, subfamily Apioideae). Plant Syst. Evol. 2004, 243, 189–210. [Google Scholar] [CrossRef]
- Downie, S.R.; Spalik, K.; Katz-Downie, D.S.; Reduron, J.-P. Major clades within Apiaceae subfamily Apioideae as inferred by phylogenetic analysis of nrDNA ITS sequences. Plant Divers. Evol. 2010, 128, 111–136. [Google Scholar] [CrossRef]
- Zhou, J.; Gong, X.; Downie, S.R.; Liu, Z.W.; Peng, H. Towards a more robust molecular phylogeny of Chinese Apiaceae subfamily Apioideae: Additional evidence from nrDNA ITS and cpDNA intron (rpl16 and rps16) sequences. Mol. Phylogenet. Evol. 2009, 53, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Peng, H.; Downie, S.R.; Liu, Z.W.; Gong, X. A molecular phylogeny of Chinese Apiaceae subfamily Apioideae inferred from nuclear ribosomal DNA internal transcribed spacer sequences. Taxon 2008, 57, 402–416. [Google Scholar]
- Iorizzo, M.; Ellison, S.; Senalik, D.; Zeng, P.; Satapoomin, P.; Huang, J.; Bowman, M.; Iovene, M.; Sanseverino, W.; Cavagnaro, P.; et al. A high-quality carrot genome assembly provides new insights into carotenoid accumulation and asterid genome evolution. Nat. Genet. 2016, 48, 657–666. [Google Scholar] [CrossRef]
- Wen, J.; Yu, Y.; Xie, D.F.; Peng, C.; Liu, Q.; Zhou, S.D.; He, X.J. A transcriptome-based study on the phylogeny and evolution of the taxonomically controversial subfamily Apioideae (Apiaceae). Ann. Bot. 2020, 125, 937–953. [Google Scholar] [CrossRef]
- Downie, S.R.; Jansen, R.K. A Comparative Analysis of Whole Plastid Genomes from the Apiales: Expansion and Contraction of the Inverted Repeat, Mitochondrial to Plastid Transfer of DNA, and Identification of Highly Divergent Noncoding Regions. Syst. Bot. 2015, 40, 336–351. [Google Scholar] [CrossRef]
- Gou, W.; Jia, S.B.; Price, M.; Guo, X.L.; Zhou, S.D.; He, X.J. Complete Plastid Genome Sequencing of Eight Species from Hansenia, Haplosphaera and Sinodielsia (Apiaceae): Comparative Analyses and Phylogenetic Implications. Plants 2020, 9, 1523. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Xie, D.; Peng, C.; Gui, L.; Price, M.; Zhou, S.; He, X. Molecular evolution and phylogenetic relationships of Ligusticum (Apiaceae) inferred from the whole plastome sequences. BMC Ecol. Evol. 2022, 22, 55. [Google Scholar] [CrossRef]
- Liu, C.K.; Lei, J.Q.; Jiang, Q.P.; Zhou, S.D.; He, X.J. The complete plastomes of seven Peucedanum plants: Comparative and phylogenetic analyses for the Peucedanum genus. BMC Plant Biol. 2022, 22, 101. [Google Scholar] [CrossRef] [PubMed]
- Samigullin, T.; Logacheva, M.; Terentieva, E.; Degtjareva, G.; Pimenov, M.; Valiejo-Roman, C. Plastid Phylogenomic Analysis of Tordylieae Tribe (Apiaceae, Apioideae). Plants 2022, 11, 709. [Google Scholar] [CrossRef]
- Cai, J.; Qin, H.-H.; Lei, J.-Q.; Liu, C.-K.; He, X.-J.; Zhou, S.-D. The phylogeny of Seseli (Apiaceae, Apioideae): Insights from molecular and morphological data. BMC Plant Biol. 2022, 22, s12870–s12822. [Google Scholar] [CrossRef]
- Clarkson, J.J.; Zuntini, A.R.; Maurin, O.; Downie, S.R.; Plunkett, G.M.; Nicolas, A.N.; Smith, J.F.; Feist, M.A.E.; Gutierrez, K.; Malakasi, P.; et al. A higher-level nuclear phylogenomic study of the carrot family (Apiaceae). Am. J. Bot. 2021, 108, 1252–1269. [Google Scholar] [CrossRef]
- Shan, R.H.; Sheh, M.L. Umbelliferae. In Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 1992; Volume 55, p. 560. [Google Scholar]
- Pan, Z.H.; Waston, M.F. Apiaceae (Umbelliferae). In Flora of China; Wu, Z.Y., Raven, R.H., Eds.; Science Press and Missouri Botanical Garden Press: Beijing, China; St. Louis, MI, USA, 2005; Volume 14, p. 18. [Google Scholar]
- She, M.L.; Su, P. The floristic analysis of endemic genera in Chinese Umbelliferae. Bull. Nanjing Bot. Gard. Mem. Sun Yat Sen 1987, 14–26. [Google Scholar]
- Fang, Z.X.; Liao, C.L. Annals of Medicinal Plants in Enshi, Hubei; Hubei Science and Technology Press: Wuhan, China, 2006. [Google Scholar]
- Pimenov, M.G.; Kljuykov, E.V. Generic disposition of Vicatia bipinnata in Melanosciadium and a new species (Umbelliferae). Feddes Repert. 2006, 117, 466–475. [Google Scholar] [CrossRef]
- Pimenov, M.G. Updated checklist of Chinese Umbelliferae: Nomenclature, synonymy, typification, distribution. Turczaninowia 2017, 20, 106–239. [Google Scholar] [CrossRef]
- Tan, J.B.; Wang, Z.X.; Xie, D.F.; He, X.J. Pimpinella rhomboidea var. tenuiloba is a synonym of Melanosciadium bipinnatum (Apiaceae). Nord. J. Bot. 2015, 33, 659–661. [Google Scholar] [CrossRef]
- Liao, C.; Downie, S.R.; Li, Q.; Yu, Y.; He, X.; Zhou, B. New Insights into the Phylogeny of Angelica and its Allies (Apiaceae) with Emphasis on East Asian Species, Inferred from nrDNA, cpDNA, and Morphological Evidence. Syst. Bot. 2013, 38, 266–281. [Google Scholar] [CrossRef]
- Liao, C.Y.; Gao, Q.; Katz-Downie, D.S.; Downie, S.R. A systematic study of North American Angelica species (Apiaceae) based on nrDNA ITS and cpDNA sequences and fruit morphology. J. Syst. Evol. 2022, 60, 789–808. [Google Scholar] [CrossRef]
- Leute, G.-H. Untersuchungen über den Yerwandtschaftskreis der Gattung Ligusticum L. (Umbelliferae). Ann. Naturhist. Mus. Wien 1969, 73, 55–98. [Google Scholar]
- Pimenov, M.G.; Kljuykov, E.V. New nomenclatural combinations for Chinese Umbelliferae. Feddes Repert. 1999, 110, 481–491. [Google Scholar] [CrossRef]
- Govaerts, R. The World Checklist of Vascular Plants (WCVP). R. Bot. Gard. Kew. 2023. Available online: https://www.gbif.org/ (accessed on 3 January 2024).
- Guisinger, M.M.; Kuehl, J.V.; Boore, J.L.; Jansen, R.K. Extreme reconfiguration of plastid genomes in the angiosperm family Geraniaceae: Rearrangements, repeats, and codon usage. Mol. Biol. Evol. 2011, 28, 583–600. [Google Scholar] [CrossRef]
- Xie, D.F.; Yu, H.X.; Price, M.; Xie, C.; Deng, Y.Q.; Chen, J.P.; Yu, Y.; Zhou, S.D.; He, X.J. Phylogeny of Chinese Allium Species in Section Daghestanica and Adaptive Evolution of Allium (Amaryllidaceae, Allioideae) Species Revealed by the Chloroplast Complete Genome. Front. Plant Sci. 2019, 10, 460. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.L.; Zheng, H.Y.; Price, M.; Zhou, S.D.; He, X.J. Phylogeny and Comparative Analysis of Chinese Chamaesium Species Revealed by the Complete Plastid Genome. Plants 2020, 9, 965. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Ma, W.; Yang, X.; Wang, L.; Zhao, T.; Liang, L.; Wang, G.; Ma, Q. Plastome phylogenomics provide new perspective into the phylogeny and evolution of Betulaceae (Fagales). BMC Plant Biol. 2022, 22, 611. [Google Scholar] [CrossRef] [PubMed]
- Ravi, V.; Khurana, J.P.; Tyagi, A.K.; Khurana, P. An update on chloroplast genomes. Plant Syst. Evol. 2008, 271, 101–122. [Google Scholar] [CrossRef]
- Ren, T.; Li, Z.-X.; Xie, D.-F.; Gui, L.-J.; Peng, C.; Wen, J.; He, X.-J. Plastomes of eight Ligusticum species: Characterization, genome evolution, and phylogenetic relationships. BMC Plant Biol. 2020, 20, 519. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cai, J.; Qin, H.H.; Price, M.; Zhang, Z.; Yu, Y.; Xie, D.F.; He, X.J.; Zhou, S.D.; Gao, X.F. Phylogeny, Age, and Evolution of Tribe Lilieae (Liliaceae) Based on Whole Plastid Genomes. Front. Plant Sci. 2021, 12, 699226. [Google Scholar] [CrossRef]
- Yang, Z.; Ferguson, D.K.; Yang, Y. New insights into the plastome evolution of Lauraceae using herbariomics. BMC Plant Biol. 2023, 23, 387. [Google Scholar] [CrossRef]
- Xue, T.-T.; Janssens, S.B.; Liu, B.-B.; Yu, S.-X. Phylogenomic conflict analyses of the plastid and mitochondrial genomes via deep genome skimming highlight their independent evolutionary histories: A case study in the cinquefoil genus Potentilla sensu lato (Potentilleae, Rosaceae). Mol. Phylogenetics Evol. 2024, 190, 107956. [Google Scholar] [CrossRef]
- Fu, N.; Ji, M.; Rouard, M.; Yan, H.-F.; Ge, X.-J. Comparative plastome analysis of Musaceae and new insights into phylogenetic relationships. BMC Genom. 2022, 23, 223. [Google Scholar] [CrossRef]
- Alzahrani, D.; Albokhari, E.; Yaradua, S.; Abba, A. Complete chloroplast genome sequences of Dipterygium glaucum and Cleome chrysantha and other Cleomaceae Species, comparative analysis and phylogenetic relationships. Saudi J. Biol. Sci. 2021, 28, 2476–2490. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, S.-Q.; Xie, H.-H.; Liu, J.-Q.; Gao, X.-F. Comparative plastid genome analyses of Rosa: Insights into the phylogeny and gene divergence. Tree Genet. Genomes 2022, 18, 20. [Google Scholar] [CrossRef]
- Wang, M.; Wang, X.; Sun, J.; Wang, Y.; Ge, Y.; Dong, W.; Yuan, Q.; Huang, L. Phylogenomic and evolutionary dynamics of inverted repeats across Angelica plastomes. BMC Plant Biol. 2021, 21, 26. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Li, J.; Zuo, Y.; Qin, Q.; Zeng, S.; Rennenberg, H.; Deng, H. Plastome variations reveal the distinct evolutionary scenarios of plastomes in the subfamily Cereoideae (Cactaceae). BMC Plant Biol. 2023, 23, 132. [Google Scholar] [CrossRef] [PubMed]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.J.; Moore, M.J.; Li, D.Z.; Yi, T.S. PGA: A software package for rapid, accurate, and flexible batch annotation of plastomes. Plant Methods 2019, 15, 50. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y.; dePamphilis, C.W.; Yi, T.S.; Li, D.Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Zheng, S.; Poczai, P.; Hyvonen, J.; Tang, J.; Amiryousefi, A. Chloroplot: An Online Program for the Versatile Plotting of Organelle Genomes. Front. Genet. 2020, 11, 576124. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest, a Program to Evaluate the Fit of Several Models of Evolution to a Given Data and Unrooted Tree, version 2.2.; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Wen, J.; Xie, D.F.; Price, M.; Ren, T.; Deng, Y.Q.; Gui, L.J.; Guo, X.L.; He, X.J. Backbone phylogeny and evolution of Apioideae (Apiaceae): New insights from phylogenomic analyses of plastome data. Mol. Phylogenet. Evol. 2021, 161, 107183. [Google Scholar] [CrossRef]
- Pimenov, M.G.; Kljuykov, E.V.; Ostroumova, T.A. Reduction of Notopterygium to Hansenia (Umbelliferae). Willdenowia 2008, 38, 155–172. [Google Scholar] [CrossRef]
- Jia, S.-B.; Guo, X.-L.; Zhou, S.-D.; He, X.-J. Hansenia pinnatiinvolucellata is conspecific with H. weberbaueriana (Apiaceae) based on morphology and molecular data. Phytotaxa 2019, 418, 203–210. [Google Scholar] [CrossRef]
- Tan, J.-B.; Jia, S.-B.; He, X.-J.; Ma, X.-G. Accommodating Haplosphaera in Hansenia (Apiaceae) based on morphological and molecular evidence. Phytotaxa 2020, 464, 207–216. [Google Scholar] [CrossRef]
- Jiang, Q.-P.; Liu, C.-K.; Xie, D.-F.; Zhou, S.-D.; He, X.-J. Plastomes Provide Insights into Differences between Morphology and Molecular Phylogeny: Ostericum and Angelica (Apiaceae) as an Example. Diversity 2022, 14, 776. [Google Scholar] [CrossRef]
- Spooner, D.M.; Ruess, H.; Iorizzo, M.; Senalik, D.; Simon, P. Entire plastid phylogeny of the carrot genus (Daucus, Apiaceae): Concordance with nuclear data and mitochondrial and nuclear DNA insertions to the plastid. Am. J. Bot. 2017, 104, 296–312. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.Y.; Guo, X.L.; Price, M.; He, X.J.; Zhou, S.D. Effects of Mountain Uplift and Climatic Oscillations on Phylogeography and Species Divergence of Chamaesium (Apiaceae). Front. Plant Sci. 2021, 12, 673200. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.; Deusch, O.; Stawski, N.; Grunheit, N.; Goremykin, V. Chloroplast genome phylogenetics: Why we need independent approaches to plant molecular evolution. Trends Plant Sci. 2005, 10, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Raubeson, L.A.; Peery, R.; Chumley, T.W.; Dziubek, C.; Fourcade, H.M.; Boore, J.L.; Jansen, R.K. Comparative chloroplast genomics: Analyses including new sequences from the angiosperms Nuphar advena and Ranunculus macranthus. BMC Genom. 2007, 8, 174. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Gao, L.; Wang, B.; Su, Y.J.; Wang, T. The complete chloroplast genome sequence of Cephalotaxus oliveri (Cephalotaxaceae): Evolutionary comparison of Cephalotaxus chloroplast DNAs and insights into the loss of inverted repeat copies in gymnosperms. Genome Biol. Evol. 2013, 5, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Downie, S.R.; Yu, Y.; Zhang, X.; Chen, W.; He, X.; Liu, S. Molecular systematics of Angelica and allied genera (Apiaceae) from the Hengduan Mountains of China based on nrDNA ITS sequences: Phylogenetic affinities and biogeographic implications. J. Plant Res. 2009, 122, 403–414. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Character | Taxon | ||||
|---|---|---|---|---|---|
| M. pimpinelloideum | M. bipinnatum | A. tsinlingensis | L. angelicifolium | M. jinzhaiensis | |
| Live form | monocarpic | monocarpic | monocarpic | monocarpic | monocarpic |
| Plant height (cm) | 50–150 | 60–150 | 60–150 | 80–180 | 40–140 |
| Leaf in outline (basal) | broadly ovate-triangular, 1–2-ternate | broadly ovate-triangular, 2–3-ternate | broadly ovate-triangular, 1–2-ternate | broadly ovate-triangular, 2-ternate | broadly ovate-triangular, 1–2-ternate |
| median leaflets (pinnae) (basal) | rhombic to narrow-ovate, 7–15 × 3–9 cm, base cuneate, margin incised-serrate, teeth mucronate, with attenuate apex | rhombic-obovate, 3–7.5 × 2–4 cm, base cuneate, margin incised-serrate, teeth mucronate, with attenuate apex | rhombic-obovate, 7–13 × 5–9 cm, base cuneate, margin incised-serrate, teeth mucronate, with attenuate apex | rhombic to oblong-ovate, 3.5–6 × 1.5–3 cm, sometimes 3-lobed, base cuneate, margin incised-serrate, teeth mucronate, with attenuate apex | short-petiolulate, rhombic-obovate, 10–15 × 5–10 cm, base cuneate; margin incised-serrate, teeth mucronate, with attenuate apex |
| lateral leaflets (pinnae) (basal) | ovate or longovate, 3–10 × 2–6 cm, base truncate, margin incised-serrate, teeth mucronate, apex acuminate | rhombic-obovate to longovate, 2–5.5 × 1–3 cm, base cuneate, margin incised-serrate, teeth mucronate, with attenuate apex | broad-ovate, 5–12 × 3–7 cm, base truncate, margin incised-serrate, teeth mucronate, apex acuminate | rhombic to oblong-ovate, 1–5 × 1–2 cm, sometimes 1–2-lobed, base cuneate, margin incised-serrate, teeth mucronate, with attenuate apex | broad-ovate, 7–12 × 4–7 cm, sometimes 1–2-lobed, base truncate, margin incised-serrate, teeth mucronate, apex acuminate or cuspidatus |
| Sheaths | not inflated to inflated | slightly inflated | not inflated | inflated | not inflated |
| Umbels rays | 6–13, short, very unequal | 14–30, very unequal | 9–20, very unequal | 10–30, very unequal | 3–12, very unequal, short |
| Petals | purplish to purple; the outer not enlarged | purplish, the outer not enlarged | white, the outer slightly enlarged | white to purple, the outer not enlarged | purplish; the outer not enlarged |
| Calyx teeth | absent or very small | absent | absent | absent | minute |
| Fruit | ovoid-globose to subglobose; 3.01–4.45 × 4.16–5.64 mm | oblong-ovoid to subglobose; 2.34–4.64 × 3.28–3.93 | oblong to suborbicular; 2.96–3.88 × 4.19–5.54 | oblong-ovoid; 2.95–3.96 × 5.08–6.46 | oblong; 2.43–3.84 × 4.03–6.68 |
| Mericarp ribs | 5 ribs; ribs prominent, ridged | 5 ribs; ribs prominent, ridged | 5 ribs; dorsal ribs narrow-winged, lateral ribs winged to broad-winged | 5 ribs; dorsal ribs narrow-winged, lateral ribs winged | 3–5 ribs; dorsal ribs narrow-winged to winged, lateral ribs winged to broad-winged |
| Endosperm (at commissural side) | plane to slightly concave | concave | plane to concave | slightly concave to concave | plane to slightly concave |
| Vittae in dorsal furrows | 3–4 | 2–5 | 2–4 | 1–4 | 2–4 |
| Vittae in commissure | 4–6 | 6–8 | 4–8 | 4–6 | 6–8 |
| Species | Size (bp) | LSC Length (bp) | SSC Length (bp) | IR Length (bp) | Number of Different Genes/Total Number of Genes | Number of Different Protein-Coding Genes (Duplicated in IR) | Number of Different tRNA Genes (Duplicated in IR) | Number of Different rRNA Genes (Duplicated in IR) | Number of Genes Duplicated in IR | GC Content (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| Angelica dahurica | 146,847 | 93,539 | 17,674 | 17,817 | 114/129 | 80 (4) | 30 (6) | 4 (4) | 15 | 37.5 |
| Angelica cincta | 146,765 | 93,297 | 17,504 | 17,982 | 114/129 | 80 (4) | 30 (6) | 4 (4) | 15 | 37.5 |
| Angelica sylvestris | 147,138 | 93,459 | 17,563 | 18,058 | 114/129 | 80 (4) | 30 (6) | 4 (4) | 15 | 37.6 |
| Angelica tsinlingensis | 163,608 | 76,649 | 17,549 | 34,705 | 114/144 | 80 (18) | 30 (7) | 4 (4) | 30 | 37.5 |
| Ligusticum angelicifolium | 163,798 | 76,900 | 17,464 | 34,717 | 114/144 | 80 (18) | 30 (7) | 4 (4) | 30 | 37.4 |
| Melanosciadium jinzhaiensis | 164,544 | 76,578 | 17,539 | 35,212 (35,215) | 114/144 | 80 (18) | 30 (7) | 4 (4) | 30 | 37.4 |
| Melanosciadium pimpinelloideum | 164,329 | 76,450 | 17,565 | 35,157 | 114/144 | 80 (18) | 30 (7) | 4 (4) | 30 | 37.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Q.-P.; Guo, X.-L.; Zhao, A.-Q.; Fan, X.; Li, Q.; Zhou, S.-D.; He, X.-J. Phylogeny and Taxonomic Revision of the Genus Melanosciadium (Apiaceae), Based on Plastid Genomes and Morphological Evidence. Plants 2024, 13, 907. https://doi.org/10.3390/plants13060907
Jiang Q-P, Guo X-L, Zhao A-Q, Fan X, Li Q, Zhou S-D, He X-J. Phylogeny and Taxonomic Revision of the Genus Melanosciadium (Apiaceae), Based on Plastid Genomes and Morphological Evidence. Plants. 2024; 13(6):907. https://doi.org/10.3390/plants13060907
Chicago/Turabian StyleJiang, Qiu-Ping, Xian-Lin Guo, An-Qi Zhao, Xing Fan, Qing Li, Song-Dong Zhou, and Xing-Jin He. 2024. "Phylogeny and Taxonomic Revision of the Genus Melanosciadium (Apiaceae), Based on Plastid Genomes and Morphological Evidence" Plants 13, no. 6: 907. https://doi.org/10.3390/plants13060907