

The Position of Lophozia dubia (Lophoziaceae, Marchantiophyta) in the Phylogenetic System of Lophozia and the Distribution of Lophozia in Southeast Eurasia, Extending to Indonesia

Abstract
:1. Introduction
2. Results
2.1. Molecular Genetics
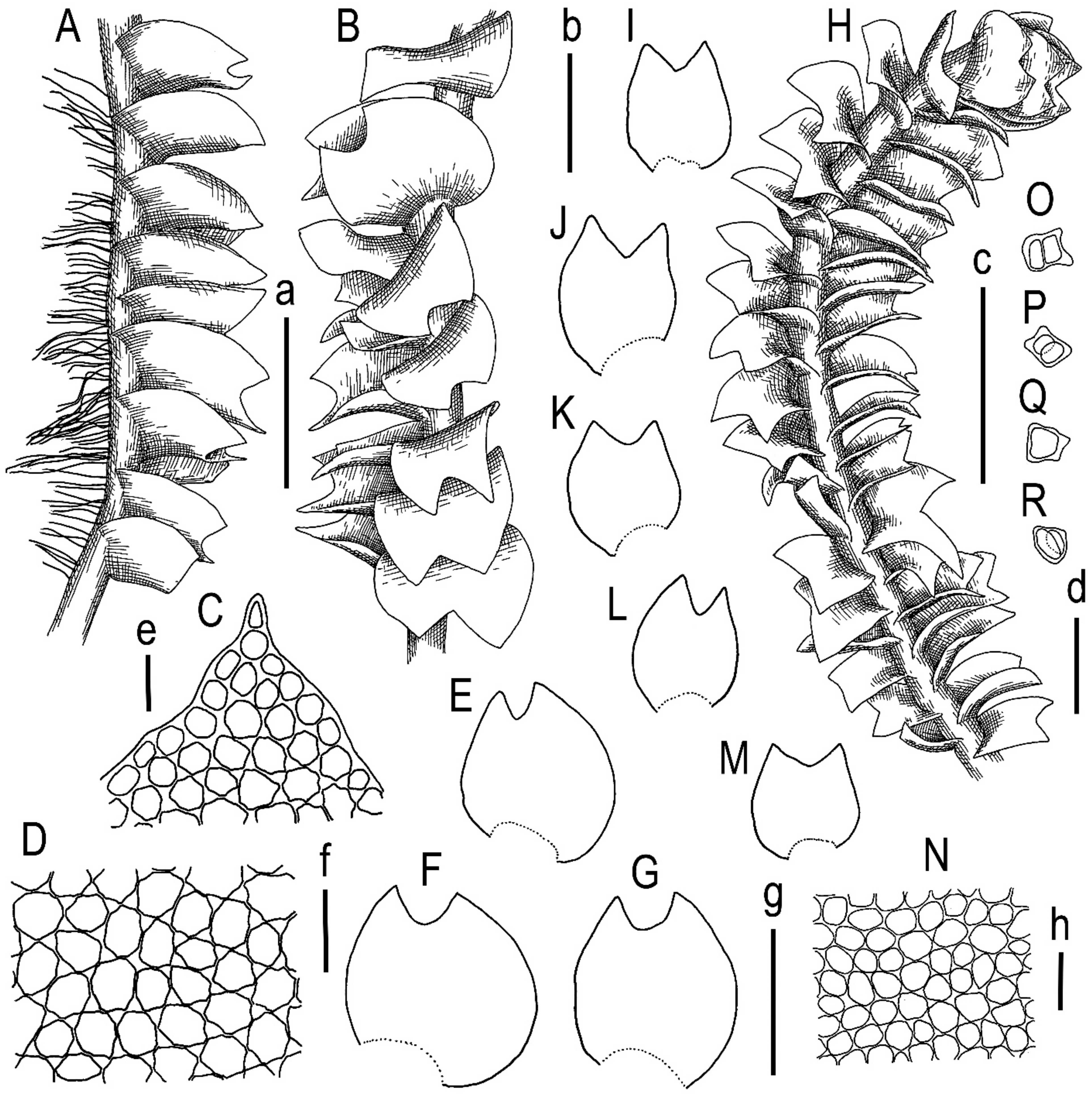
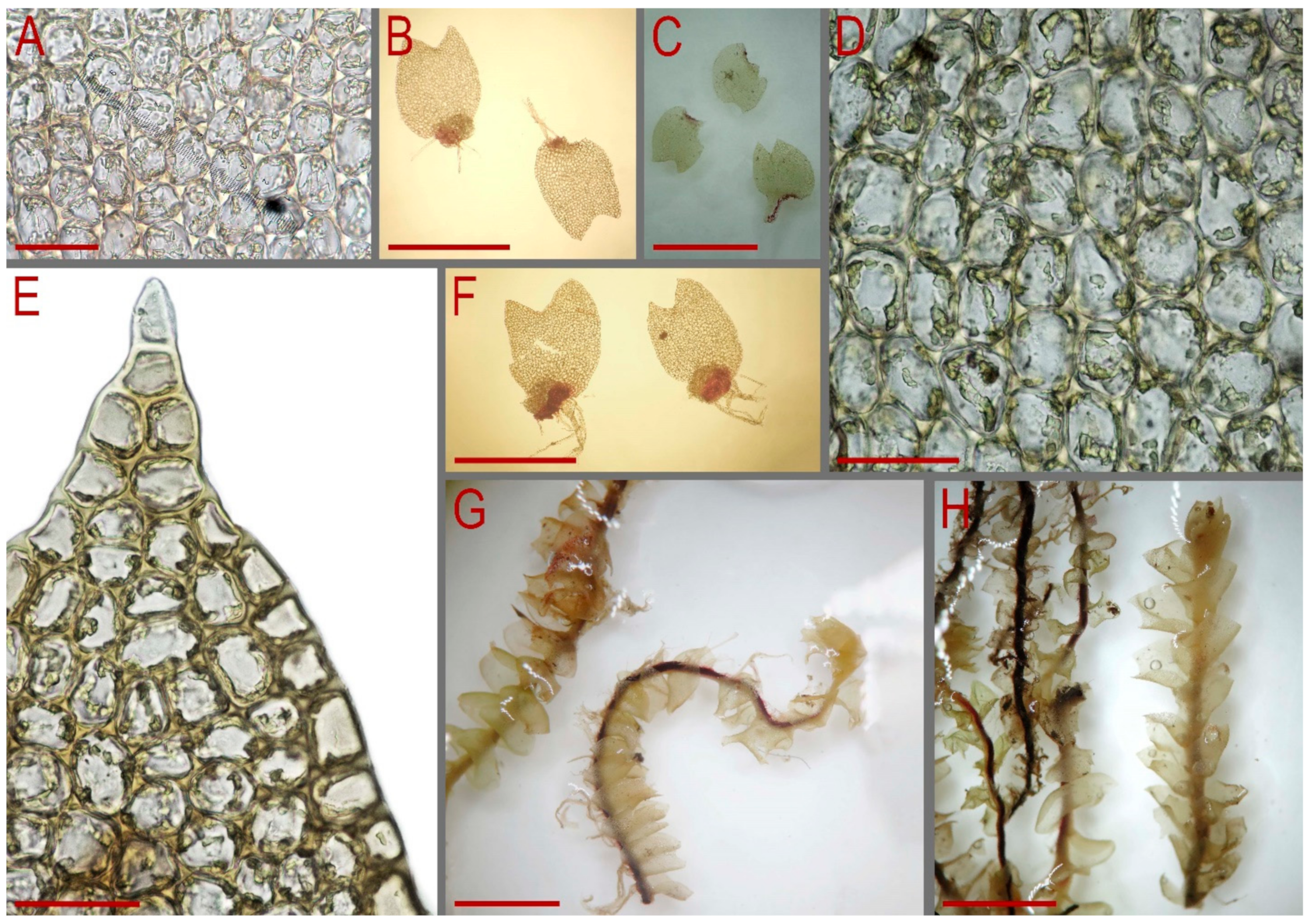
2.2. Morphology
3. Discussion
- =
- Anastrophyllum pallidum Steph., Bull. Herb. Boissier ser. 2, 1: 1131 (Sp. Hepat. 2: 114), 1901 (Lophozia pallida (Steph.) Grolle, J. Jap. Bot. 39: 174, 1964) syn. nov.
- =
- Lophozia handelii Herzog, Symb. Sin. 5: 14, 1930 syn. nov.
4. Materials and Methods
4.1. Morphological Studies
4.2. Molecular Genetic Research
4.2.1. Taxon Sampling
4.2.2. DNA Isolation, Amplification and Sequencing
4.2.3. Phylogenetic Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Du Mortier, B.-C. Sylloge Jungermannidearum Europæ Indigenarum: Earum Genera et Species Systematicé Complectens; Biodiversity Heritage Library: Washington, DC, USA, 1831; 110p. [Google Scholar] [CrossRef]
- Du Mortier, B.-C. Recueil d’Observations sur les Jungermanniacées; Biodiversity Heritage Library: Washington, DC, USA, 1835; 27p. [Google Scholar] [CrossRef]
- Buch, H. Vorarbeiten zu einer Lebermoosflora Fenno-scandias. I. Ein Versuch zur Aufspaltung der Gattungen Lophozia Dum. und Sphenolobus Steph. Memo. Soc. Pro Fauna Flora Fenn. 1932, 8, 282–297. [Google Scholar]
- Buch, H. Vorarbeiten zur eine Lebermoosflora Fenno-Scandias. VII. Über die verwandtschaftlichen Beziehungen zwischen den Arten der Gattungen Sphenolobus St. sens. lat. und Lophozia Dum. sens. lat. Memo. Soc. Pro Fauna Flora Fenn. 1942, 17, 283–290. [Google Scholar]
- Buch, H. Lophozia perssonii Buch et S. Arnell, spec. nov. Bot. Not. 1944, 97, 381–387. [Google Scholar]
- Schuster, R.M. Notes on nearctic hepaticae. V. The status of Lophozia gracillima Buch and its relation to Lophozia longidens, Lophozia porphyroleuca and Sphenolobus ascendens. Bryologist 1952, 55, 173–185. [Google Scholar] [CrossRef]
- Schuster, R.M. Boreal hepaticae. A manual of the liverworts of Minnesota and adjacent regions. Am. Midl. Nat. 1953, 49, 257–684. [Google Scholar] [CrossRef]
- Evans, A.W. Recent segregates from the genera Lophozia and Sphenolobus. Bryologist 1935, 38, 61–70. [Google Scholar] [CrossRef]
- Andrews, A.L.R. Taxonomic Notes, VII. The Lophozia–Sphenolobus Complex. Bryologist 1948, 49, 33–37. [Google Scholar] [CrossRef]
- Söderström, L.; Séneca, A.N.A. World distribution patterns in the Lophoziaceae/Scapaniaceae complex (Hepaticae, Bryophyta). J. Hattori Bot. Lab. 2006, 100, 431–441. [Google Scholar]
- Schuster, R.M. The hepaticae of Prince Edward Islands. II. On Gymnocoleopsis (Schust.) Schust., Lophozia cylindriformis (Mitt.) Steph. and the subgeneric classification of the genus Lophozia Dumort. J. Hattori Bot. Lab. 1995, 78, 119–135. [Google Scholar]
- Konstantinova, N.A.; Vilnet, A.A. New taxa and new combinations in Jungermanniales (Hepaticae). Arctoa 2009, 18, 65–67. [Google Scholar] [CrossRef]
- Söderström, L.; Hagborg, A.; von Konrat, M. Early Land Plants Today. Index of liverwort and hornwort names published 2017–2018. Phytotaxa 2020, 440, 1–24. [Google Scholar] [CrossRef]
- Söderström, L.; Váňa, J.; Hagborg, A.; von Konrat, M. Notes on early land plants today. 35. Notes on Lophoziaceae (Marchantiophyta). Phytotaxa 2013, 97, 27–35. [Google Scholar] [CrossRef]
- Vilnet, A.A.; Konstantinova, N.A.; Troitsky, A.V. Molecular insight on phylogeny and systematics of the Lophoziaceae, Scapaniaceae, Gymnomitriaceae and Jungermanniaceae. Arctoa 2010, 19, 31–50. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Vilnet, A.A. Lophozia fuscovirens sp. nov. (Lophoziaceae, Marchantiophyta): The second taxon with brown gemmae within Lophozia s.s. Nord. J. Bot. 2019, 37, e02294. [Google Scholar] [CrossRef]
- Ellis, L.T.; Afonina, O.M.; Czernyadjeva, I.V.; Ivchenko, T.G.; Kholod, S.S.; Kotkova, V.M.; Kuzmina, E.Y.; Potemkin, A.D.; Sergeeva, Y.M.; Asthana, A.K.; et al. New national and regional bryophyte records, 61. J. Bryol. 2019, 41, 364–384. [Google Scholar] [CrossRef]
- Konstantinova, N.A.; Vilnet, A.A.; Mamontov, Y.S. A new species of the genus Lophozia (Lophoziaceae) from the Svalbard Archipelago. Arctoa 2020, 29, 124–134. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Fedosov, V.E.; Maltseva, Y.D.; Milyutina, I.A.; Klimova, K.G.; Nguyen, H.M.; Troitsky, A.V. Overview of Schistochilopsis (Hepaticae) in Pacific Asia with the Description Protochilopsis gen. nov. Plants 2020, 9, 850. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Maltseva, Y.D.; Vilnet, A.A.; Choi, S.S. The transfer of Tritomaria koreana to Lophozia has led to recircumscription of the genus and shown convergence in Lophoziaceae (Hepaticae). Phytotaxa 2021, 512, 41–56. [Google Scholar] [CrossRef]
- Bakalin, V.A. Notes on Lophozia IV. Some new taxa of Lophozia sensu stricto. Ann. Bot. Fenn. 2003, 40, 47–52. [Google Scholar]
- Hodgetts, N.G.; Söderström, L.; Blockeel, T.L.; Caspari, S.; Ignatov, M.S.; Konstantinova, N.A.; Lockhart, N.; Papp, B.; Schröck, C.; Sim-Sim, M.; et al. An annotated checklist of bryophytes of Europe, Macaronesia and Cyprus. J. Bryol. 2020, 42, 1–116. [Google Scholar] [CrossRef]
- Choi, S.S.; Bakalin, V.A.; Kim, C.H.; Sun, B.Y. Unrecorded Liverwort species from Korean flora. Korean J. Plant Taxon. 2012, 42, 80–90. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Xiong, Y. Lophozia silvicola (Scapaniaceae, Hepaticae)—An Unexpected Record of a Boreal Species in the Subtropical Forest of Guizhou (Southern China). Herzogia 2015, 28, 44–49. [Google Scholar] [CrossRef]
- Bakalin, V.A. Monograficheskaia Obrabotka Roda Lophozia (Dumort.) Dumort. s. str. [Monograph of the Genus Lophozia (Dumort.) Dumort. s. str.]; Nauka: Moscow, Russia, 2005; 240p. [Google Scholar]
- Grolle, R. Die Lebermoose Nepals. In Khumbu Himal—Ergebnisse des Forschungsunternehmens Nepal Himalaya; Hellmich, W., Ed.; Springer: Berlin/Heidelberg, Germany, 1966; pp. 262–298. [Google Scholar]
- Long, D.G.; Grolle, R. Hepaticae of Bhutan II. J. Hattori Bot. Lab. 1990, 68, 381–440. [Google Scholar] [CrossRef]
- Kitagawa, N. A revision of the family Lophoziaceae of Japan and adjacent regions. I. J. Hattori Bot. Lab. 1965, 28, 239–291. [Google Scholar]
- Schiffner, V. Expositio plantarum in itinere suo Indico annis 1893–1894 suscepto collectarum I. Denkschr./Akad. Wiss. Wien Math.-Naturwissenschaftliche Kl. 1898, 67, 153–203. [Google Scholar]
- Söderström, L.; Gradstein, S.R.; Hagborg, A. Checklist of the hornworts and liverworts of Java. Phytotaxa 2010, 9, 53–149. [Google Scholar] [CrossRef]
- Váňa, J.; Söderström, L.; Hagborg, A.; von Konrat, M. Notes on early land plants today. 8. New combinations and some lectotypifications in Mesoptychia. Phytotaxa 2012, 65, 52–56. [Google Scholar] [CrossRef]
- Herzog, T. Lebermoose aus Sumatra. Ann. Naturhistorischen Mus. Wien 1943, 53, 348–373. [Google Scholar]
- Bakalin, V.A. The status and treatment of the genus Hattoriella (H. Inoue) H. Inoue. Arctoa 2003, 12, 91–96. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Choi, S.S.; Sun, B.-Y. A new species of Tritomaria (Lophoziaceae) from the Korean Peninsula. Arctoa 2010, 18, 163–166. [Google Scholar] [CrossRef]
- Index Herbariorum. Available online: https://sweetgum.nybg.org/science/ih/ (accessed on 4 January 2024).
- Bakalin, V.A.; Fedosov, V.E.; Klimova, K.G.; Maltseva, Y.D.; Fedorova, A.V.; Choi, S.S. Konstantinovia Is Not Monotypic, and a New Attempt to Determine Relationships in Cephaloziellaceae–Scapaniaceae Superclade (Marchantiophyta). Plants 2024, 13, 15. [Google Scholar] [CrossRef]
- Friedl, T. Evolution of the polyphyletic genus Pleurastrum (Chlorophyta): Inferences from nuclear-encoded ribosomal DNA sequences and motile cell ultrastructure. Phycologia 1996, 35, 456–469. [Google Scholar] [CrossRef]
- Milyutina, I.A.; Goryunov, D.V.; Ignatov, M.S.; Ignatova, E.A.; Troitsky, A.V. The phylogeny of Schistidium (Bryophyta, Grimmiaceae) based on the primary and secondary structure of nuclear rDNA internal transcribed spacers. Mol. Biol. 2010, 44, 883–897. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 15, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Felsenstein, J. Maximum Likelihood and Minimum-Steps Methods for Estimating Evolutionary Trees from Data on Discrete Characters. Syst. Biol. 1973, 22, 240–249. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F.; Nielsen, R.; Bollback, J.P. Bayesian inference of phylogeny and its impact on evolutionary biology. Science 2001, 294, 2310–2314. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| № | Taxon | Infraspecific p-Distances, ITS1–2/trnL–F, | Interspecific p-Distances, ITS1–2/trnL–F, | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | |||
| 1 | Lophozia dubia | n/c/n/c | 5.1 | 3.1 | 3.3 | 3.4 | 0.3 | 5.9 | 3.6 | 4.1 | 2.7 | 3.2 | 3.2 | 2.7 | 2.7 | 2.7 | 2.7 | |
| 2 | Lophozia ascendens | n/c/n/c | 6.9 | 2.7 | 4.3 | 3.0 | 6.4 | 4.5 | 3.6 | 4.3 | 2.7 | 2.7 | 3.5 | 2.7 | 2.7 | 2.7 | 2.7 | |
| 3 | Lophozia austrosibirica | n/c/n/c | 6.4 | 7.5 | 2.4 | 0.2 | 4.2 | 4.8 | 2.4 | 2.4 | 0.7 | 1.5 | 1.8 | 0.4 | 0.4 | 0.4 | 0.4 | |
| 4 | Lophozia fuscovirens | 0.4/0 | 6.8 | 8.1 | 7.5 | 2.4 | 4.0 | 4.7 | 2.3 | 2.5 | 2.3 | 2.4 | 2.4 | 1.9 | 1.9 | 1.9 | 1.9 | |
| 5 | Lophozia guttulata | n/c/n/c | 6.4 | 7.5 | 0 | 7.5 | 4.2 | 4.8 | 2.6 | 2.4 | 0.9 | 1.8 | 1.9 | 0.6 | 0.6 | 0.6 | 0.6 | |
| 6 | Lophozia koreana | 0.5/0.2 | 5.2 | 8.0 | 8.4 | 7.6 | 8.4 | 6.4 | 4.4 | 4.9 | 3.8 | 3.8 | 3.8 | 3.7 | 3.7 | 3.7 | 3.7 | |
| 7 | Lophozia lantratoviae | 0.8/0.2 | 5.9 | 5.8 | 6.1 | 6.8 | 6.1 | 6.2 | 5.9 | 4.9 | 4.7 | 4.4 | 5.7 | 4.6 | 4.8 | 4.6 | 4.6 | |
| 8 | Lophozia longiflora | n/c/n/c | 4.5 | 5.4 | 5.6 | 4.1 | 5.6 | 5.2 | 4.1 | 3.7 | 2.7 | 3.0 | 1.8 | 2.4 | 2.4 | 2.4 | 2.4 | |
| 9 | Lophozia pacifica | n/c/n/c | 6.4 | 7.3 | 2.2 | 7.4 | 2.2 | 7.8 | 5.8 | 5.2 | 2.3 | 1.6 | 3.3 | 2.0 | 2.0 | 2.0 | 2.0 | |
| 10 | Lophozia schusteriana | n/c/n/c | 7.1 | 7.4 | 1.5 | 7.5 | 1.5 | 7.9 | 5.9 | 5.4 | 2.3 | 1.3 | 2.0 | 0.2 | 0.2 | 0.2 | 0.2 | |
| 11 | Lophozia silvicola | —/n/c | — | — | — | — | — | — | — | — | — | — | 2.8 | 1.3 | 1.3 | 1.3 | 1.3 | |
| 12 | Lophozia silvicoloides | 0/0 | 4.8 | 5.5 | 6.3 | 5.5 | 6.3 | 5.6 | 4.6 | 2.8 | 5.6 | 5.8 | — | 1.8 | 1.8 | 1.8 | 1.8 | |
| 13 | Lophozia wenzelii | n/c/n/c | 7.6 | 8.2 | 1.7 | 8.3 | 1.7 | 8.8 | 6.4 | 6.1 | 2.5 | 1.5 | — | 6.6 | 0 | 0 | 0 | |
| 14 | Lophozia wenzelii var. lapponica | n/c/n/c | 6.9 | 7.3 | 0.9 | 7.4 | 0.9 | 8.2 | 5.7 | 5.4 | 1.7 | 0.9 | — | 5.8 | 0.9 | 0 | 0 | |
| 15 | Lophozia wenzelii var. litoralis | n/c/n/c | 6.9 | 7.8 | 0.9 | 7.8 | 0.9 | 8.2 | 6.1 | 5.5 | 2.2 | 1.7 | — | 6.2 | 1.9 | 1.0 | 0 | |
| 16 | Lophozia wenzelii var. massularioides | n/c/n/c | 6.9 | 7.8 | 1.3 | 7.8 | 1.3 | 8.5 | 5.8 | 5.4 | 2.2 | 1.1 | — | 6.0 | 1.0 | 0.5 | 1.4 | |
| Specimen | Specimen Voucher | GenBank Accession Number | |
|---|---|---|---|
| ITS1–2 | trnL-trnF | ||
| Lophozia dubia Schiffn. | China, Yunnan Province, Diqing Prefecture, V.A. Bakalin & W.Z. Ma, C-83-7-18 (VBGI) | OR982393 | OR995729 |
| Lophozia ascendens (Warnst.) R.M. Schust. | Russia, Buryatiya, N.A. Konstantinova, 109-3-01 (KPABG) | DQ875089 | DQ875054 |
| Lophozia austrosibirica Bakalin | Russia, Buryatiya, V.A. Bakalin, 15-9-99 (KPABG) | DQ875105 | DQ875069 |
| Lophozia fuscovirens Bakalin & Vilnet | Russia, Magadan Prov., V.A. Bakalin, Mag-28-32-13 (KPABG) | MK007091 | MK012211 |
| Lophozia fuscovirens Bakalin & Vilnet | Russia, Magadan Prov., V.A. Bakalin, Mag-50-16-11 (KPABG) | MK007092 | MK012212 |
| Lophozia fuscovirens Bakalin & Vilnet | Russia, Magadan Prov., V.A. Bakalin, Mag-30-4-14 (KPABG) | MK007093 | MK012213 |
| Lophozia fuscovirens Bakalin & Vilnet | Norway, Svalbard, A. Savchenko, CA16-12-1c | MK774737 | MK779914 |
| Lophozia guttulata (Lindb. & Arnell) A. Evans | Russia, Buryatiya, N.A. Konstantinova, 81-1-01 (KPABG) | DQ875108 | DQ875072 |
| Lophozia koreana (Bakalin, S.S. Choi & B.Y. Sun) Maltseva, Vilnet & Bakalin | South Korea, Jeollabuk-do, V.A. Bakalin & S.S. Choi, Kor-74-5-19 (VGBI) | MW685441 | MW654174 |
| Lophozia koreana (Bakalin, S.S. Choi & B.Y. Sun) Maltseva, Vilnet & Bakalin | South Korea, Jeollabuk-do, V.A. Bakalin & S.S. Choi, Kor-75-15-19 (VGBI) | MW685442 | MW654175 |
| Lophozia koreana (Bakalin, S.S. Choi & B.Y. Sun) Maltseva, Vilnet & Bakalin | South Korea, Jeollabuk-do, V.A. Bakalin & S.S. Choi, Kor-76-1-19 (VGBI) | MW685443 | MW654176 |
| Lophozia lantratoviae Bakalin | Russia, Buryatiya, V.A. Bakalin, 76-7-01 (KPABG) | DQ875090 | DQ875055 |
| Lophozia lantratoviae Bakalin | China, Sichuan Prov., V.A. Bakalin, China-35-9-17, 37536 (VBGI) | MK007086 | MK012206 |
| Lophozia lantratoviae Bakalin | China, Sichuan Prov., V.A. Bakalin, China-39-3-17, 37283 (VBGI) | MK007087 | MK012207 |
| Lophozia lantratoviae Bakalin | Russia, Primorsky Terr., V.A. Bakalin, P-44-20-10 (KPABG) | MK007088 | MK012208 |
| Lophozia lantratoviae Bakalin | Russia, North Ossetia—Alanya Rep., A.V. Rumyantseva, 123153 (KPABG) | MW872337 | MW887533 |
| Lophozia longiflora (Nees) Schiffn. | Russia, Chitinskaya Prov, V.A. Bakalin, 11-5-00 (KPABG) | DQ875101 | DQ875066 |
| Lophozia pacifica Bakalin | Russia, Kamchatka Terr., V.A. Bakalin, K-16-2-02-VB, 103630 (KPABG) | MK007089 | MK012209 |
| Lophozia schusteriana Schljakov | Russia, Murmanskaya Prov, V.A. Bakalin, G9331 (KPABG) | DQ875103 | DQ875067 |
| Lophozia silvicola H. Buch | Russia, Sakhalin Prov., Sakhalin Isl., V.A. Bakalin, S-14-4-17 (VBGI) | MK007090 | MK012210 |
| Lophozia silvicoloides N. Kitag. | Russia, Kamchatskaya Prov, V.A. Bakalin, K-57-23-02-VB (KPABG) | DQ875098 | DQ875063 |
| Lophozia silvicoloides N. Kitag. | Russia, Murmanskaya Prov, N.A. Konstantinova, 356-4-00 (KPABG) | DQ875099 | DQ875064 |
| Lophozia silvicoloides N. Kitag. | Norway, Svalbard, N.A. Konstantinova, K241-1b-12 (KPABG) | MW298767 | MW297148 |
| Lophozia wenzelii (Nees) Steph. | Russia, Murmanskaya Prov, N.A. Konstantinova, 9329 (KPABG) | DQ875109 | DQ875073 |
| Lophozia wenzelii var. lapponica H. Buch & S.W. Arnell | Svalbard, Spitsbergen, N.A. Konstantinova, 124-2-04 (KPABG) | DQ875112 | DQ875076 |
| Lophozia wenzelii var. litoralis (Arnell) Bakalin | Russia, Murmanskaya Prov, V.A. Bakalin, 12-3-02 (KPABG) | DQ875110 | DQ875074 |
| Lophozia wenzelii var. massularioides Bakalin | Russia, Caucasus, Onipchenko, 31.08.83 (MHA) | DQ875111 | DQ875075 |
| Lophoziopsis excisa (Dicks.) Konstant. & Vilnet | Svalbard, Spitsbergen, N.A. Konstantinova, 104-1-04 (KPABG) | DQ875091 | DQ875056 |
| Lophoziopsis excisa (Dicks.) Konstant. & Vilnet | Russia, Murmanskaya Prov, N.A. Konstantinova, 41-2-97 (KPABG) | DQ875092 | DQ875057 |
| Lophoziopsis excisa (Dicks.) Konstant. & Vilnet | Svalbard, Spitsbergen, N.A. Konstantinova, K-21-2-05 (KPABG) | DQ875093 | DQ875058 |
| Lophoziopsis excisa (Dicks.) Konstant. & Vilnet | Russia, Maryi-El, N.A. Konstantinova, K437-2-04 (KPABG) | EF065691 | EF065684 |
| Lophoziopsis excisa (Dicks.) Konstant. & Vilnet | China, Sichuan Prov., V.A. Bakalin, China-37-2-17, 37462 (VBGI) | MK007094 | MK012214 |
| Lophoziopsis excisa (Dicks.) Konstant. & Vilnet | Norway, Svalbard, A. Savchenko, CA 364-2a-11 (KPABG) | MW298770 | MW297151 |
| Lophoziopsis longidens (Lindb.) Konstant. & Vilnet | Russia, Murmanskaya Prov, N.A. Konstantinova, 360-2-00 (KPABG) | DQ875094 | DQ875059 |
| Lophoziopsis pellucida (R.M. Schust.) Konstant. & Vilnet | Russia, Komi, M. Dulin, 103640 (KPABG) | EF065694 | EF065686 |
| Lophoziopsis pellucida (R.M. Schust.) Konstant. & Vilnet | Russia, Murmanskaya Prov., N.A. Konstantinova, 39-2a-03 (KPABG) | EF065695 | EF065687 |
| Lophoziopsis pellucida (R.M. Schust.) Konstant. & Vilnet | Russia, Magadan Prov., V.A. Bakalin, Mag-44-37-11, 115673 (KPABG) | MK007097 | MK012217 |
| Lophoziopsis pellucida (R.M. Schust.) Konstant. & Vilnet | Russia, Trans-Baikal Terr., Yu.S. Mamontov, 356-3-6 (KPABG) | MW298773 | MW297154 |
| Lophoziopsis polaris (R.M. Schust.) Konstant. & Vilnet | Russia, Magadan Prov., V.A. Bakalin, Mag-57-16-11, 115699 (KPABG) | MK007098 | MK012218 |
| Lophoziopsis polaris (R.M. Schust.) Konstant. & Vilnet | Norway, Svalbard, Konstantinova N.A., Savchenko A.N., K129-07 (KPABG) | MT334459 | MT338482 |
| Lophoziopsis polaris (R.M. Schust.) Konstant. & Vilnet | Norway, Svalbard, A.N. Savchenko, CA 19-29-3 (KPABG) | MW298774 | MW297155 |
| Lophoziopsis polaris var. sphagnorum (R.M. Schust.) Konstant. & Vilnet | Russia, Yakutiya, V.A. Bakalin, 23-11-00 (KPABG) | DQ875096 | DQ875061 |
| Lophoziopsis propagulifera (Gottsche) Konstant. & Vilnet | Russia, Kamchatskaya Prov, V.A. Bakalin, K-53-6-02-VB (KPABG) | DQ875097 | DQ875062 |
| Obtusifolium obtusum (Lindb.) S.W. Arnell | Sw-48-30-13 (VGBI) | MT504415 | MT476337 |
| Trilophozia quinquedentata (Huds.) Bakalin | Russia, Kareliya, Bakalin 2 July 1997 | EU791804 | AY327786 |
| Trilophozia quinquedentata f. gracilis (R.M. Schust.) Konstant. | Svalbard, North-East Land, N.A. Konstantinova, K 118-2-06 (KPABG) | EU791802 | EU791684 |
| Trilophozia quinquedentata f. gracilis (R.M. Schust.) Konstant. | Svalbard, North-East Land, N.A. Konstantinova, K 72-2-06 (KPABG) | EU791803 | EU791685 |
| Trilophozia quinquedentata (Huds.) Bakalin | Norway, Svalbard, Konstantinova N.A., Savchenko A.N., K70-08 (KPABG) | MT334460 | MT338484 |
| Tritomaria exsectiformis (Breidl.) Schiffn. ex Loeske | Russia, Rep. Buryatiya, N.A. Konstantinova, 83-4-01 (KPABG) | EU791801 | EU791683 |
| Tritomaria scitula (Taylor) Jørg. | Russia, Komi Rep., M. Dulin, N.A. Konstantinova, G 101301 (KPABG) | EU791799 | EU791681 |
| Locus | Sequence (5′-3′) | Direction | Annealing Temperature (°C) | Reference |
|---|---|---|---|---|
| trnL–F cpDNA | CGAAATTGGTAGACGCTGCG | forward | 62 | [20] |
| trnL–F cpDNA | TGCCAGAAACCAGATTTGAAC | reverse | 58 | [20] |
| ITS 1–2 nrDNA | ACCTGCGGAAGGATCATTG | forward | 58 | [37] |
| ITS 1–2 nrDNA | GATATGCTTAAACTCAGCGG | reverse | 58 | [38] |
| Initial denaturation | 3 min—94 °C | |
| Denaturation | 30 s—95 °C | |
| Annealing | 20 s (trnL–F), 30 s (ITS 1–2) | 35 cycles |
| at 58 °C (trnL–F), 60 °C (ITS 1–2) | ||
| Elongation | 30 s—72 °C | |
| Final elongation | 3 min—72 °C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakalin, V.A.; Maltseva, Y.D.; Troitsky, A.V. The Position of Lophozia dubia (Lophoziaceae, Marchantiophyta) in the Phylogenetic System of Lophozia and the Distribution of Lophozia in Southeast Eurasia, Extending to Indonesia. Plants 2024, 13, 367. https://doi.org/10.3390/plants13030367
Bakalin VA, Maltseva YD, Troitsky AV. The Position of Lophozia dubia (Lophoziaceae, Marchantiophyta) in the Phylogenetic System of Lophozia and the Distribution of Lophozia in Southeast Eurasia, Extending to Indonesia. Plants. 2024; 13(3):367. https://doi.org/10.3390/plants13030367
Chicago/Turabian StyleBakalin, Vadim A., Yulia D. Maltseva, and Aleksey V. Troitsky. 2024. "The Position of Lophozia dubia (Lophoziaceae, Marchantiophyta) in the Phylogenetic System of Lophozia and the Distribution of Lophozia in Southeast Eurasia, Extending to Indonesia" Plants 13, no. 3: 367. https://doi.org/10.3390/plants13030367