
Grazing Intensity Alters Leaf and Spike Photosynthesis, Transpiration, and Related Parameters of Three Grass Species on an Alpine Steppe in the Qilian Mountains

Abstract
:1. Introduction
2. Results
2.1. Environmental Parameters
2.2. Photosynthesis Parameters
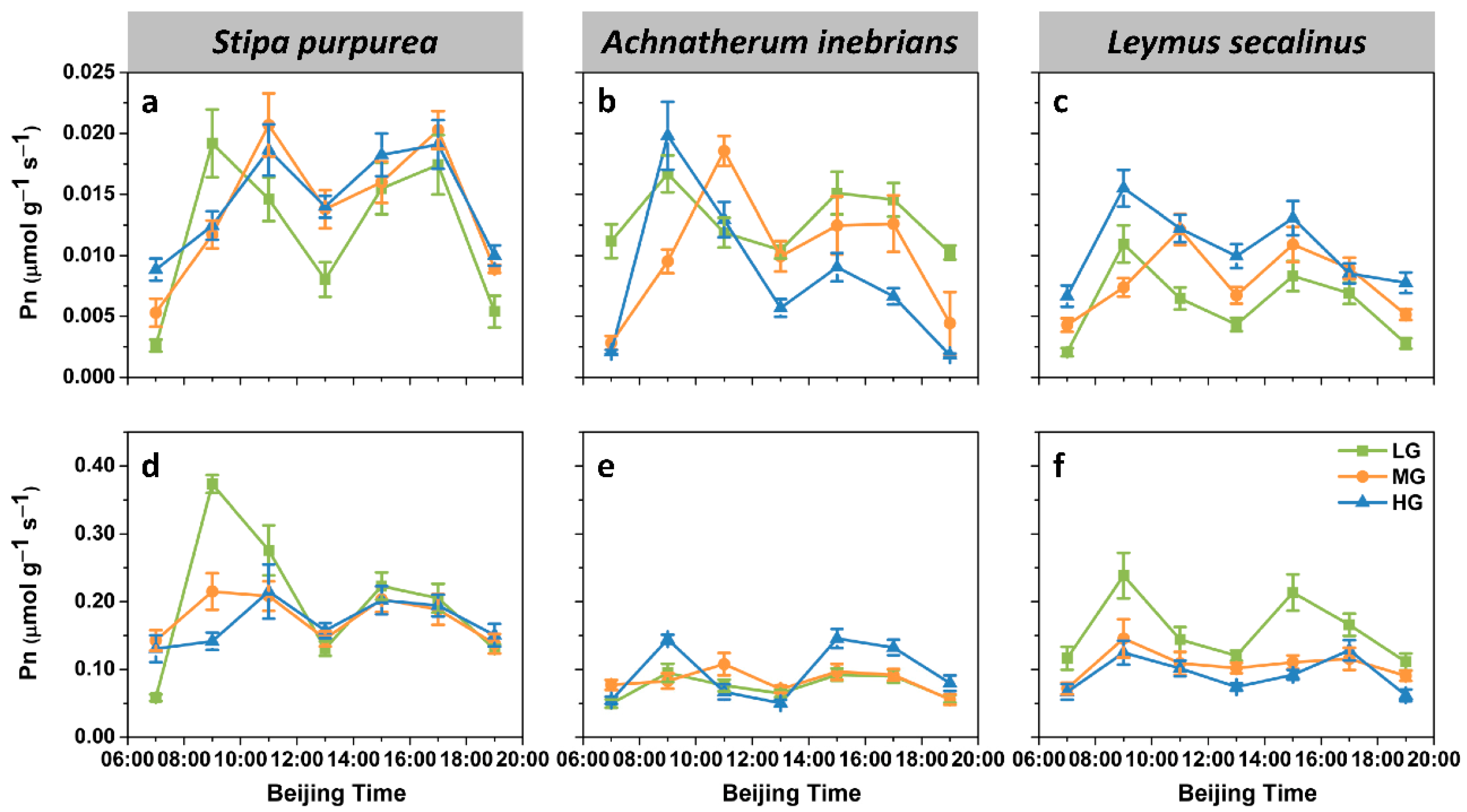
2.2.1. Net Photosynthetic Rate
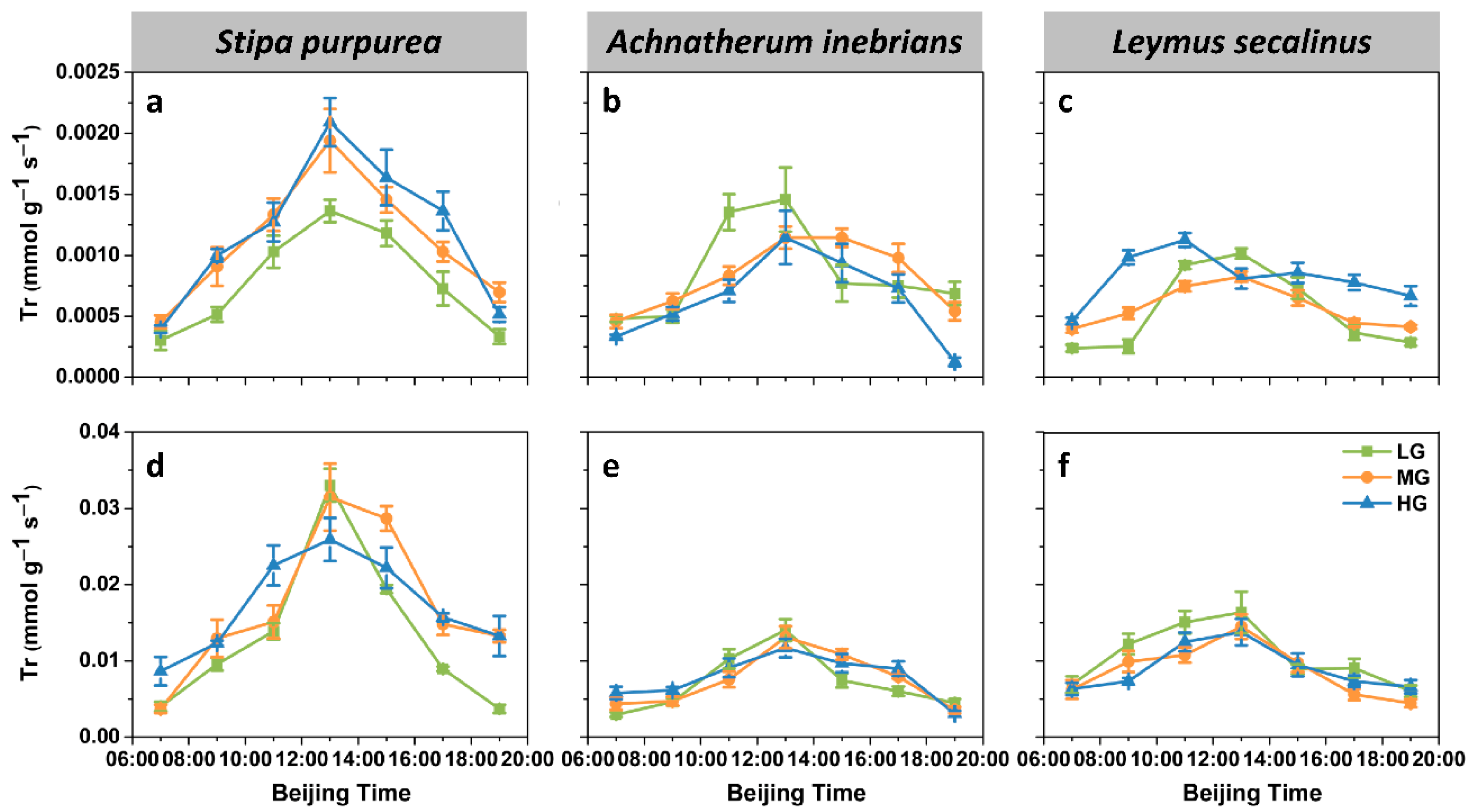
2.2.2. Transpiration Rate
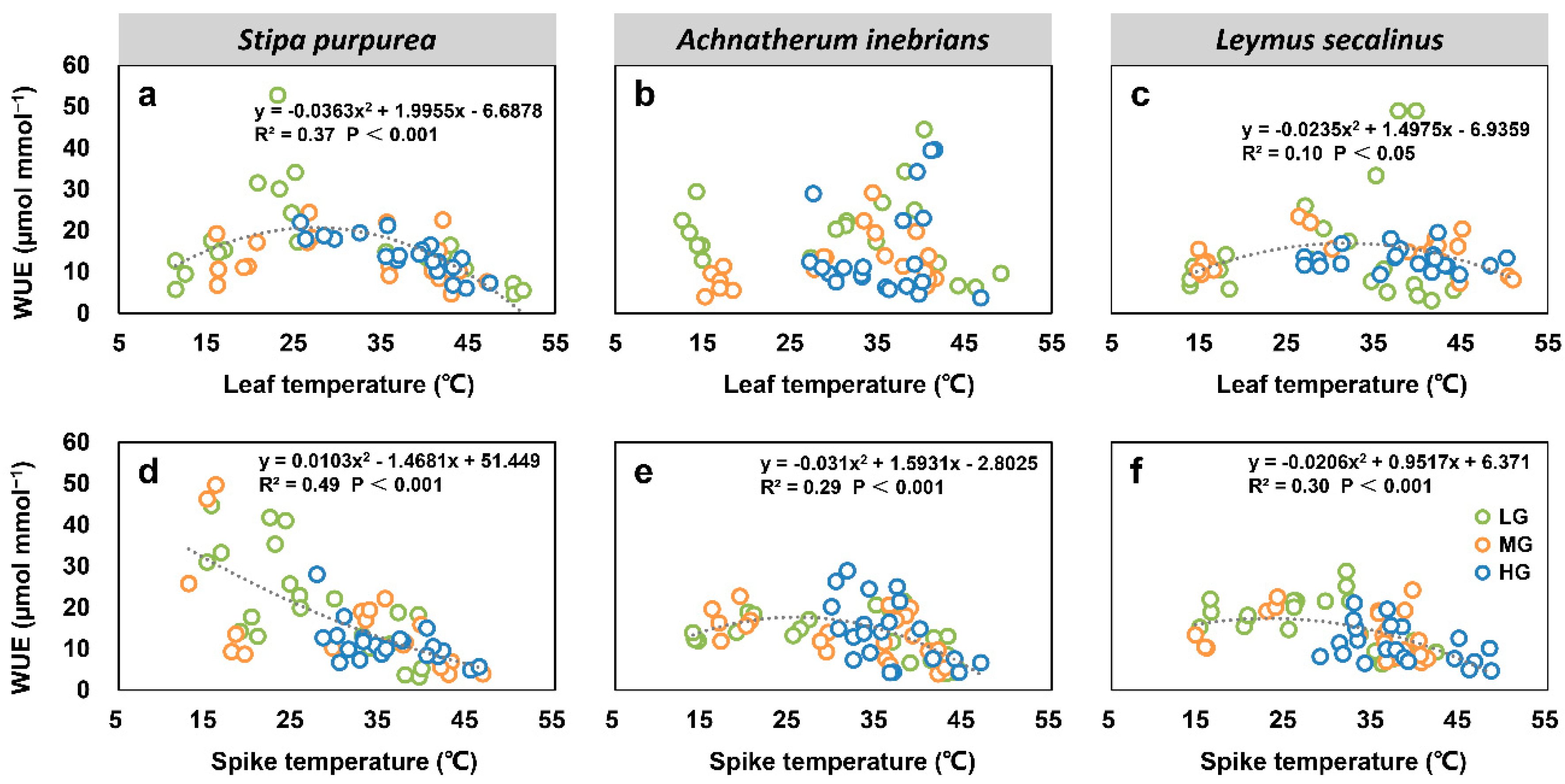
2.2.3. Water Use Efficiency
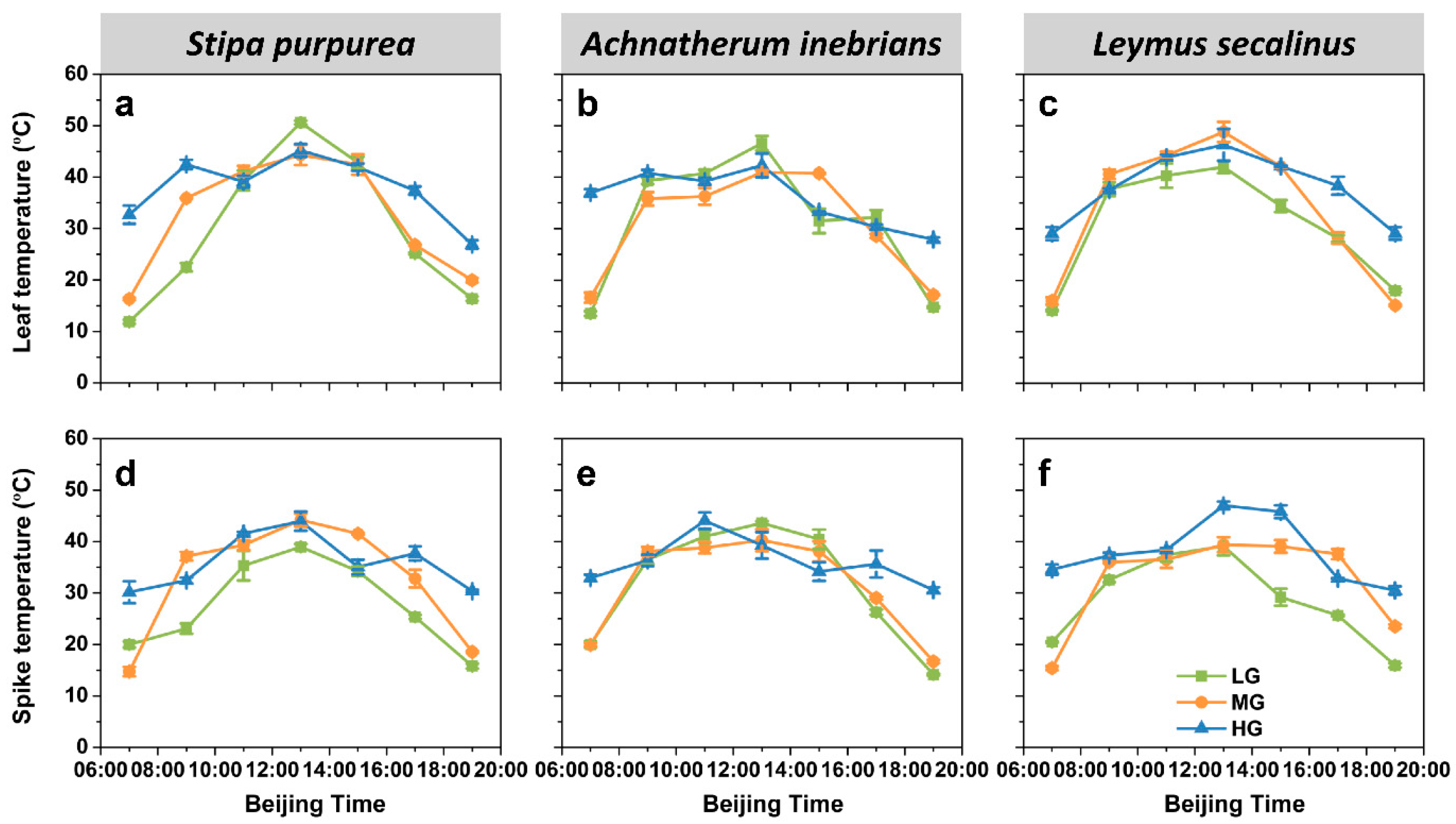
2.2.4. Organ Temperature
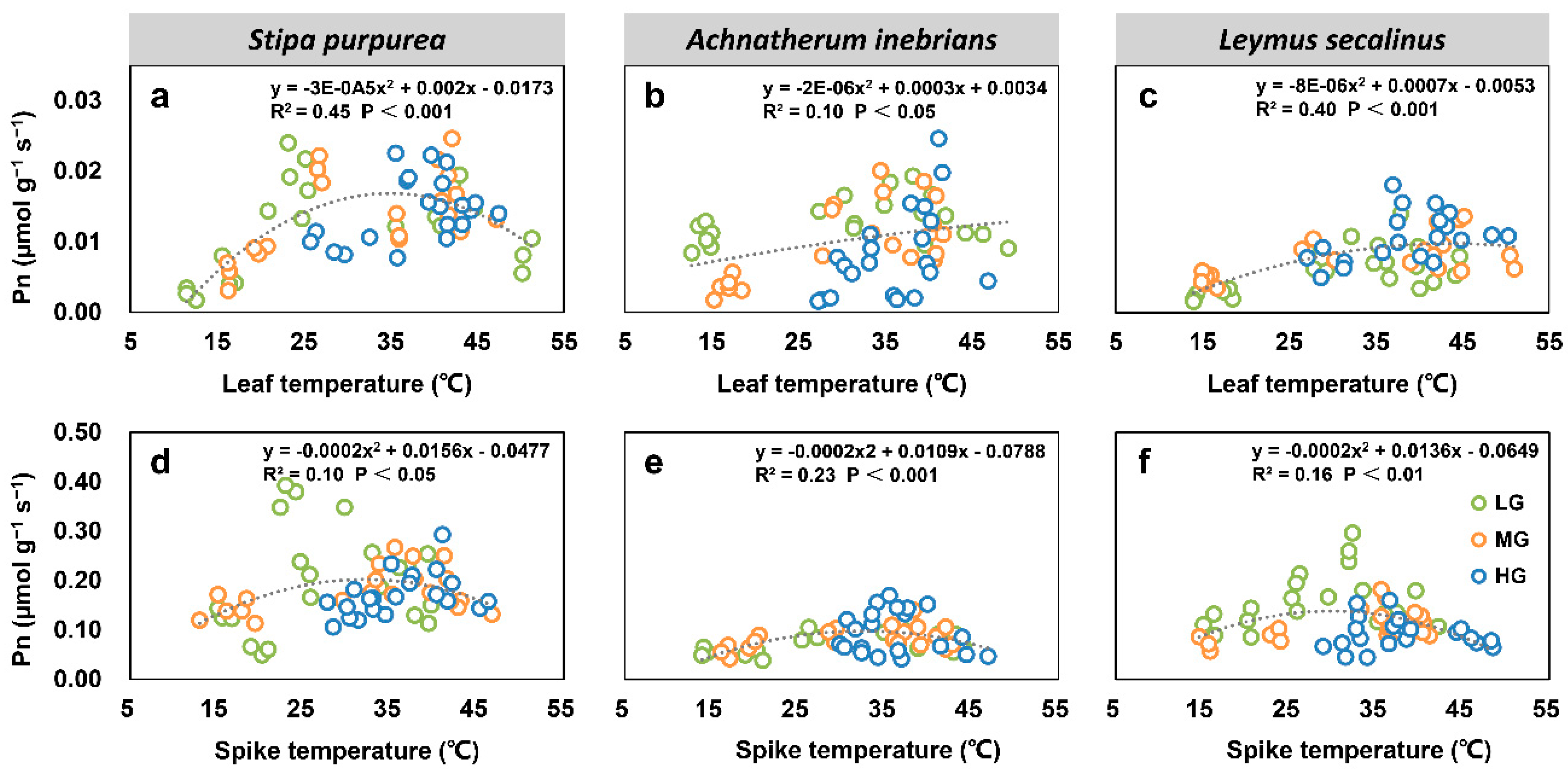
2.2.5. Correlations between Net Photosynthetic Rate, Transpiration Rate, Water Use Efficiency, and Organ Temperature under Grazing Intensity
2.3. Chlorophyll Content
2.3.1. Chlorophyll a, Chlorophyll b and Total Chlorophyll
2.3.2. Total Carotenoid Content, the chl a/chl b Ratio and TC/Total Carotenoid Ratio
3. Discussion
3.1. Effect of Grazing Intensity on Photosynthesis Parameters
3.2. Effect of Grazing Intensity on Photosynthetic Pigments
4. Materials and Methods
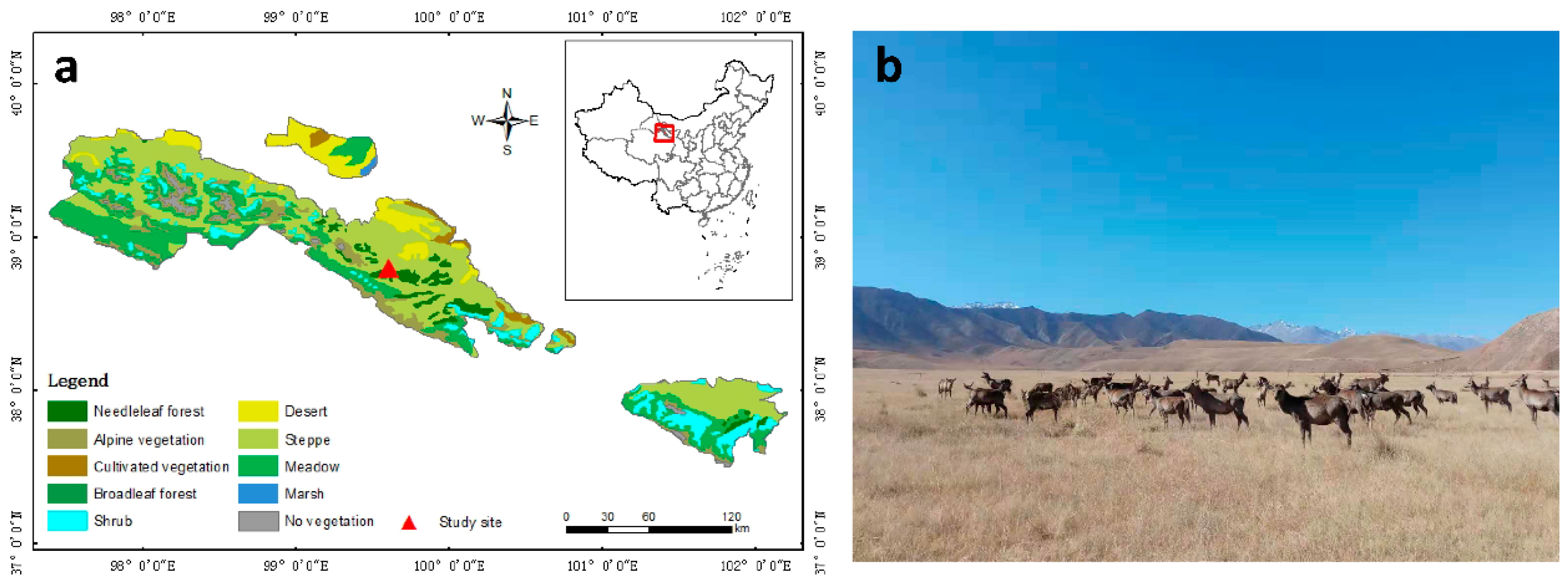
4.1. Study area and Experimental Design
4.2. Photosynthesis Parameters and Photosynthetic Pigments
4.3. Data and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Luo, C.; Xu, G.; Chao, Z.; Wang, S.; Lin, X.; Hu, Y.; Zhang, Z.; Duan, J.; Chang, X.; Su, A.; et al. Effect of warming and grazing on litter mass loss and temperature sensitivity of litter and dung mass loss on the Tibetan plateau. Glob. Chang. Biol. 2010, 16, 1606–1617. [Google Scholar] [CrossRef]
- Sala, O.E.; Yahdjian, L.; Havstad, K.; Aguiar, M.R. Rangeland ecosystem services: Nature’s supply and humans’ demand. In Rangeland Systems; Springer: Cham, Switzerland, 2017; pp. 467–489. [Google Scholar]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Guanter, L.; Zhang, Y.; Jung, M.; Joiner, J.; Voigt, M.; Berry, J.A.; Frankenberg, C.; Huete, A.R.; Zarco-Tejada, P.; Lee, J.E.; et al. Global and time-resolved monitoring of crop photosynthesis with chlorophyll fluorescence. Proc. Natl. Acad. Sci. USA 2014, 111, E1327–E1333. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.; Lan, Z.; Li, W.; Shao, R.; Shan, Y.; Wan, H.; Taube, F.; Bai, Y. Differential responses of plant functional trait to grazing between two contrasting dominant C3 and C4 species in a typical steppe of Inner Mongolia, China. Plant Soil 2011, 340, 141–155. [Google Scholar] [CrossRef]
- Xiao, H.; Peng, Z.; Xu, C.; Zhang, D.; Chai, J.; Pan, T.; Yu, X. Yak and Tibetan sheep trampling inhibit reproductive and photosynthetic traits of medicago ruthenica var. inschanica. Environ. Monit. Assess 2018, 190, 507. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, Y.; Hou, F.; Millner, J.; Wang, Z.; Chang, S. Grazing activity increases decomposition of yak dung and litter in an alpine meadow on the Qinghai-Tibet plateau. Plant Soil 2019, 444, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Huang, Q.; Mi, X.; Bai, Y.; Zhang, M.; Li, X. Grazing every month minimizes size but boosts photosynthesis in stipa grandis in the steppe of inner Mongolia, China. J. Arid Land 2018, 10, 601–611. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zang, R.; Chen, H. Effects of grazing on photosynthetic features and soil respiration of rangelands in the Tianshan Mountains of Northwest China. Sci. Rep. 2016, 6, 30087. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Dong, S.; Li, S.; Xiao, J.; Han, Y.; Yang, M.; Zhang, J.; Gao, X.; Xu, Y.; Li, Y.; et al. Grazing enhances plant photosynthetic capacity by altering soil nitrogen in alpine grasslands on the Qinghai-Tibetan plateau. Agric. Ecosyst. Environ. 2019, 280, 161–168. [Google Scholar] [CrossRef]
- Lu, Z.; Pan, Y.; Hu, W.; Cong, R.; Ren, T.; Guo, S.; Lu, J. The photosynthetic and structural differences between leaves and siliques of Brassica napus exposed to potassium deficiency. BMC Plant Biol. 2017, 17, 240. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Zhang, L.; Kuang, C.; Guo, Y.; Huang, C.; Deng, L.; Sun, X.; Zhan, G.; Hu, Z.; Wang, H.; et al. Important photosynthetic contribution of silique wall to seed yield-related traits in Arabidopsis thaliana. Photosynth. Res. 2018, 137, 493–501. [Google Scholar] [CrossRef]
- Hu, L.; Zhang, Y.; Xia, H.; Fan, S.; Song, J.; Lv, X.; Kong, L. Photosynthetic characteristics of non-foliar organs in main C3 cereals. Physiol. Plant. 2019, 166, 226–239. [Google Scholar] [CrossRef]
- Tambussi, E.A.; Bort, J.; Guiamet, J.J.; Nogués, S.; Araus, J.L. The photosynthetic role of ears in C3 cereals: Metabolism, water use efficiency and contribution to grain yield. Crit. Rev. Plant Sci. 2007, 26, 1–16. [Google Scholar] [CrossRef]
- Simkin, A.J.; Faralli, M.; Ramamoorthy, S.; Lawson, T. Photosynthesis in non-foliar tissues: Implications for yield. Plant J. 2019, 101, 1001–1015. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Bragado, R.; Molero, G.; Reynolds, M.P.; Araus, J.L. Photosynthetic contribution of the ear to grain filling in wheat: A comparison of different methodologies for evaluation. J. Exp. Bot. 2016, 67, 2787–2798. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, Y.; Luo, H.; Li, W.; Oguchi, R.; Fan, D.; Chow, W.; Zhang, W. Important photosynthetic contribution from the non-foliar green organs in cotton at the late growth stage. Planta 2012, 235, 325–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brazel, A.J.; Ó’Maoiléidigh, D.S. Photosynthetic activity of reproductive organs. J. Exp. Bot. 2019, 70, 1737–1754. [Google Scholar] [CrossRef] [PubMed]
- Ziegler-Jöns, A. Gas exchange of ears of cereals in response to carbon dioxide and light: I. Relative contributions of parts of the ears of wheat, oat, and barley to the gas exchange of the whole organ. Planta 1989, 178, 84–91. [Google Scholar] [CrossRef]
- Šebela, D.; Quiñones, C.; Olejníčková, J.; Jagadish, K.S.V. Temporal chlorophyll fluorescence signals to track changes in optical properties of maturing rice panicles exposed to high night temperature. Field Crops Res. 2015, 177, 75–85. [Google Scholar] [CrossRef]
- Jia, S.; Lv, J.; Jiang, S.; Liang, T.; Liu, C.; Jing, Z. Response of wheat ear photosynthesis and photosynthate carbon distribution to water deficit. Photosynthetica 2015, 53, 95–109. [Google Scholar] [CrossRef] [Green Version]
- Hein, J.A.; Sherrard, M.E.; Manfredi, K.P.; Abebe, T. The fifth leaf and spike organs of barley (Hordeum vulgare L.) display different physiological and metabolic responses to drought stress. BMC Plant Biol. 2016, 16, 248. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.; Yang, T.; Zeng, B.; Zhu, X.; Xu, H. Vegetation cover variation in the Qilian Mountains and its response to climate change in 2000–2011. J. Mt. Sci. 2013, 10, 1050–1062. [Google Scholar] [CrossRef]
- Cheng, G.; Li, X.; Zhao, W.; Xu, Z.; Feng, Q.; Xiao, S.; Xiao, H. Integrated study of the water–ecosystem–economy in the Heihe River Basin. Natl. Sci. Rev. 2014, 1, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Baranova, A.; Schickhoff, U.; Wang, S.; Jin, M. Mountain pastures of Qilian Shan: Plant communities, grazing impact and degradation status (Gansu province, NW China). Hacquetia 2016, 15, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Sun, F.; Lü, Y.; Wang, J.; Hu, J.; Fu, B. Soil moisture dynamics of typical ecosystems in response to precipitation: A monitoring-based analysis of hydrological service in the Qilian Mountains. Catena 2015, 129, 63–75. [Google Scholar] [CrossRef]
- Yao, Z.; Zhao, C.; Yang, K.; Liu, W.; Li, Y.; You, J.; Xiao, J. Alpine grassland degradation in the Qilian Mountains, China—A case study in Damaying Grassland. Catena 2016, 137, 494–500. [Google Scholar] [CrossRef]
- Yuan, H.; Hou, F. Grazing intensity and soil depth effects on soil properties in alpine meadow pastures of Qilian Mountain in northwest China. Acta Agric. Scand. B Soil Plant Sci. 2015, 65, 222–232. [Google Scholar] [CrossRef] [Green Version]
- Hou, F.; Ren, J. Trampling of Gansu Wapiti (Cervus elaphus kansuensis Pocock) and its effects on soil properties in winter grazing grassland. Acta Ecol. Sin. 2003, 23, 486–494. [Google Scholar]
- Geiger, D.R.; Servaites, J.C. Diurnal regulation of photosynthetic carbon metabolism in C3 plants. Annu. Rev. Plant Biol. 1994, 45, 235–256. [Google Scholar] [CrossRef]
- Gao, Z.; Xu, B.; Wang, J.; Huo, L.; Li, S. Diurnal and seasonal variations in photosynthetic characteristics of switchgrass in semiarid region on the Loess Plateau of China. Photosynthetica 2015, 53, 489–498. [Google Scholar] [CrossRef]
- Zhang, X.; Lang, D.; Zhang, E.; Bai, C.; Wang, H. Diurnal changes in photosynthesis and antioxidants of Angelica sinensis as influenced by cropping systems. Photosynthetica 2013, 51, 252–258. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Jaafar, H.Z.E. Photosynthetic capacity, photochemical efficiency and chlorophyll content of three varieties of Labisia pumila Benth. exposed to open field and greenhouse growing conditions. Acta Physiol. Plant 2011, 33, 2179–2185. [Google Scholar] [CrossRef] [Green Version]
- Yin, C.; Berninger, F.; Li, C. Photosynthetic responses of Populus przewalski subjected to drought stress. Photosynthetica 2006, 44, 62–68. [Google Scholar] [CrossRef]
- Detling, J.K.; Dyer, M.I.; Winn, D.T. Net photosynthesis, root respiration, and regrowth of Bouteloua gracilis following simulated grazing. Oecologia 1979, 41, 127–134. [Google Scholar] [CrossRef]
- Castro, H.; Nabais, C.; Alados, C.L.; Freitas, H. Effects of cessation of grazing on leaf-level photosynthesis of Periploca laevigata. Appl. Veg. Sci. 2003, 6, 255–260. [Google Scholar] [CrossRef]
- Redondo-Gómez, S.; Mancilla-Leytón, J.M.; Mateos-Naranjo, E.; Cambrollé, J.; Martín-Vicente, A. Differential photosynthetic performance of three Mediterranean shrubs under grazing by domestic goats. Photosynthetica 2010, 48, 348–354. [Google Scholar] [CrossRef]
- Harrison, M.T.; Kelman, W.M.; Moore, A.D.; Evans, J.R. Grazing winter wheat relieves plant water stress and transiently enhances photosynthesis. Funct. Plant Biol. 2010, 37, 726–736. [Google Scholar] [CrossRef]
- Nowak, R.; Caldwell, M. A test of compensatory photosynthesis in the field: Implications for herbivory tolerance. Oecologia 1984, 61, 311–318. [Google Scholar] [CrossRef]
- Irisarri, J.G.N.; Derner, J.D.; Porensky, L.M.; Augustine, D.J.; Reeves, J.; Mueller, K.E. Grazing intensity differentially regulates ANPP response to precipitation in North American semiarid grasslands. Ecol. Appl. 2016, 26, 1370–1380. [Google Scholar] [CrossRef] [Green Version]
- Miles, C.O.; Lane, G.A.; di Menna, M.E.; Garthwaite, I.; Piper, E.L.; Ball, O.J.P.; Latch, G.C.M.; Allen, J.M.; Hunt, M.B.; Bush, L.P.; et al. High levels of ergonovine and lysergic acid amide in toxic Achnatherum inebrians accompany infection by an Acremonium-like endophytic fungus. J. Agric. Food Chem. 1996, 44, 1285–1290. [Google Scholar] [CrossRef]
- Shi, Z.C. Important Poisonous Plants of China Grassland; China Agriculture Press: Beijing, China, 1997. [Google Scholar]
- Liang, Y.; Wang, H.; Li, C.; Nan, Z.; Li, F. Effects of feeding drunken horse grass infected with Epichloë gansuensis endophyte on animal performance, clinical symptoms and physiological parameters in sheep. BMC Vet. Res. 2017, 13, 223. [Google Scholar] [CrossRef] [Green Version]
- Gaffney, R.; Porensky, L.M.; Gao, F.; Irisarri, J.G.; Durante, M.; Derner, J.D.; Augustine, D.J. Using APAR to Predict Aboveground Plant Productivity in Semi-Arid Rangelands: Spatial and Temporal Relationships Differ. Remote Sens. 2018, 10, 1474. [Google Scholar] [CrossRef] [Green Version]
- Pieruschka, R.; Huber, G.; Berry, J.A. Control of transpiration by radiation. Proc. Natl. Acad. Sci. USA 2010, 107, 13372–13377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Short, T.; Fox, R.; Bauerle, W. Transpiration, leaf temperature and stomatal resistance of a greenhouse cucumber crop. Agric. For. Meteorol. 1990, 51, 197–209. [Google Scholar] [CrossRef]
- O’Sullivan, O.S.; Heskel, M.A.; Reich, P.B.; Tjoelker, M.G.; Weerasinghe, K.W.L.K.; Penillard, A.; Zhu, L.L.; Egerton, J.J.G.; Bloomfield, K.J.; Creek, D.; et al. Thermal limits of leaf metabolism across biomes. Glob. Chang. Biol. 2017, 23, 209–223. [Google Scholar] [CrossRef] [Green Version]
- Ilnitsky, O.A.; Pashtetsky, A.V.; Plugatar, Y.V.; Korsakova, S.P. Dependency of a Photosynthesis Rate in Nerium oleander L. on Environmental Factors, Leaf Temperature, Transpiration, and Their Change During Vegetation in Subtropics. Russ. Agric. Sci. 2018, 44, 224–228. [Google Scholar] [CrossRef]
- Sakata, T.; Nakano, T.; Kachi, N. Effects of internal conductance and Rubisco on the optimum temperature for leaf photosynthesis in Fallopia japonica growing at different altitudes. Ecol. Res. 2015, 30, 163–171. [Google Scholar] [CrossRef]
- Shen, X.; Liu, B.; Zhou, D. Effects of grassland degradation on air and soil temperature in Songnen plain of northeast China. Curr. Sci. India 2015, 108, 2098–2101. [Google Scholar]
- Ferrante, P.; Gennusa, M.; Peri, G.; Rizzo, G.; Scaccianoce, G. Vegetation growth parameters and leaf temperature: Experimental results from a six plots green roofs’ system. Energy 2016, 115, 1723–1732. [Google Scholar] [CrossRef]
- Monteiro, M.V.; Blanuša, T.; Verhoef, A.; Hadley, P.; Cameron, R.W.F. Relative importance of transpiration rate and leaf morphological traits for the regulation of leaf temperature. Aust. J. Bot. 2016, 64, 32–44. [Google Scholar] [CrossRef]
- Urban, J.; Ingwers, M.W.; McGuire, M.A.; Teskey, R.O. Increase in leaf temperature opens stomata and decouples net photosynthesis from stomatal conductance in Pinus taeda and Populus deltoides x nigra. J. Exp. Bot. 2017, 68, 1757–1767. [Google Scholar] [CrossRef]
- Gajanayake, B.; Reddy, K.R.; Shankle, M.W.; Arancibia, R.A. Growth, developmental, and physiological responses of two sweetpotato (Ipomoea batatas L. [Lam]) cultivars to early season soil moisture deficit. Sci. Hortic. 2014, 168, 218–228. [Google Scholar] [CrossRef]
- Yan, C.; Chen, H.; Fan, T.; Huang, Y.; Yu, S.; Chen, S.; Hong, X. Rice flag leaf physiology, organ and canopy temperature in response to water stress. Plant Prod. Sci. 2012, 15, 92–99. [Google Scholar] [CrossRef]
- Nourtier, M.; Chanzy, A.; Cailleret, M.; Yingge, X.; Huc, R.; Davi, H. Transpiration of silver Fir (Abies alba mill.) during and after drought in relation to soil properties in a Mediterranean mountain area. Ann. For. Sci. 2014, 71, 683–695. [Google Scholar] [CrossRef]
- Sun, D.; Liddle, M.J. Plant morphological characteristics and resistance to simulated trampling. Environ. Manag. 1993, 17, 511–521. [Google Scholar] [CrossRef]
- Trenholm, L.E.; Duncan, R.R.; Carrow, R.N. Wear tolerance, shoot performance, and spectral reflectance of seashore paspalum and bermudagrass. Crop Sci. 1999, 39, 1147–1152. [Google Scholar] [CrossRef]
- Xu, L.; Yu, F.; Drunen, V.E.; Schieving, F.; Dong, M.; Anten, N.P.R. Trampling, defoliation and physiological integration affect growth, morphological and mechanical properties of a root-suckering clonal tree. Ann. Bot. 2012, 109, 1001–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Freitas, S.M.A.; Yu, F.; Dong, M.; Anten, N.P.R.; Werger, M.J.A. Effects of trampling on morphological and mechanical traits of dryland shrub species do not depend on water availability. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faralli, M.; Matthews, J.; Lawson, T. Exploiting natural variation and genetic manipulation of stomatal conductance for crop improvement. Curr. Opin. Plant Biol. 2019, 49, 1–7. [Google Scholar] [CrossRef]
- Araus, J.L.; Brown, H.R.; Febrero, A.; Bort, J.; Serret, M.D. Ear photosynthesis, carbon isotope discrimination and the contribution of respiratory CO2 to differences in grain mass in durum wheat. Plant Cell Environ. 1993, 16, 383–392. [Google Scholar] [CrossRef]
- Ye, X.; Yu, F.; Dong, M. A trade-off between guerrilla and phalanx growth forms in Leymus secalinus under different nutrient supplies. Ann. Bot. 2006, 98, 187–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, Y.; He, W.; Pan, X.; Dong, M. Partial mechanical stimulation facilitates the growth of the rhizomatous plant Leymus secalinus: Modulation by clonal integration. Ann. Bot. 2011, 107, 693–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, M. Effects of severing rhizome on clonal growth in rhizomatous grass species Psammochloa villosa and Leymus secalinus. Acta Bot. Sin. 1999, 41, 194–198. [Google Scholar]
- Lv, Z.N. Study on Reproductive Characteristics of Several Representative Rhizomatous Grasses. Master Thesis, Inner Mongolia Agricultural University, Inner Mongolia, China, 2010. [Google Scholar]
- Liu, F.; Andersen, M.N.; Jacobsen, S.E.; Jensen, C.R. Stomatal control and water use efficiency of soybean (Glycine max L. Merr.) during progressive soil drying. Environ. Exp. Bot. 2005, 54, 33–40. [Google Scholar] [CrossRef]
- Medrano, H.; Tomás, M.; Martorell, S.; Flexasa, J.; Hernándeza, E.; Rossellóa, J.; Poub, A.; Escalonaa, J.; Bota, J. From leaf to whole-plant water use efficiency (WUE) in complex canopies: Limitations of leaf WUE as a selection target. Crop J. 2015, 3, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, J.P.; Flury, M.; Huggins, D.R.; Bezdicek, D.F. Soil water and nitrogen dynamics in dryland cropping systems of Washington State, USA. Soil Tillage Res. 2003, 71, 33–47. [Google Scholar] [CrossRef]
- Cheng, L.; Zhang, L.; Wang, Y.P.; Canadell, J.G.; Chiew, F.; Beringer, J.; Li, L.; Miralles, D.G.; Piao, S.; Zhang, Y. Recent increases in terrestrial carbon uptake at little cost to the water cycle. Nat. Commun. 2017, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Ortuño, M.; Serra-Mora, P.; Herráez-Hernández, R.; Verdú-Andrés, J.; Campíns-Falcó, P. A new tool for direct non-invasive evaluation of chlorophyll a content from diffuse reflectance measurements. Sci. Total Environ. 2017, 609, 370–376. [Google Scholar] [CrossRef]
- Mutanga, O.; Skidmore, A.K. Red edge shift and biochemical content in grass canopies. ISPRS J. Photogramm. Remote Sens. 2007, 62, 34–42. [Google Scholar] [CrossRef]
- Mutanga, O.; Skidmore, A.K. Discriminating sodium concentration in a mixed grass species environment of the Kruger National Park using field spectrometry. Int. J. Remote Sens. 2004, 25, 4191–4201. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, W.; Zhou, Q. Reflectance variation within the in-chlorophyll centre waveband for robust retrieval of leaf chlorophyll content. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Du, E.; Dong, D.; Zeng, X.; Sun, Z.; Jiang, X.; Vries, W.D. Direct effect of acid rain on leaf chlorophyll content of terrestrial plants in China. Sci. Total Environ. 2017, 605, 764–769. [Google Scholar] [CrossRef]
- Akram, N.A.; Ashraf, M. Improvement in growth, chlorophyll pigments and photosynthetic performance in salt-stressed plants of sunflower (Helianthus annuus L.) by foliar application of 5-aminolevulinic acid. Agrochimica 2011, 55, 94–104. [Google Scholar]
- Kannan, N.D.; Kulandaivelu, G. Drought induced changes in physiological, biochemical and phytochemical properties of Withania somnifera Dun. J. Med. Plant Res. 2011, 5, 3929–3935. [Google Scholar] [CrossRef]
- Ling, B.; Goodin, D.G.; Raynor, E.J.; Joern, A. Hyperspectral Analysis of Leaf Pigments and Nutritional Elements in Tallgrass Prairie Vegetation. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Esteban, R.; Barrutia, O.; Artetxe, U.; Fernández-Marín, B.; Hernández, A.; García-Plazaola, J.I. Internal and external factors affecting photosynthetic pigment composition in plants: A meta-analytical approach. New Phytol. 2015, 206, 268–280. [Google Scholar] [CrossRef]
- Neilsen, J.A.D.; Rangsrikitphoti, P.; Durnford, D.G. Evolution and regulation of Bigelowiella natans light-harvesting antenna system. J. Plant Physiol. 2017, 217, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Qi, S.; Cao, F.; Zhang, J.; Zhao, F.; Li, C.; Wang, C. Toxic effects of boscalid on the growth, photosynthesis, antioxidant system and metabolism of Chlorella vulgaris. Environ. Pollut. 2018, 242, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Ballottari, M.; Dall’Osto, L.; Morosinotto, T.; Bassi, R. Contrasting behavior of higher plant photosystem I and II antenna systems during acclimation. J. Biol. Chem. 2007, 282, 8947–8958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, J.M.; Chow, W.S.; Rivas, J.D.L. Dynamic flexibility in the structure and function of photosystem II in higher plant thylakoid membranes: The grana enigma. Photosynth. Res. 2008, 98, 575–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinç, E.; Ceppi, M.G.; Tóth, S.Z.; Bottka, S.; Schansker, G. The chl a fluorescence intensity is remarkably insensitive to changes in the chlorophyll content of the leaf as long as the chl a/b ratio remains unaffected. BBA Bioenerg. 2012, 1817, 770–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballottari, M.; Alcocer, M.J.P.; D’Andrea, C.; Viola, D.; Bassi, R. Regulation of photosystem I light harvesting by zeaxanthin. Proc. Natl. Acad. Sci. USA 2014, 111, E2431–E2438. [Google Scholar] [CrossRef] [Green Version]
- Terashima, I.; Inoue, Y. Vertical gradient in photosynthetic properties of spinach chloroplast dependent on intra-leaf light environment. Plant Cell Physiol. 1985, 26, 781–785. [Google Scholar] [CrossRef]
- Davison, P.A.; Hunter, C.N.; Horton, P. Overexpression of β-carotene hydroxylase enhances stress tolerance in Arabidopsis. Nature 2002, 418, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Hendry, G.A.F.; Price, A.H. Stress indicators: Chlorophylls and carotenoids. In Methods in Comparative Plant Ecology; Hendry, G.A.F., Grime, J.P., Eds.; Chapman & Hall: London, UK, 1993; pp. 148–152. [Google Scholar]
- Sonobe, R.; Miura, Y.; Sano, T.; Horie, H. Estimating leaf carotenoid contents of shade-grown tea using hyperspectral indices and PROSPECT–D inversion. Int. J. Remote Sens. 2018, 39, 1306–1320. [Google Scholar] [CrossRef]
- Oliveira, M.T.; Matzek, V.; Medeiros, C.D.; Rivas, R.; Falcao, H.M.; Santos, M.G. Stress tolerance and ecophysiological ability of an invader and a native species in a seasonally dry tropical forest. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Molina-Montenegro, M.A.; Penuelas, J.; Munné-Bosch, S.; Sardans, J. Higher plasticity in ecophysiological traits enhances the performance and invasion success of Taraxacum officinale (dandelion) in alpine environments. Biol. Invasions 2012, 14, 21–33. [Google Scholar] [CrossRef]
- Savchenko, T.; Yanykin, D.; Khorobrykh, A.; Terentyev, V.; Klimov, V.; Dehesh, K. The hydroperoxide lyase branch of the oxylipin pathway protects against photoinhibition of photosynthesis. Planta 2017, 245, 1179–1192. [Google Scholar] [CrossRef]
- Ranjbarfordoei, A.; Samson, R.; Damme, V.P. Photosynthesis performance in sweet almond [Prunus dulcis (Mill) D. Webb] exposed to supplemental UV-B radiation. Photosynthetica 2011, 49, 107. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Xu, H.; Zhang, J.; Liang, G.; Liu, Y.; Guo, A. Effect of low temperature on chlorophyll biosynthesis in albinism line of wheat (Triticum aestivum) FA85. Physiol. Plant. 2012, 145, 384–394. [Google Scholar] [CrossRef] [PubMed]
- Strid, Å.; Porra, R.J. Alterations in pigment content in leaves of Pisum sativum after exposure to supplementary UV-B. Plant Cell Physiol. 1992, 33, 1015–1023. [Google Scholar] [CrossRef]
- Habibi, F.; Normahamadi, G.H.; Heidary, S.A.H.; Eivazi, A.; Majidi, H. Effect of cold stress on cell membrane stability, chlorophyll a and b contain and proline accumulation in wheat (Triticum aiestivum L.) variety. Afr. J. Agric. Res. 2011, 6, 5854–5859. [Google Scholar] [CrossRef]
- Essemine, J.; Xiao, Y.; Qu, M.; Mi, H.; Zhu, X. Cyclic electron flow may provide some protection against PSII photoinhibition in rice (Oryza sativa L.) leaves under heat stress. J. Plant Physiol. 2017, 211, 138–146. [Google Scholar] [CrossRef]
- Eric, L.; William, O.; Stephen, M.; Richard, M. Isolation and characterization of a light-harvesting chlorophyll a/b protein complex associated with photosystem I. Plant Physiol. 1984, 74, 650–655. [Google Scholar]
- Niinemets, U. Photosynthesis and resource distribution through plant canopies. Plant Cell Environ. 2007, 30, 1052–1071. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, Ü. A review of light interception in plant stands from leaf to canopy in different plant functional types and in species with varying shade tolerance. Ecol. Res. 2010, 25, 693–714. [Google Scholar] [CrossRef]
- Mhatre, A.; Patil, S.; Agarwal, A.; Pandit, R.; Lali, A. Influence of nitrogen source on photochemistry and antenna size of the photosystems in marine green macroalgae, Ulva lactuca. Photosynth. Res. 2019, 139, 539–551. [Google Scholar] [CrossRef] [PubMed]
- Derks, A.; Schaven, K.; Bruce, D. Diverse mechanisms for photoprotection in photosynthesis. Dynamic regulation of photosystem II excitation in response to rapid environmental change. BBA Bioenerg. 2015, 1847, 468–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, N.E.; Zigmantas, D.; Valkunas, L.; Li, X.; Niyogi, K.K.; Fleming, G.R. Carotenoid cation formation and the regulation of photosynthetic light harvesting. Science 2005, 307, 433–436. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Fischer, A.L.; Steen, C.J.; Iwai, M.; Morris, J.M.; Walla, P.J.; Niyogi, K.K.; Fleming, G.R. Chlorophyll-carotenoid excitation energy transfer in high-light-exposed thylakoid membranes investigated by snapshot transient absorption spectroscopy. J. Am. Chem. Soc. 2018, 140, 11965–11973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demmig-Adams, B. Survey of thermal energy dissipation and pigment composition in sun and shade leaves. Plant Cell Physiol. 1998, 39, 474–482. [Google Scholar] [CrossRef]
- Zhang, X.; Wollenweber, B.; Jiang, D.; Liu, F.; Zhao, J. Water deficits and heat shock effects on photosynthesis of a transgenic Arabidopsis thaliana constitutively expressing ABP9, a bZIP transcription factor. J. Exp. Bot. 2008, 59, 839–848. [Google Scholar] [CrossRef] [Green Version]
- González, J.A.; Gallardo, M.G.; Boero, C.; Cruz, M.L.; Prado, F.E. Altitudinal and seasonal variation of protective and photosynthetic pigments in leaves of the world’s highest elevation trees Polylepis tarapacana (Rosaceae). Acta Oecol. 2007, 32, 36–41. [Google Scholar] [CrossRef]
- Raynor, E.J.; Derner, J.D.; Baldwin, T.; Ritten, J.P.; Augustine, D.J. Multidecadal directional shift in shortgrass stocking rates. Rangel. Ecol. Manag. 2020, 74, 72–80. [Google Scholar] [CrossRef]
- Hou, F.; Nan, Z.; Xie, Y.; Li, X.; Lin, H.; Ren, J. Integrated crop-livestock production systems in China. Rangel. J. 2008, 30, 221–231. [Google Scholar] [CrossRef]
- Wu, Z.; Hou, X.; Zhu, Y.; He, S.; Zhou, Y. Vegetation of China; Science Press: Beijing, China, 1980. [Google Scholar]
- Zhang, Y.; Chen, X.; Cheng, Y.; Chang, S.; Hou, F. Effects of stocking rates on functional group diversity and forage quality in rangeland of Qilian Mountain, China. J. Environ. Biol. 2015, 36, 713–719. [Google Scholar]
- Stępień, P.; Kłobus, G. Water relations and photosynthesis in Cucumis sativus L. leaves under salt stress. Biol. Plant. 2006, 50, 610. [Google Scholar] [CrossRef]
- Inskeep, W.P.; Bloom, P.R. Extinction coefficients of chlorophyll a and b in N, N-dimethylformamide and 80% acetone. Plant Physiol. 1985, 77, 483–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | GI | Chl a | Chl b | TC | Car | Chl a/b | TC/Car |
|---|---|---|---|---|---|---|---|
| (AUM ha−1) | (mg g−1 FW) | (mg g−1 FW) | (mg g−1 FW) | (mg g−1 FW) | |||
| S. purpurea | LG | 1.69 ± 0.09 c | 0.67 ± 0.02 b | 2.36 ± 0.08 c | 0.42 ± 0.04 b | 2.54 ± 0.19 abc | 5.60 ± 0.28 c |
| MG | 1.72 ± 0.07 c | 0.70 ± 0.06 b | 2.41 ± 0.11 c | 0.34 ± 0.03 bc | 2.50 ± 0.20 abc | 7.24 ± 0.51 c | |
| HG | 2.05 ± 0.07 b | 0.96 ± 0.10 a | 3.01 ± 0.15 b | 0.29 ± 0.03 c | 2.17 ± 0.20 bc | 10.40 ± 0.44 b | |
| A. inebrians | LG | 2.68 ± 0.14 a | 0.96 ± 0.02 a | 3.64 ± 0.15 a | 0.58 ± 0.03 a | 2.80 ± 0.12 abc | 6.31 ± 0.25 c |
| MG | 2.43 ± 0.02 a | 0.95 ± 0.01 a | 3.38 ± 0.03 a | 0.56 ± 0.01 a | 2.56 ± 0.02 abc | 6.04 ± 0.07 c | |
| HG | 1.51 ± 0.08 c | 0.77 ± 0.02 ab | 2.29 ± 0.09 c | 0.42 ± 0.01 b | 1.96 ± 0.09 c | 5.43 ± 0.09 c | |
| L. secalinus | LG | 0.65 ± 0.02 d | 0.21 ± 0.02 c | 0.85 ± 0.02 de | 0.08 ± 0.00 d | 3.22 ± 0.32 a | 11.25 ± 0.32 b |
| MG | 0.45 ± 0.02 d | 0.14 ± 0.01 c | 0.59 ± 0.01 e | 0.06 ± 0.00 d | 3.15 ± 0.33 ab | 9.74 ± 0.19 b | |
| HG | 0.68 ± 0.00 d | 0.32 ± 0.03 c | 1.00 ± 0.04 d | 0.05 ± 0.00 d | 2.19 ± 0.22 b c | 19.01 ± 1.18 a | |
| Mean | S. purpurea | 1.82 ± 0.07 B | 0.78 ± 0.06 B | 2.59 ± 0.12 B | 0.35 ± 0.03 B | 2.40 ± 0.11 B | 7.75 ± 0.73 B |
| A. inebrians | 2.21 ± 0.18 A | 0.89 ± 0.03 A | 3.10 ± 0.21 A | 0.52 ± 0.03 A | 2.44 ± 0.13 B | 5.93 ± 0.15 C | |
| L. secalinus | 0.59 ± 0.04 C | 0.22 ± 0.03 C | 0.82 ± 0.06 C | 0.06 ± 0.00 C | 2.85 ± 0.22 A | 13.33 ± 1.48 A | |
| LG | 1.67 ± 0.30 A | 0.61 ± 0.11 A | 2.28 ± 0.41 A | 0.36 ± 0.08 A | 2.85 ± 0.15 A | 7.72 ± 0.90 B | |
| MG | 1.53 ± 0.29 B | 0.60 ± 0.12 A | 2.13 ± 0.41 A | 0.32 ± 0.07 B | 2.74 ± 0.15 A | 7.67 ± 0.57 B | |
| HG | 1.41 ± 0.20 B | 0.68 ± 0.10 A | 2.10 ± 0.30 A | 0.26 ± 0.05 C | 2.10 ± 0.10 B | 11.61 ± 2.02 A | |
| Significance level | Species | *** | *** | *** | *** | * | *** |
| GI | *** | * | * | *** | *** | *** | |
| Species × GI | *** | *** | *** | * | NS | *** |
| Species | GI | Chl a | Chl b | TC | Car | Chl a/b | TC/Car |
|---|---|---|---|---|---|---|---|
| (AUM ha−1) | (mg g−1 FW) | (mg g−1 FW) | (mg g−1 FW) | (mg g−1 FW) | |||
| S. purpurea | LG | 0.55 ± 0.02 a | 0.32 ± 0.03 a | 0.87 ± 0.05 a | 0.073 ± 0.007 b c | 1.74 ± 0.13 ef | 12.05 ± 0.92 ab |
| MG | 0.45 ± 0.01 ab | 0.12 ± 0.00 cde | 0.57 ± 0.01 bc | 0.093 ± 0.009 b | 3.65 ± 0.10 ab | 6.22 ± 0.54 c d | |
| HG | 0.48 ± 0.02 a | 0.17 ± 0.01 bc | 0.65 ± 0.02 b | 0.143 ± 0.008 a | 2.82 ± 0.14 cd | 4.54 ± 0.15 d | |
| A. inebrians | LG | 0.35 ± 0.02 bc | 0.32 ± 0.02 a | 0.67 ± 0.04 b | 0.090 ± 0.005 b | 1.08 ± 0.08 f | 7.47 ± 0.21 c |
| MG | 0.45 ± 0.01 ab | 0.21 ± 0.01 b | 0.65 ± 0.02 b | 0.061 ± 0.003 cd | 2.17 ± 0.12 de | 10.73 ± 0.43 b | |
| HG | 0.37 ± 0.04 b | 0.09 ± 0.01 de | 0.46 ± 0.04 cd | 0.033 ± 0.002 e | 3.92 ± 0.08 a | 13.91 ± 1.09 a | |
| L. secalinus | LG | 0.35 ± 0.04 bc | 0.15 ± 0.01 bcd | 0.50 ± 0.05 cd | 0.048 ± 0.005 de | 2.28 ± 0.24 de | 10.42 ± 0.75 b |
| MG | 0.20 ± 0.02 d | 0.07 ± 0.00 e | 0.27 ± 0.02 e | 0.057 ± 0.001 cd | 3.10 ± 0.33 bc | 4.77 ± 0.41 cd | |
| HG | 0.26 ± 0.02 cd | 0.10 ± 0.00 de | 0.36 ± 0.02 de | 0.061 ± 0.003 cd | 2.64 ± 0.17 cd | 5.84 ± 0.06 cd | |
| Mean | S. purpurea | 0.49 ± 0.02 A | 0.20 ± 0.03 A | 0.70 ± 0.05 A | 0.103 ± 0.011 A | 2.74 ± 0.28 A | 7.60 ± 1.18 B |
| A. inebrians | 0.39 ± 0.02 B | 0.21 ± 0.03 A | 0.59 ± 0.04 B | 0.061 ± 0.008 B | 2.39 ± 0.42 A | 10.70 ± 0.99 A | |
| L. secalinus | 0.27 ± 0.03 C | 0.11 ± 0.01 B | 0.37 ± 0.04 C | 0.055 ± 0.003 B | 2.67 ± 0.17 A | 7.01 ± 0.90 B | |
| LG | 0.41 ± 0.04 A | 0.27 ± 0.03 A | 0.68 ± 0.06 A | 0.070 ± 0.007 A | 1.70 ± 0.19 B | 9.98 ± 0.75 A | |
| MG | 0.37 ± 0.04 B | 0.13 ± 0.02 B | 0.50 ± 0.06 B | 0.070 ± 0.006 A | 2.97 ± 0.24 A | 7.24 ± 0.93 B | |
| HG | 0.37 ± 0.03 B | 0.12 ± 0.01 B | 0.49 ± 0.05 B | 0.079 ± 0.017 A | 3.13 ± 0.21 A | 8.10 ± 1.50 B | |
| Significance level | Species | *** | *** | *** | *** | NS | *** |
| GI | * | *** | *** | NS | *** | *** | |
| Species × GI | *** | *** | *** | *** | *** | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Hou, F.; Ren, J. Grazing Intensity Alters Leaf and Spike Photosynthesis, Transpiration, and Related Parameters of Three Grass Species on an Alpine Steppe in the Qilian Mountains. Plants 2021, 10, 294. https://doi.org/10.3390/plants10020294
Li J, Hou F, Ren J. Grazing Intensity Alters Leaf and Spike Photosynthesis, Transpiration, and Related Parameters of Three Grass Species on an Alpine Steppe in the Qilian Mountains. Plants. 2021; 10(2):294. https://doi.org/10.3390/plants10020294
Chicago/Turabian StyleLi, Jin, Fujiang Hou, and Jizhou Ren. 2021. "Grazing Intensity Alters Leaf and Spike Photosynthesis, Transpiration, and Related Parameters of Three Grass Species on an Alpine Steppe in the Qilian Mountains" Plants 10, no. 2: 294. https://doi.org/10.3390/plants10020294