
Assessment of the Spatial Invasion Risk of Intentionally Introduced Alien Plant Species (IIAPS) under Environmental Change in South Korea



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Areas
2.2. Occurrence of Intentionally Introduced Alien Plant Species
2.3. Environmental Variables
2.4. Species Distribution Modeling
2.5. Model Evaluation and Validation
2.6. Prediction of the Spatial Distribution of IIAPS
2.7. Prediction of the Spatial Invasion Risk of IIAPS
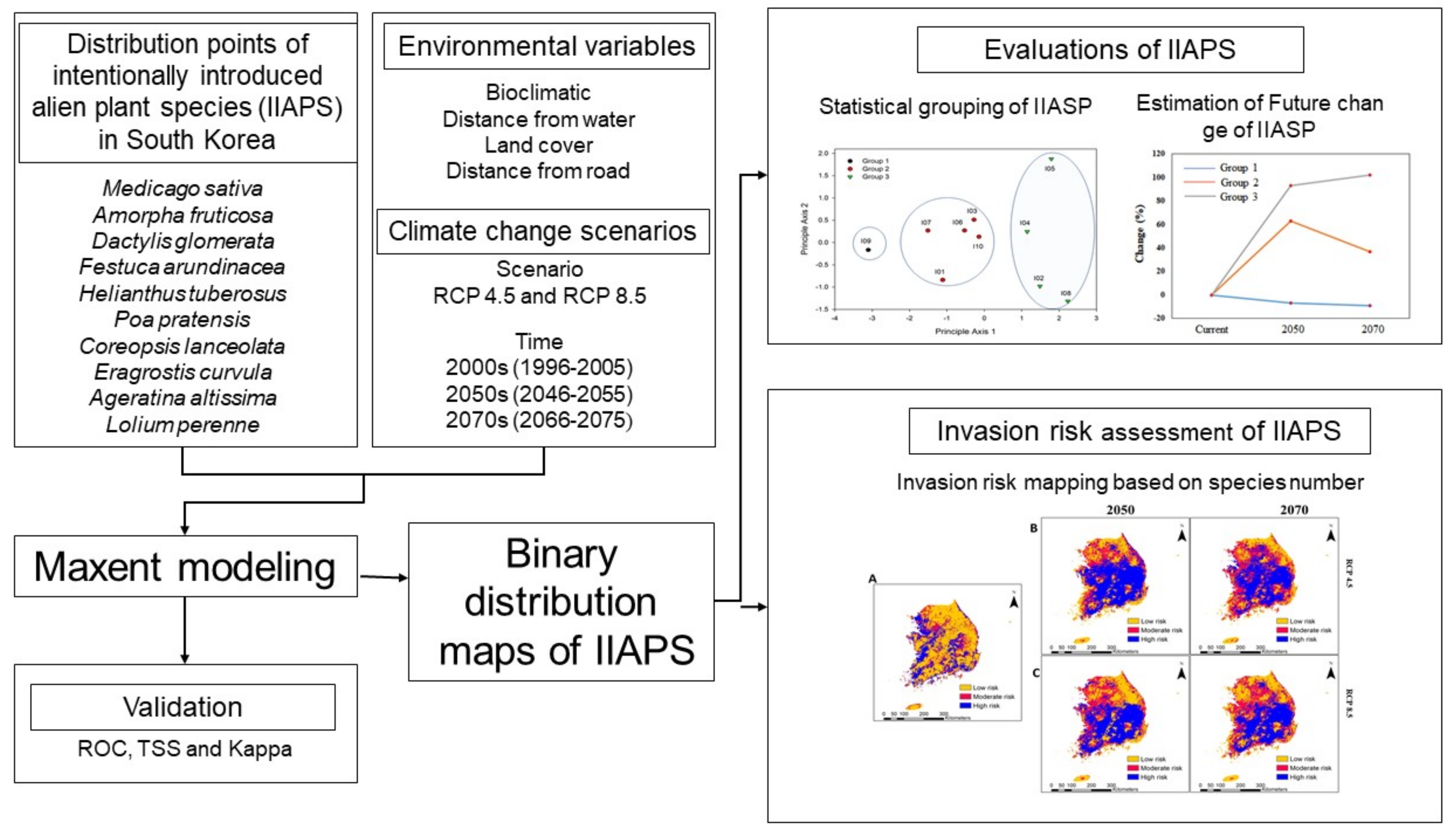
3. Results
3.1. Selection and Evaluation of Variables
3.2. AUC, TSS, and Kappa Values Show Excellent Model Prediction for All IIAPS
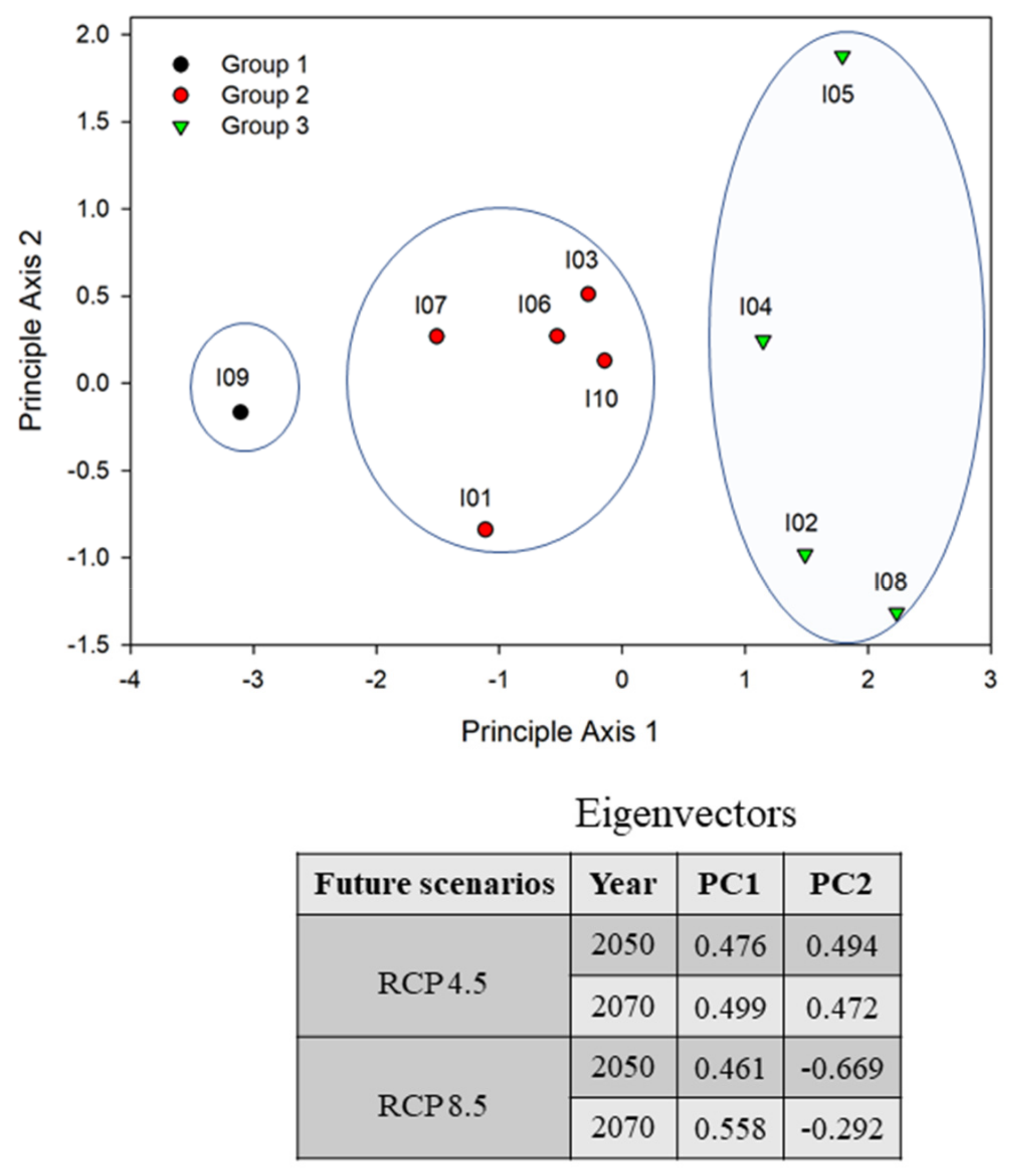
3.3. Environmental Changes Positively Regulate the Spatial Distribution of IIAPS in South Korea
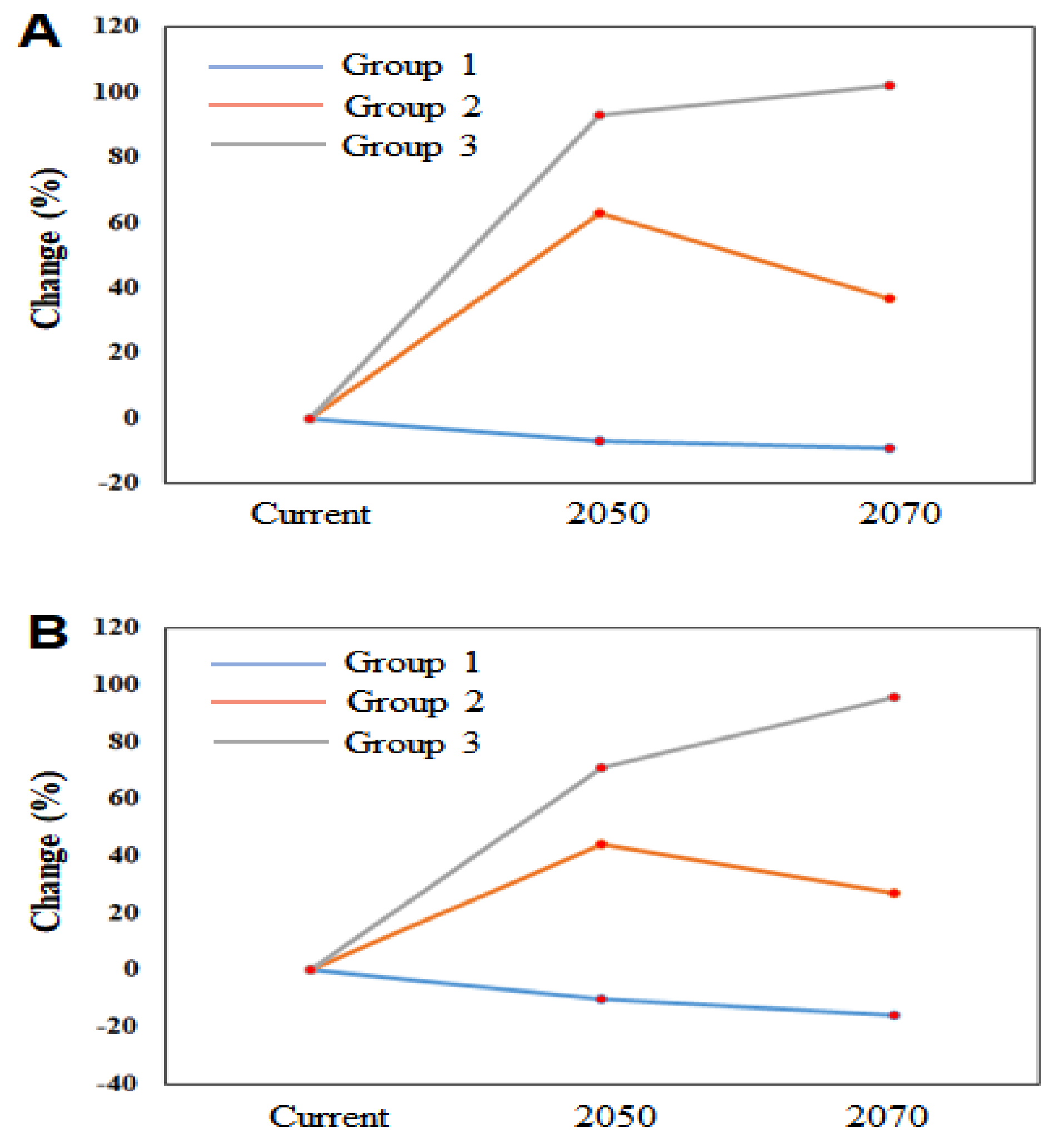
3.4. Environmental Changes Increase the Spatial Invasion Risk of IIAPS in South Korea
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ansong, M.; Pickering, C. What’sa weed? Knowledge, attitude and behaviour of park visitors about weeds. PLoS ONE 2015, 10, e0135026. [Google Scholar] [CrossRef] [Green Version]
- Parker, C. Parasitic weeds: A world challenge. Weed Sci. 2012, 60, 269–276. [Google Scholar] [CrossRef]
- IUCN. Invasive Alien Species and Cliamte Change. Available online: http://iucn.org (accessed on 28 August 2021).
- Cook, G.D.; Dias, L. It was no accident: Deliberate plant introductions by Australian government agencies during the 20th century. Aust. J. Bot. 2006, 54, 601–625. [Google Scholar] [CrossRef]
- Van Klinken, R.D.; Friedel, M.H. Unassisted invasions: Understanding and responding to Australia’s high-impact environmental grass weeds. Aust. J. Bot. 2018, 65, 678–690. [Google Scholar] [CrossRef] [Green Version]
- Keller, R.P.; Geist, J.; Jeschke, J.M.; Kühn, I. Invasive species in Europe: Ecology, status, and policy. Environ. Sci. Eur. 2011, 23, 23. [Google Scholar] [CrossRef] [Green Version]
- Divíšek, J.; Chytrý, M.; Beckage, B.; Gotelli, N.J.; Lososová, Z.; Pyšek, P.; Richardson, D.M.; Molofsky, J. Similarity of introduced plant species to native ones facilitates naturalization, but differences enhance invasion success. Nat. Commun. 2018, 9, 4631. [Google Scholar] [CrossRef]
- Ordonez, A.; Wright, I.J.; Olff, H. Functional differences between native and alien species: A global-scale comparison. Funct. Ecol. 2010, 24, 1353–1361. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P. Naturalization of introduced plants: Ecological drivers of biogeographical patterns. New Phytol. 2012, 196, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Weber, E. Invasive Plant Species of the World: A Reference Guide to Environmental Weeds; CABI: Potsdam, Germany, 2017. [Google Scholar]
- Fletcher, D.; Gillingham, P.; Britton, J.; Blanchet, S.; Gozlan, R.E. Predicting global invasion risks: A management tool to prevent future introductions. Sci. Rep. 2016, 6, 26316. [Google Scholar] [CrossRef] [Green Version]
- Fahad, S.; Hussain, S.; Chauhan, B.S.; Saud, S.; Wu, C.; Hassan, S.; Tanveer, M.; Jan, A.; Huang, J. Weed growth and crop yield loss in wheat as influenced by row spacing and weed emergence times. Crop Protect. 2015, 71, 101–108. [Google Scholar] [CrossRef]
- Bradley, B.A.; Wilcove, D.S.; Oppenheimer, M. Climate change increases risk of plant invasion in the Eastern United States. Biol. Invasions 2010, 12, 1855–1872. [Google Scholar] [CrossRef]
- Paini, D.R.; Sheppard, A.W.; Cook, D.C.; De Barro, P.J.; Worner, S.P.; Thomas, M.B. Global threat to agriculture from invasive species. Proc. Natl. Acad. Sci. USA 2016, 113, 7575–7579. [Google Scholar] [CrossRef] [Green Version]
- Walsh, J.R.; Carpenter, S.R.; Vander Zanden, M.J. Invasive species triggers a massive loss of ecosystem services through a trophic cascade. Proc. Natl. Acad. Sci. USA 2016, 113, 4081–4085. [Google Scholar] [CrossRef] [Green Version]
- Thuiller, W.; Richardson, D.M.; Midgley, G.F. Will climate change promote alien plant invasions. In Biological Invasions; Springer: Berlin/Heidelberg, Germany, 2008; pp. 197–211. [Google Scholar]
- Weltzin, J.F.; Belote, R.T.; Sanders, N.J. Biological invaders in a greenhouse world: Will elevated CO2 fuel plant invasions? Front. Ecol. Environ. 2003, 1, 146–153. [Google Scholar] [CrossRef]
- Debouk, H.; de Bello, F.; Sebastià, M.-T. Functional trait changes, productivity shifts and vegetation stability in mountain grasslands during a short-term warming. PLoS ONE 2015, 10, e0141899. [Google Scholar]
- Rasmussen, K.; Thyrring, J.; Muscarella, R.; Borchsenius, F. Climate-change-induced range shifts of three allergenic ragweeds (Ambrosia L.) in Europe and their potential impact on human health. PeerJ 2017, 5, e3104. [Google Scholar] [CrossRef] [Green Version]
- Hellmann, J.J.; Byers, J.E.; Bierwagen, B.G.; Dukes, J.S. Five potential consequences of climate change for invasive species. Conserv. Biol. 2008, 22, 534–543. [Google Scholar] [CrossRef]
- Ahmad, R.; Khuroo, A.A.; Charles, B.; Hamid, M.; Rashid, I.; Aravind, N. Global distribution modelling, invasion risk assessment and niche dynamics of Leucanthemum vulgare (Ox-eye Daisy) under climate change. Sci. Rep. 2019, 9, 11395. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2013; p. 1535. [Google Scholar]
- Manzoor, S.A.; Griffiths, G.; Lukac, M. Land use and climate change interaction triggers contrasting trajectories of biological invasion. Ecol. Indic. 2021, 120, 106936. [Google Scholar] [CrossRef]
- Vila, M.; Ibáñez, I. Plant invasions in the landscape. Landsc. Ecol. 2011, 26, 461–472. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, C.; Allen, J.M.; Li, W.; Boyer, M.A.; Segerson, K.; Silander, J.A. Analysis and prediction of land use changes related to invasive species and major driving forces in the state of Connecticut. Land 2016, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Mosher, E.S.; Silander, J.A.; Latimer, A.M. The role of land-use history in major invasions by woody plant species in the northeastern North American landscape. Biol. Invasions 2009, 11, 2317–2328. [Google Scholar] [CrossRef] [Green Version]
- Young, J.S.; Shin, H.T.; Kim, S.J.; An, J.B.; Heo, T.I.; Chung, J.M.; Cho, Y.C. Invasive Alien Plants in South Korea; Korea National Arboretum: Pocheon, Korea, 2017. [Google Scholar]
- NIE. Monitoring of Invasive Alien Species Designated by the Act on the Conservation and Use of Biological Diversity (IV); National Institute of Ecology: Seocheon-gun, Korea, 2017. [Google Scholar]
- Lee, I.-Y.; Oh, Y.-J.; Hong, S.-H.; Heo, S.-J.; Lee, C.-Y.; Park, K.W.; Cho, S.-H.; Kwon, O.-D.; Im, I.-B.; Kim, S.-K. Occurrence of weed flora and changes in weed vegetation in orchard fields of Korea. Weed Turfgrass Sci. 2017, 6, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Jongchul Park, J.K. Land Use and land Cover. 2021. Available online: http://nationalatlas.ngii.go.kr/pages/page_756.php (accessed on 11 November 2021).
- Choung, Y.-J. Land cover change detection in the Nakdong river basin using LiDAR data and multi-temporal landsat imagery. J. Korean Assoc. Geogr. Inf. Stud. 2015, 18, 135–148. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Kang, E.S.; Lee, S.-R.; Oh, S.H.; Kim, D.-K.; Jung, S.-Y.; Son, D.C. Comprehensive review about alien plants in Korea. Korean J. Plant Taxon. 2020, 50, 89–119. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A.; Peterson, A.T.; Soberón, J.; Overton, J.M.; Aragón, P.; Lobo, J.M. Use of niche models in invasive species risk assessments. Biol. Invasions 2011, 13, 2785–2797. [Google Scholar] [CrossRef]
- NIBR. Korean Red List of Threatened Species, 2nd ed.; Min-Hwan Suh, B.-Y.L., Kim, S.T., Park, C.-H., Oh, H.-K., Kim, H.-Y., Lee, J.-H., Lee, S.-Y., Eds.; National Institute of Biological Resources, Ministry of Environment: Incheon, Korea, 2014; p. 242.
- Koo, K.A.; Kong, W.S.; Nibbelink, N.P.; Hopkinson, C.S.; Lee, J.H. Potential effects of climate change on the distribution of cold-tolerant evergreen broadleaved woody plants in the Korean Peninsula. PLoS ONE 2015, 10, e0134043. [Google Scholar] [CrossRef]
- KMA. Climate of Korea; KMA: Seoul, Korea, 2021; Available online: http://web.kma.go.kr (accessed on 27 August 2021).
- NIER. The Second and Third National Ecosystem Survey: 1997–2012; National Institute of Environmental Research: Incheon, Korea, 2013.
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef] [Green Version]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- Busby, J. Bioclim, a bioclimatic analysis and prediction system. In Nature Conservation: Cost Effective Biological Surveys and Data Analysis; Margules, C., Austin, M., Eds.; CSIRO: Canberra, Australia, 1991; pp. 64–68. [Google Scholar]
- Amiri, M.; Tarkesh, M.; Jafari, R.; Jetschke, G. Bioclimatic variables from precipitation and temperature records vs. remote sensing-based bioclimatic variables: Which side can perform better in species distribution modeling? Ecol. Inform. 2020, 57, 101060. [Google Scholar] [CrossRef]
- McGuffie, K.; Henderson-Sellers, A. The Climate Modelling Primer; John Wiley & Sons: Hoboken, NJ, USA, 2014. [Google Scholar]
- Lee, I.H.; Park, S.H.; Kang, H.S.; Cho, C.H. Regional climate projections using the HadGEM3-RA. In Proceedings of the 3rd International Conference on Earth System Modelling, Hamburg, Germany, 17–21 September 2012. [Google Scholar]
- NIMS. National Institute of Meterological Sciences HadGEM3-RA. Available online: http://cordex-ea.climate.go.kr/cordex/models.do (accessed on 19 July 2021).
- Robert, J.; Hijmans, S.P.; Leathwick, J.; Elith, J. Species Distribution Modeling Package ‘Dismo’. 2020. Available online: https://cran.r-project.org/web/packages/dismo/dismo.pdf (accessed on 11 October 2021).
- Adhikari, P.; Shin, M.-S.; Jeon, J.-Y.; Kim, H.W.; Hong, S.; Seo, C. Potential impact of climate change on the species richness of subalpine plant species in the mountain national parks of South Korea. J. Ecol. Environ. 2018, 42, 36. [Google Scholar] [CrossRef]
- Adhikari, P.; Jeon, J.-Y.; Kim, H.W.; Shin, M.-S.; Adhikari, P.; Seo, C. Potential impact of climate change on plant invasion in the Republic of Korea. J. Ecol. Environ. 2019, 43, 36. [Google Scholar] [CrossRef]
- Shin, M.-S.; Seo, C.; Lee, M.; Kim, J.-Y.; Jeon, J.-Y.; Adhikari, P.; Hong, S.-B. Prediction of Potential Species Richness of Plants Adaptable to Climate Change in the Korean Peninsula. J. Environ. Impact Assess. 2018, 27, 562–581. [Google Scholar] [CrossRef]
- Kim, H.W.; Adhikari, P.; Chang, M.H.; Seo, C. Potential Distribution of Amphibians with Different Habitat Characteristics in Response to Climate Change in South Korea. Animals 2021, 11, 2185. [Google Scholar] [CrossRef]
- Adhikari, P.; Jeon, J.-Y.; Kim, H.W.; Oh, H.-S.; Adhikari, P.; Seo, C. Northward Range Expansion of Southern Butterflies According to Climate Change in South Korea. J. Clim. Chang. Res. 2020, 11, 643–656. [Google Scholar] [CrossRef]
- Jeon, J.-Y.; Adhikari, P.; Seo, C. Impact of climate change on potential dispersal of Paeonia obovata (paeoniaceae), a critically endangered medicinal plant of South Korea. Ecol. Environ. Conserv. 2020, 26, S145–S155. [Google Scholar]
- McDougall, K.L.; Lembrechts, J.; Rew, L.J.; Haider, S.; Cavieres, L.A.; Kueffer, C.; Milbau, A.; Naylor, B.J.; Nuñez, M.A.; Pauchard, A. Running off the road: Roadside non-native plants invading mountain vegetation. Biol. Invasions 2018, 20, 3461–3473. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.H.; Lee, Y.H.; Lee, G.; Lee, D.-H.; Adhikari, P. Predicting Impacts of Climate Change on Northward Range Expansion of Invasive Weeds in South Korea. Plants 2021, 10, 1604. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Wiens, J.A.; Stralberg, D.; Jongsomjit, D.; Howell, C.A.; Snyder, M.A. Niches, models, and climate change: Assessing the assumptions and uncertainties. Proc. Natl. Acad. Sci. USA 2009, 106, 19729–19736. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, U.B.; Shrestha, B.B. Climate change amplifies plant invasion hotspots in Nepal. Divers. Distrib. 2019, 25, 1599–1612. [Google Scholar] [CrossRef] [Green Version]
- Fournier, A.; Penone, C.; Pennino, M.G.; Courchamp, F. Predicting future invaders and future invasions. Proc. Natl. Acad. Sci. USA 2019, 116, 7905–7910. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudik, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Thuiller, W.G.D.; Gueguen, M.; Engler, R.; Breiner, F. Package ‘biomod2’: Ensemble Platform for Species Distribution Modeling, Version 3.5.1. 2021. Available online: https://cran.r-project.org/web/packages/biomod2/index.html (accessed on 11 October 2021).
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Pearson, R.G. Species’ distribution modeling for conservation educators and practitioners. Lessons Conserv. 2010, 3, 54–89. [Google Scholar]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Araújo, M.B. Niche properties and geographical extent as predictors of species sensitivity to climate change. Glob. Ecol. Biogeogr. 2005, 14, 347–357. [Google Scholar] [CrossRef]
- Lobo, J.M.; Jiménez-Valverde, A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Farzin Shabani, L.K.; Ahmadi, M. Assessing accuracy methods of species distribution models: AUC, specificity, sensitivity and the true skill statistic. Glob. J. Hum. Soc. Sci. Res. 2018, 18, 7–17. [Google Scholar]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How much does climate change threaten European forest tree species distributions? Glob. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Phillips, S.J. A Brief Tutorial on Maxent 2017. Available online: https://biodiversityinformatics.amnh.org/open_source/maxent/Maxent (accessed on 11 October 2021).
- Baldwin, R.A. Use of maximum entropy modeling in wildlife research. Entropy 2009, 11, 854–866. [Google Scholar] [CrossRef]
- Hulme, P.E.; Bacher, S.; Kenis, M.; Klotz, S.; Kühn, I.; Minchin, D.; Nentwig, W.; Olenin, S.; Panov, V.; Pergl, J. Grasping at the routes of biological invasions: A framework for integrating pathways into policy. J. Appl. Ecol. 2008, 45, 403–414. [Google Scholar] [CrossRef]
- Adhikari, P.; Park, S.-M.; Kim, T.-W.; Lee, J.-W.; Kim, G.-R.; Han, S.-H.; Oh, H.-S. Seasonal and altitudinal variation in roe deer (Capreolus pygargus tianschanicus) diet on Jeju Island, South Korea. J. Asia-Pac. Biodivers. 2016, 9, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Elith, J. On estimating probability of presence from use–availability or presence–background data. Ecology 2013, 94, 1409–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarnevich, C.S.; Stohlgren, T.J.; Barnett, D.; Kartesz, J. Filling in the gaps: Modelling native species richness and invasions using spatially incomplete data. Divers. Distrib. 2006, 12, 511–520. [Google Scholar] [CrossRef]
- West, A.M.; Kumar, S.; Brown, C.S.; Stohlgren, T.J.; Bromberg, J. Field validation of an invasive species Maxent model. Ecol. Inform. 2016, 36, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wang, Y. Applying various algorithms for species distribution modelling. Integr. Zool. 2013, 8, 124–135. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Stockwell, D. The GARP modelling system: Problems and solutions to automated spatial prediction. Int. J. Geogr. Inf. Sci. 1999, 13, 143–158. [Google Scholar] [CrossRef]
- Hyung-Kyu, N.; Song, Y.-J.; Kwon, S.-I.; Eo, J.; Kim, M.-H. Potential changes in the distribution of seven agricultural indicator plant species in response to climate change at Agroecosystem in South Korea. Korean J. Ecol. Environ. 2018, 51, 221–233. [Google Scholar] [CrossRef]
- Chung, M.Y.; Son, S.; Suh, G.U.; Herrando-Moraira, S.; Lee, C.H.; López-Pujol, J.; Chung, M.G. The Korean Baekdudaegan Mountains: A glacial refugium and a biodiversity hotspot that needs to be conserved. Front. Genet. 2018, 9, 489. [Google Scholar] [CrossRef] [Green Version]
- Chung, G.Y.; Chang, K.S.; Chung, J.-M.; Choi, H.J.; Paik, W.-K.; Hyun, J.-O. A checklist of endemic plants on the Korean Peninsula. Korean J. Plant Taxon. 2017, 47, 264–288. [Google Scholar] [CrossRef] [Green Version]
- Dullinger, I.; Wessely, J.; Bossdorf, O.; Dawson, W.; Essl, F.; Gattringer, A.; Klonner, G.; Kreft, H.; Kuttner, M.; Moser, D. Climate change will increase the naturalization risk from garden plants in Europe. Glob. Ecol. Biogeogr. 2017, 26, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Pellegrini, E.; Buccheri, M.; Martini, F.; Boscutti, F. Agricultural land use curbs exotic invasion but sustains native plant diversity at intermediate levels. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Mortensen, D.A.; Rauschert, E.S.; Nord, A.N.; Jones, B.P. Forest roads facilitate the spread of invasive plants. Invasive Plant Sci. Manag. 2009, 2, 191–199. [Google Scholar] [CrossRef]
- KOSIS. Yearly road statistictics. Available online: http://kosis.kr (accessed on 18 July 2021).
- Catriona, E. Rogers, J.P.M. Climate change and ecosystems of the Mid-Atlantic Region. Clim. Res. 2000, 14, 235–244. [Google Scholar]
- Ni, M.; Deane, D.C.; Li, S.; Wu, Y.; Sui, X.; Xu, H.; Chu, C.; He, F.; Fang, S. Invasion success and impacts depend on different characteristics in non-native plants. Divers. Distrib. 2021. [Google Scholar] [CrossRef]
- Leishman, M.R.; Haslehurst, T.; Ares, A.; Baruch, Z. Leaf trait relationships of native and invasive plants: Community-and global-scale comparisons. New Phytol. 2007, 176, 635–643. [Google Scholar] [CrossRef]
- Carboni, M.; Münkemüller, T.; Lavergne, S.; Choler, P.; Borgy, B.; Violle, C.; Essl, F.; Roquet, C.; Munoz, F.; Consortium, D. What it takes to invade grassland ecosystems: Traits, introduction history and filtering processes. Ecol. Lett. 2016, 19, 219–229. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M. Traits associated with invasiveness in alien plants: Where do we stand. In Biol. Invasions; Springer: Berlin/Heidelberg, Germany, 2008; pp. 97–125. [Google Scholar]
- Buri, A.; Cianfrani, C.; Pinto-Figueroa, E.; Yashiro, E.; Spangenberg, J.E.; Adatte, T.; Verrecchia, E.; Guisan, A.; Pradervand, J.-N. Soil factors improve predictions of plant species distribution in a mountain environment. Prog. Phys. Geogr. 2017, 41, 703–722. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M. Invasive species, environmental change and management, and health. Annu. Rev. Environ. Resour. 2010, 35, 25–55. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IAPAS Group a | ID No. | Scientific Name | Common Name | Native Range | Mode of Introduction | Introduction Period | Degree of Naturalization |
|---|---|---|---|---|---|---|---|
| Group 1 | I223 | Medicago sativa | Alfalfa | Mediterranean | Intentional (Pasture) | Before 1949 | III |
| Group 2 | I072 | Amorpha fruticosa | Bastard indigo bush | North America | Intentional (Erosion control) | Before 1949 | V |
| I138 | Dactylis glomerata | Orchard grass | North Africa | Intentional (Pasture) | Before 1949 | V | |
| I165 | Festuca arundinacea | Tall fescue | North Africa | Intentional (Pasture) | Before 1970 | V | |
| I176 | Helianthus tuberosus | Jerusalem artichoke | North America | Intentional (medicinal) | Before 1911 | V | |
| I258 | Poa pratensis | Kentucky bluegrass | Temperate zone | Intentional (Erosion control) | Before 1949 | IV | |
| Group 3 | I129 | Coreopsis lanceolata | Lance leaf coreopsis | North America | Intentional (Erosion control) | Before 1963 | V |
| I150 | Eragrostis curvula | African love grass | North Africa | Intentional (Erosion control) | Before 1990 | IV | |
| I157 | Ageratina altissima | White snakeroot | North America | Intentional (Gardening) | Before 1990 | IV | |
| I210 | Lolium perenne | Ryegrass | North Africa | Intentional (Pasture) | Before 1970 | IV |
| Name of Species | Bio1 | Bio3 | Bio4 | Bio12 | Bio13 | Bio14 | d-Road | d-Water | Land Cover |
|---|---|---|---|---|---|---|---|---|---|
| Amorpha fruticosa | 6.15 a | 10.73 | 6.72 | 8.54 | 0.76 | 11.28 | 4.09 | 9.51 | 42.22 |
| Coreopsis lanceolata | 8.50 | 4.93 | 10.03 | 7.98 | 3.88 | 2.23 | 8.27 | 9.41 | 44.77 |
| Dactylis glomerata | 6.03 | 5.59 | 12.62 | 0.67 | 20.79 | 5.74 | 3.06 | 1.07 | 44.42 |
| Eragrostis curvula | 23.67 | 13.53 | 32.39 | 2.07 | 8.48 | 6.23 | 1.70 | 0.00 | 11.93 |
| Ageratina altissima | 4.18 | 18.88 | 12.72 | 3.37 | 42.98 | 11.10 | 4.94 | 0.42 | 1.42 |
| Festuca arundinacea | 16.33 | 11.51 | 0.88 | 6.44 | 8.33 | 3.77 | 2.33 | 2.01 | 48.40 |
| Helianthus tuberosus | 8.67 | 1.50 | 12.37 | 2.28 | 0.42 | 0.87 | 2.00 | 2.49 | 69.41 |
| Lolium perenne | 11.83 | 1.15 | 7.71 | 37.50 | 3.72 | 2.40 | 3.73 | 0.52 | 31.42 |
| Medicago sativa | 1.15 | 0.68 | 15.34 | 34.30 | 11.17 | 1.66 | 0.16 | 0.17 | 35.37 |
| Poa pratensis | 2.05 | 11.51 | 5.51 | 14.09 | 8.43 | 14.06 | 1.65 | 1.03 | 41.67 |
| Name of Species | No. of Species Presence Points | AUC Value | TSS Value | Kappa Value |
|---|---|---|---|---|
| Amorpha fruticosa | 516 | 0.76 | 0.79 | 0.67 |
| Coreopsis lanceolata | 806 | 0.73 | 0.85 | 0.66 |
| Dactylis glomerata | 634 | 0.72 | 0.81 | 0.57 |
| Eragrostis curvula | 110 | 0.75 | 0.77 | 0.67 |
| Ageratina altissima | 104 | 0.92 | 0.77 | 0.79 |
| Festuca arundinacea | 1076 | 0.73 | 0.72 | 0.64 |
| Helianthus tuberosus | 734 | 0.74 | 0.75 | 0.71 |
| Lolium perenne | 228 | 0.78 | 0.76 | 0.66 |
| Medicago sativa | 242 | 0.76 | 0.74 | 0.68 |
| Species Names | Current (km2) | RCP 4.5 | RCP 8.5 | ||
|---|---|---|---|---|---|
| 2050 (%) | 2070 (%) | 2050 (%) | 2070 (%) | ||
| Medicago sativa | 44,427 | −7 | −9 | −10 | −16 |
| Amorpha fruticosa | 38,060 | 43 | 8 | 63 | 23 |
| Dactylis glomerata | 37,565 | 61 | 91 | 42 | 37 |
| Festuca arundinacea | 32,317 | 83 | 34 | 39 | 45 |
| Helianthus tuberosus | 38,656 | 65 | 14 | 33 | 2 |
| Poa pratensis | 34,272 | 41 | 94 | 42 | 66 |
| Coreopsis lanceolata | 30,027 | 98 | 64 | 101 | 98 |
| Eragrostis curvula | 38,113 | 101 | 92 | 59 | 98 |
| Ageratina altissima | 15,725 | 150 | 156 | 45 | 71 |
| Lolium perenne | 30,317 | 74 | 104 | 107 | 145 |
| Provinces | Total Area (Km2) a | Current (%) b | 2050 (%) c | 2070 (%) d | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Low | Moderate | High | Low | Moderate | High | Low | Moderate | High | ||
| Gangwon | 16,503.73 | 72.21 | 20.62 | 6.94 | 36.78 | 45.29 | 17.93 | 36.15 | 41.67 | 22.18 |
| Gyeonggi | 9810.10 | 30.64 | 33.41 | 35.68 | 16.34 | 60.51 | 23.14 | 21.34 | 53.88 | 24.78 |
| Incheon | 614.89 | 17.27 | 34.32 | 45.63 | 16.53 | 71.34 | 12.13 | 23.68 | 63.59 | 12.74 |
| Seoul | 605.70 | 6.18 | 23.51 | 70.31 | 21.95 | 75.84 | 2.21 | 32.75 | 67.09 | 0.16 |
| Gyeongsangbuk | 18,922.94 | 48.49 | 23.36 | 28.13 | 6.73 | 14.12 | 79.15 | 5.73 | 22.07 | 72.20 |
| Chungcheongbuk | 7415.68 | 50.23 | 20.74 | 29.02 | 8.09 | 18.47 | 73.44 | 4.73 | 23.22 | 72.05 |
| Chungcheongnam | 7637.76 | 39.75 | 20.54 | 39.60 | 8.20 | 23.58 | 68.22 | 11.81 | 35.51 | 52.69 |
| Sejong | 465.24 | 21.33 | 21.74 | 56.93 | 2.77 | 18.53 | 78.70 | 5.27 | 30.39 | 64.35 |
| Daejeon | 539.55 | 38.72 | 19.04 | 42.24 | 19.17 | 52.54 | 28.28 | 9.71 | 44.62 | 45.67 |
| Jeollabuk | 7716.82 | 47.17 | 16.15 | 36.67 | 9.28 | 22.07 | 68.65 | 6.25 | 26.89 | 66.87 |
| Daegu | 880.84 | 46.35 | 16.02 | 37.63 | 24.38 | 31.27 | 44.34 | 26.64 | 45.33 | 28.03 |
| Gyeongsangnam | 9809.70 | 39.45 | 25.01 | 35.46 | 18.30 | 17.55 | 64.15 | 7.53 | 32.56 | 59.92 |
| Ulsan | 1029.49 | 40.58 | 27.72 | 31.51 | 27.03 | 42.72 | 30.25 | 19.73 | 58.85 | 21.42 |
| Jeollanam | 10,180.34 | 46.09 | 17.38 | 36.24 | 20.79 | 32.52 | 46.69 | 18.59 | 47.81 | 33.60 |
| Busan | 673.03 | 29.86 | 16.86 | 53.28 | 80.23 | 18.14 | 1.63 | 16.04 | 76.83 | 7.13 |
| Gwangju | 498.36 | 24.85 | 10.77 | 64.37 | 19.74 | 29.56 | 50.70 | 19.17 | 34.78 | 46.05 |
| Jeju | 1674.96 | 45.33 | 54.26 | 0.18 | 57.71 | 42.29 | 0.00 | 59.84 | 40.15 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adhikari, P.; Lee, Y.-H.; Park, Y.-S.; Hong, S.-H. Assessment of the Spatial Invasion Risk of Intentionally Introduced Alien Plant Species (IIAPS) under Environmental Change in South Korea. Biology 2021, 10, 1169. https://doi.org/10.3390/biology10111169
Adhikari P, Lee Y-H, Park Y-S, Hong S-H. Assessment of the Spatial Invasion Risk of Intentionally Introduced Alien Plant Species (IIAPS) under Environmental Change in South Korea. Biology. 2021; 10(11):1169. https://doi.org/10.3390/biology10111169
Chicago/Turabian StyleAdhikari, Pradeep, Yong-Ho Lee, Yong-Soon Park, and Sun-Hee Hong. 2021. "Assessment of the Spatial Invasion Risk of Intentionally Introduced Alien Plant Species (IIAPS) under Environmental Change in South Korea" Biology 10, no. 11: 1169. https://doi.org/10.3390/biology10111169