
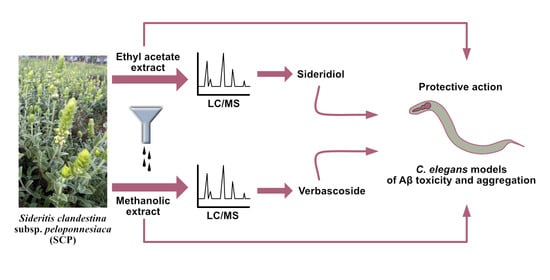
Beneficial Effects of Sideritis clandestina Extracts and Sideridiol against Amyloid β Toxicity

 , , , , , , , , , , ,
, , , , , , , , , , ,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Material and Preparation of Extracts and Compounds
2.1.1. Plant Material and Preparation of Extracts for Analysis and Characterization
2.1.2. Preparation of Extracts and Compounds for C. elegans Treatment
2.1.3. Dissolution of Sideridiol and Sample Preparation for TEM Experiments
2.2. LC-MS Analysis of the Extracts
2.3. Isolation of the Most Abundant Natural Products in the Two Extracts
2.4. Characterization of the Natural Products
2.5. C. elegans Strains and Culture
2.6. Paralysis Assays
2.7. Microscopy of CL2331 Animals and Image Analysis
2.8. Phenotypic Analysis
2.9. Protein Extraction and Dot Blotting
2.10. Synthesis and Disaggregation of Aβ42 Peptide
2.11. Transmission Electron Microscopy (TEM) and Negative Staining
2.12. Thioflavin T (ThT) Kinetic Assay
2.13. Molecular Dynamics Simulation
2.14. Statistical Analysis
3. Results
3.1. Phytochemical Composition of the Different SCP Extracts
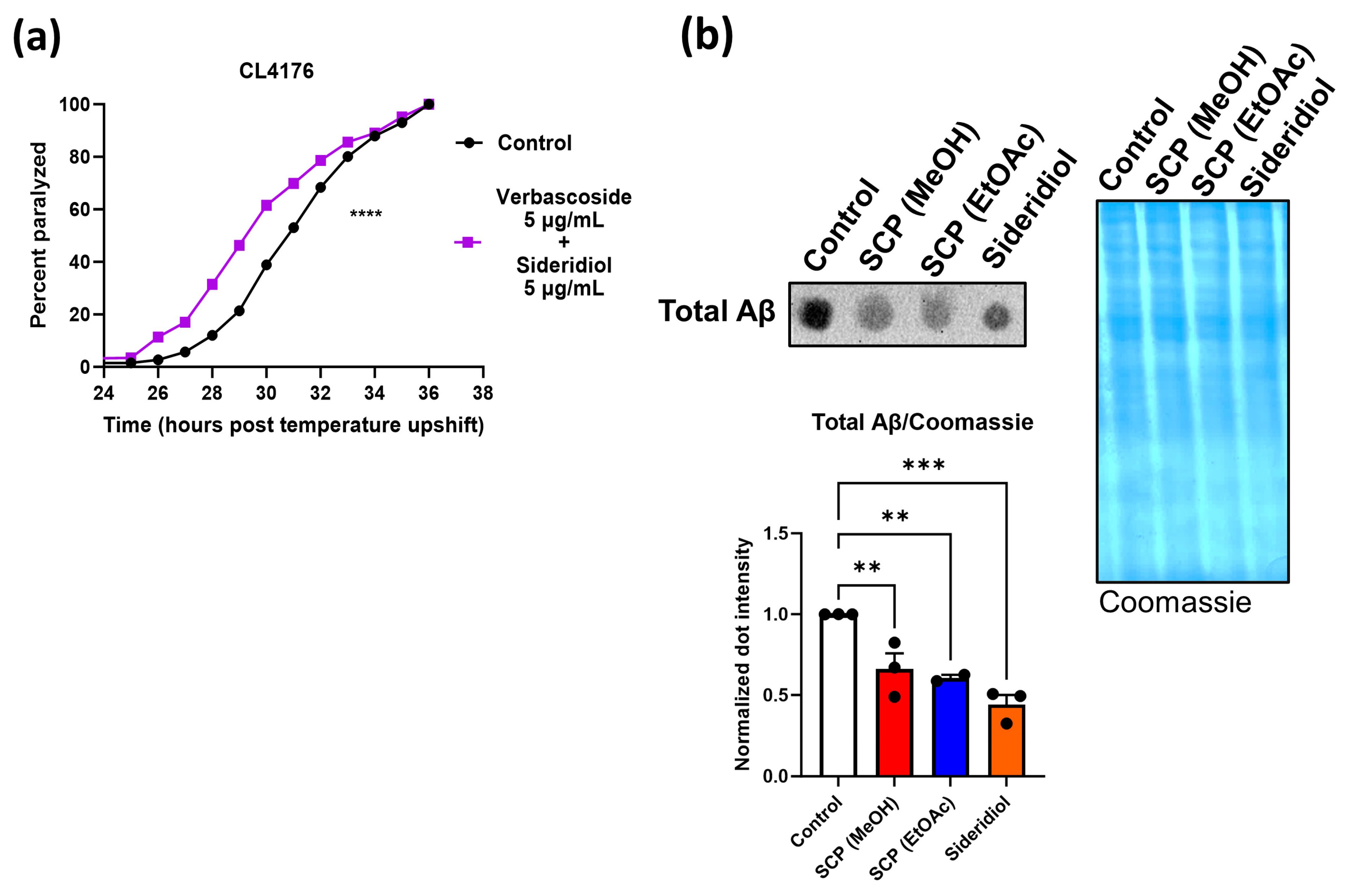
3.2. SCP Extracts Reduce Aβ Toxicity and Aggregation
3.3. Sideridiol Contributes against Aβ Toxicity and Aggregation
3.4. Verbascoside Contributes against Aβ Toxicity and Aggregation
3.5. Combination of Sideridiol and Verbascoside Do Not Act Synergistically
3.6. Sideridiol Affects the Amyloidogenicity of Aβ42
3.7. Sideridiol Affects the Tertiary Structural Stability of the Aβ42 Oligomer
3.8. Lack of Toxicity of SCP Extracts, Sideridiol and Verbascoside
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Scheltens, P.; Blennow, K.; Breteler, M.M.B.; de Strooper, B.; Frisoni, G.B.; Salloway, S.; Van der Flier, W.M. Alzheimer’s Disease. Lancet 2016, 388, 505–517. [Google Scholar] [CrossRef]
- Hou, Y.; Dan, X.; Babbar, M.; Wei, Y.; Hasselbalch, S.G.; Croteau, D.L.; Bohr, V.A. Ageing as a Risk Factor for Neurodegenerative Disease. Nat. Rev. Neurol. 2019, 15, 565–581. [Google Scholar] [CrossRef] [PubMed]
- Sadhukhan, P.; Saha, S.; Dutta, S.; Mahalanobish, S.; Sil, P.C. Nutraceuticals: An Emerging Therapeutic Approach against the Pathogenesis of Alzheimer’s Disease. Pharmacol. Res. 2018, 129, 100–114. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.S.; Khalid, M.; Kamal, M.A.; Younis, K. Study of Nutraceuticals and Phytochemicals for the Management of Alzheimer’s Disease: A Review. Curr. Neuropharmacol. 2021, 19, 1884–1895. [Google Scholar] [CrossRef]
- Wang, S.; Kong, X.; Chen, Z.; Wang, G.; Zhang, J.; Wang, J. Role of Natural Compounds and Target Enzymes in the Treatment of Alzheimer’s Disease. Molecules 2022, 27, 4175. [Google Scholar] [CrossRef] [PubMed]
- Chopra, B.; Dhingra, A.K. Natural Products: A Lead for Drug Discovery and Development. Phytother. Res. 2021, 35, 4660–4702. [Google Scholar] [CrossRef]
- Zarotsky, V.; Sramek, J.J.; Cutler, N.R. Galantamine Hydrobromide: An Agent for Alzheimer’s Disease. Am. J. Health-Syst. Pharm. AJHP Off. J. Am. Soc. Health-Syst. Pharm. 2003, 60, 446–452. [Google Scholar] [CrossRef]
- Żyżelewicz, D.; Kulbat-Warycha, K.; Oracz, J.; Żyżelewicz, K. Polyphenols and Other Bioactive Compounds of Sideritis Plants and Their Potential Biological Activity. Molecules 2020, 25, 3763. [Google Scholar] [CrossRef]
- Petrakou, K.; Iatrou, G.; Lamari, F.N. Ethnopharmacological Survey of Medicinal Plants Traded in Herbal Markets in the Peloponnisos, Greece. J. Herb. Med. 2020, 19, 100305. [Google Scholar] [CrossRef]
- Aneva, I.; Zhelev, P.; Kozuharova, E.; Danova, K.; Nabavi, S.F.; Behzad, S. Genus Sideritis, Section Empedoclia in Southeastern Europe and Turkey—Studies in Ethnopharmacology and Recent Progress of Biological Activities. Daru 2019, 27, 407–421. [Google Scholar] [CrossRef]
- Fraga, B.M. Phytochemistry and Chemotaxonomy of Sideritis Species from the Mediterranean Region. Phytochemistry 2012, 76, 7–24. [Google Scholar] [CrossRef]
- Todorova, M.; Trendafilova, A. Sideritis scardica Griseb., an Endemic Species of Balkan Peninsula: Traditional Uses, Cultivation, Chemical Composition, Biological Activity. J. Ethnopharmacol. 2014, 152, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Sideritis herba—Herbal Medicinal Product. Available online: https://www.ema.europa.eu/en/medicines/herbal/sideritis-herba (accessed on 12 December 2023).
- Strid, A.; Tan, K. Mountain Flora of Greece; Cambridge University Press: New York, NY, USA, 1986. [Google Scholar]
- Dimaki, V.D.; Zeliou, K.; Nakka, F.; Stavreli, M.; Bakratsas, I.; Papaioannou, L.; Iatrou, G.; Lamari, F.N. Characterization of Sideritis clandestina Subsp. peloponnesiaca Polar Glycosides and Phytochemical Comparison to Other Mountain Tea Populations. Molecules 2022, 27, 7613. [Google Scholar] [CrossRef] [PubMed]
- Dimaki, V.D.; Iatrou, G.; Lamari, F.N. Effect of Acidic and Enzymatic Pretreatment on the Analysis of Mountain Tea (Sideritis Spp.) Volatiles via Distillation and Ultrasound-Assisted Extraction. J. Chromatogr. A 2017, 1524, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Kostadinova, E.; Nikolova, D.; Alipieva, K.; Stefova, M.; Stefkov, G.; Evstatieva, L.; Matevski, V.; Bankova, V. Chemical Constituents of the Essential Oils of Sideritis scardica Griseb. and Sideritis raeseri Boiss and Heldr. from Bulgaria and Macedonia. Nat. Prod. Res. 2007, 21, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, T.; Li, J.; Xia, M.; Li, Y.; Wang, X.; Liu, C.; Zheng, T.; Chen, R.; Kan, D.; et al. Oxidative Stress and 4-Hydroxy-2-Nonenal (4-HNE): Implications in the Pathogenesis and Treatment of Aging-Related Diseases. J. Immunol. Res. 2022, 2022, 2233906. [Google Scholar] [CrossRef]
- Engelhart, M.J.; Geerlings, M.I.; Ruitenberg, A.; van Swieten, J.C.; Hofman, A.; Witteman, J.C.M.; Breteler, M.M.B. Dietary Intake of Antioxidants and Risk of Alzheimer Disease. JAMA 2002, 287, 3223–3229. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, X. Antioxidant Therapies for Alzheimer’s Disease. Oxid. Med. Cell. Longev. 2012, 2012, 472932. [Google Scholar] [CrossRef]
- Sinyor, B.; Mineo, J.; Ochner, C. Alzheimer’s Disease, Inflammation, and the Role of Antioxidants. J. Alzheimer’s Dis. Rep. 2020, 4, 175–183. [Google Scholar] [CrossRef]
- Collins, A.E.; Saleh, T.M.; Kalisch, B.E. Naturally Occurring Antioxidant Therapy in Alzheimer’s Disease. Antioxidants 2022, 11, 213. [Google Scholar] [CrossRef]
- Knörle, R. Extracts of Sideritis scardica as Triple Monoamine Reuptake Inhibitors. J. Neural Transm. 2012, 119, 1477–1482. [Google Scholar] [CrossRef] [PubMed]
- Dimpfel, W.; Schombert, L.; Feistel, B. Ex Vivo Characterization of the Action of Sideritis Extract Using Electrical Activity in the Rat Hippocampus Slice Preparation. Pharmacol. Pharm. 2016, 7, 407–416. [Google Scholar] [CrossRef]
- Chalatsa, I.; Arvanitis, D.A.; Mikropoulou, E.V.; Giagini, A.; Papadopoulou-Daifoti, Z.; Aligiannis, N.; Halabalaki, M.; Tsarbopoulos, A.; Skaltsounis, L.A.; Sanoudou, D. Beneficial Effects of Sideritis scardica and Cichorium spinosum against Amyloidogenic Pathway and Tau Misprocessing in Alzheimer’s Disease Neuronal Cell Culture Models. J. Alzheimer’s Dis. 2018, 64, 787–800. [Google Scholar] [CrossRef] [PubMed]
- Ververis, A.; Ioannou, K.; Kyriakou, S.; Violaki, N.; Panayiotidis, M.I.; Plioukas, M.; Christodoulou, K. Sideritis scardica Extracts Demonstrate Neuroprotective Activity against Aβ25–35 Toxicity. Plants 2023, 12, 1716. [Google Scholar] [CrossRef] [PubMed]
- Heiner, F.; Feistel, B.; Wink, M. Sideritis scardica Extracts Inhibit Aggregation and Toxicity of Amyloid-β in Caenorhabditis Elegans Used as a Model for Alzheimer’s Disease. PeerJ 2018, 6, e4683. [Google Scholar] [CrossRef] [PubMed]
- Hofrichter, J.; Krohn, M.; Schumacher, T.; Lange, C.; Feistel, B.; Walbroel, B.; Pahnke, J. Sideritis Spp. Extracts Enhance Memory and Learning in Alzheimer’s β-Amyloidosis Mouse Models and Aged C57Bl/6 Mice. J. Alzheimer’s Dis. 2016, 53, 967–980. [Google Scholar] [CrossRef]
- Kokras, N.; Poulogiannopoulou, E.; Sotiropoulos, M.G.; Paravatou, R.; Goudani, E.; Dimitriadou, M.; Papakonstantinou, E.; Doxastakis, G.; Perrea, D.N.; Hloupis, G.; et al. Behavioral and Neurochemical Effects of Extra Virgin Olive Oil Total Phenolic Content and Sideritis Extract in Female Mice. Molecules 2020, 25, 5000. [Google Scholar] [CrossRef]
- Lazarova, M.I.; Tancheva, L.P.; Tasheva, K.N.; Denev, P.N.; Uzunova, D.N.; Stefanova, M.O.; Tsvetanova, E.R.; Georgieva, A.P.; Kalfin, R.E. Effects of Sideritis scardica Extract on Scopolamine-Induced Learning and Memory Impairment in Mice. J. Alzheimer’s Dis. 2023, 92, 1289–1302. [Google Scholar] [CrossRef]
- Linardaki, Z.I.; Vasilopoulou, C.G.; Constantinou, C.; Iatrou, G.; Lamari, F.N.; Margarity, M. Differential Antioxidant Effects of Consuming Tea from Sideritis clandestina Subsp. peloponnesiaca on Cerebral Regions of Adult Mice. J. Med. Food 2011, 14, 1060–1064. [Google Scholar] [CrossRef]
- Vasilopoulou, C.G.; Kontogianni, V.G.; Linardaki, Z.I.; Iatrou, G.; Lamari, F.N.; Nerantzaki, A.A.; Gerothanassis, I.P.; Tzakos, A.G.; Margarity, M. Phytochemical Composition of “Mountain Tea” from Sideritis clandestina Subsp. clandestina and Evaluation of Its Behavioral and Oxidant/Antioxidant Effects on Adult Mice. Eur. J. Nutr. 2013, 52, 107–116. [Google Scholar] [CrossRef]
- Wightman, E.L.; Jackson, P.A.; Khan, J.; Forster, J.; Heiner, F.; Feistel, B.; Suarez, C.G.; Pischel, I.; Kennedy, D.O. The Acute and Chronic Cognitive and Cerebral Blood Flow Effects of a Sideritis scardica (Greek Mountain Tea) Extract: A Double Blind, Randomized, Placebo Controlled, Parallel Groups Study in Healthy Humans. Nutrients 2018, 10, 955. [Google Scholar] [CrossRef]
- Behrendt, I.; Schneider, I.; Schuchardt, J.P.; Bitterlich, N.; Hahn, A. Effect of an Herbal Extract of Sideritis scardica and B-Vitamins on Cognitive Performance under Stress: A Pilot Study. Int. J. Phytomed. 2016, 8, 95–103. [Google Scholar] [CrossRef]
- Dimpfel, W.; Biller, A.; Suliman, S.; Chiegoua Dipah, G.N. Psychophysiological Effects of a Combination of Sideritis and Bacopa Extract (MemoLoges®) in 32 Subjects Suffering from Mild Cognitive Impairment. A Double-Blind, Randomized, Placebo-Controlled, 2-Armed Study with Parallel Design. Adv. Alzheimer’s Dis. 2016, 5, 103–125. [Google Scholar] [CrossRef]
- Griffin, E.F.; Caldwell, K.A.; Caldwell, G.A. Genetic and Pharmacological Discovery for Alzheimer’s Disease Using Caenorhabditis elegans. ACS Chem. Neurosci. 2017, 8, 2596–2606. [Google Scholar] [CrossRef] [PubMed]
- Kilic, T.; Topcu, G.; Goren, A.C.; Aydogmus, Z.; Karagoz, A.; Yildiz, Y.K.; Aslan, I. Ent-Kaurene Diterpenoids from Sideritis lycia with Antiviral and Cytotoxic Activities. Rec. Nat. Prod. 2020, 14, 256–268. [Google Scholar] [CrossRef]
- Venditti, A.; Bianco, A.; Maggi, F.; Nicoletti, M. Polar Constituents Composition of Endemic Sideritis italica (MILL.) GREUTER et Burter from Central Italy. Nat. Prod. Res. 2013, 27, 1408–1412. [Google Scholar] [CrossRef] [PubMed]
- Tomou, E.-M.; Chatziathanasiadou, M.V.; Chatzopoulou, P.; Tzakos, A.G.; Skaltsa, H. NMR-Based Chemical Profiling, Isolation and Evaluation of the Cytotoxic Potential of the Diterpenoid Siderol from Cultivated Sideritis euboea Heldr. Molecules 2020, 25, 2382. [Google Scholar] [CrossRef] [PubMed]
- Demirtas, I.; Ayhan, B.; Sahin, A.; Aksit, H.; Elmastas, M.; Telci, I. Antioxidant Activity and Chemical Composition of Sideritis libanotica Labill. Ssp. linearis (Bentham) Borm. (Lamiaceae). Nat. Prod. Res. 2011, 25, 1512–1523. [Google Scholar] [CrossRef]
- Brenner, S. The Genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef]
- Papaevgeniou, N.; Hoehn, A.; Tur, J.A.; Klotz, L.-O.; Grune, T.; Chondrogianni, N. Sugar-Derived AGEs Accelerate Pharyngeal Pumping Rate and Increase the Lifespan of Caenorhabditis elegans. Free Radic. Res. 2019, 53, 1056–1067. [Google Scholar] [CrossRef]
- Vasilopoulou, M.A.; Gioran, A.; Theodoropoulou, M.; Koutsaviti, A.; Roussis, V.; Ioannou, E.; Chondrogianni, N. Healthspan Improvement and Anti-Aggregation Effects Induced by a Marine-Derived Structural Proteasome Activator. Redox Biol. 2022, 56, 102462. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 12 December 2023).
- RStudio: Integrated Development Environment for R. Available online: www.rstudio.com (accessed on 12 December 2023).
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- 2BEG-3D Structure of Alzheimer’s Abeta(1–42) Fibrils. Available online: https://www.rcsb.org/structure/2BEG (accessed on 12 December 2023).
- Lührs, T.; Ritter, C.; Adrian, M.; Riek-Loher, D.; Bohrmann, B.; Döbeli, H.; Schubert, D.; Riek, R. 3D Structure of Alzheimer’s Amyloid-β(1–42) Fibrils. Proc. Natl. Acad. Sci. USA 2005, 102, 17342–17347. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem in 2021: New Data Content and Improved Web Interfaces. Nucleic Acids Res. 2021, 49, D1388–D1395. [Google Scholar] [CrossRef]
- Kutzner, C.; Páll, S.; Fechner, M.; Esztermann, A.; de Groot, B.L.; Grubmüller, H. More Bang for Your Buck: Improved Use of GPU Nodes for GROMACS 2018. J. Comput. Chem. 2019, 40, 2418–2431. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High Performance Molecular Simulations through Multi-Level Parallelism from Laptops to Supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef]
- Huang, J.; MacKerell, A.D.J. CHARMM36 All-Atom Additive Protein Force Field: Validation Based on Comparison to NMR Data. J. Comput. Chem. 2013, 34, 2135–2145. [Google Scholar] [CrossRef]
- Vanommeslaeghe, K.; Hatcher, E.; Acharya, C.; Kundu, S.; Zhong, S.; Shim, J.; Darian, E.; Guvench, O.; Lopes, P.; Vorobyov, I.; et al. CHARMM General Force Field: A Force Field for Drug-like Molecules Compatible with the CHARMM All-Atom Additive Biological Force Fields. J. Comput. Chem. 2010, 31, 671–690. [Google Scholar] [CrossRef]
- Bussi, G.; Donadio, D.; Parrinello, M. Canonical Sampling through Velocity Rescaling. J. Chem. Phys. 2007, 126, 14101. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; DiNola, A.; Haak, J.R. Molecular Dynamics with Coupling to an External Bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef]
- Parrinello, M.; Rahman, A. Polymorphic Transitions in Single Crystals: A New Molecular Dynamics Method. J. Appl. Phys. 1981, 52, 7182–7190. [Google Scholar] [CrossRef]
- Nosé, S.; Klein, M.L. Constant Pressure Molecular Dynamics for Molecular Systems. Mol. Phys. 1983, 50, 1055–1076. [Google Scholar] [CrossRef]
- Piozzi, F.; Bruno, M.; Rosselli, S.; Maggio, A. The Diterpenoids from the Genus Sideritis. In Studies in Natural Products Chemistry; Atta-ur-Rahman, B.T.-S., Ed.; Elsevier: Amsterdam, The Netherlands, 2006; Volume 33, pp. 493–540. ISBN 1572-5995. [Google Scholar]
- Link, C.D.; Taft, A.; Kapulkin, V.; Duke, K.; Kim, S.; Fei, Q.; Wood, D.E.; Sahagan, B.G. Gene Expression Analysis in a Transgenic Caenorhabditis elegans Alzheimer’s Disease Model. Neurobiol. Aging 2003, 24, 397–413. [Google Scholar] [CrossRef] [PubMed]
- McColl, G.; Roberts, B.R.; Pukala, T.L.; Kenche, V.B.; Roberts, C.M.; Link, C.D.; Ryan, T.M.; Masters, C.L.; Barnham, K.J.; Bush, A.I.; et al. Utility of an Improved Model of Amyloid-Beta (Aβ1–42) Toxicity in Caenorhabditis elegans for Drug Screening for Alzheimer’s Disease. Mol. Neurodegener. 2012, 7, 57. [Google Scholar] [CrossRef]
- Link, C.D.; Fonte, V.; Roberts, C.M.; Hiester, B.; Silverman, M.A.; Stein, G.H. The Beta Amyloid Peptide Can Act as a Modular Aggregation Domain. Neurobiol. Dis. 2008, 32, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Blake, C.C.F.; Sunde, M. From the Globular to the Fibrous State: Protein Structure and Structural Conversion in Amyloid Formation. Q. Rev. Biophys. 1998, 31, 1–39. [Google Scholar] [CrossRef]
- LeVine, H. Thioflavine T Interaction with Amyloid β-Sheet Structures. Amyloid 1995, 2, 1–6. [Google Scholar] [CrossRef]
- O’Brien, R.J.; Wong, P.C. Amyloid Precursor Protein Processing and Alzheimer’s Disease. Annu. Rev. Neurosci. 2011, 34, 185–204. [Google Scholar] [CrossRef]
- Kabsch, W.; Sander, C. Dictionary of Protein Secondary Structure: Pattern Recognition of Hydrogen-Bonded and Geometrical Features. Biopolymers 1983, 22, 2577–2637. [Google Scholar] [CrossRef] [PubMed]
- Touw, W.G.; Baakman, C.; Black, J.; te Beek, T.A.H.; Krieger, E.; Joosten, R.P.; Vriend, G. A Series of PDB-Related Databanks for Everyday Needs. Nucleic Acids Res. 2015, 43, D364–D368. [Google Scholar] [CrossRef]
- Soto, C.; Castaño, E.M.; Frangione, B.; Inestrosa, N.C. The Alpha-Helical to Beta-Strand Transition in the Amino-Terminal Fragment of the Amyloid Beta-Peptide Modulates Amyloid Formation. J. Biol. Chem. 1995, 270, 3063–3067. [Google Scholar] [CrossRef]
- Leung, M.C.K.; Williams, P.L.; Benedetto, A.; Au, C.; Helmcke, K.J.; Aschner, M.; Meyer, J.N. Caenorhabditis Elegans: An Emerging Model in Biomedical and Environmental Toxicology. Toxicol. Sci. 2008, 106, 5–28. [Google Scholar] [CrossRef] [PubMed]
- Hunt, P.R. The C. elegans Model in Toxicity Testing. J. Appl. Toxicol. 2017, 37, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Rollins, J.A.; Howard, A.C.; Dobbins, S.K.; Washburn, E.H.; Rogers, A.N. Assessing Health Span in Caenorhabditis elegans: Lessons From Short-Lived Mutants. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 473–480. [Google Scholar] [CrossRef]
- Ewbank, J.J.; Zugasti, O. C. elegans: Model Host and Tool for Antimicrobial Drug Discovery. Dis. Model. Mech. 2011, 4, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Xiong, C.; Kornfeld, K. Measurements of Age-Related Changes of Physiological Processes That Predict Lifespan of Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2004, 101, 8084–8089. [Google Scholar] [CrossRef] [PubMed]
- Topçu, G.; Ertaş, A.; Öztürk, M.; Dinçel, D.; Kılıç, T.; Halfon, B. Ent-Kaurane Diterpenoids Isolated from Sideritis congesta. Phytochem. Lett. 2011, 4, 436–439. [Google Scholar] [CrossRef]
- Topcu, G.; Goren, A.C. Biological Activity of Diterpenoids Isolated from Anatolian Lamiaceae Plants. Rec. Nat. Prod. 2007, 1, 1–16. [Google Scholar]
- Bondì, M.L.; Bruno, M.; Piozzi, F.; Baser, K.H.C.; Simmonds, M.S. Diversity and Antifeedant Activity of Diterpenes from Turkish Species of Sideritis. Biochem. Syst. Ecol. 2000, 28, 299–303. [Google Scholar] [CrossRef]
- Çarıkçı, S.; Kılıç, T.; Gören, A.C.; Dirmenci, T.; Alim Toraman, G.Ö.; Topçu, G. Chemical Profile of the Anatolian Sideritis Species with Bioactivity Studies. Pharm. Biol. 2023, 61, 1484–1511. [Google Scholar] [CrossRef] [PubMed]
- Pardo, F.; Perich, F.; Villarroel, L.; Torres, R. Isolation of Verbascoside, an Antimicrobial Constituent of Buddleja globosa Leaves. J. Ethnopharmacol. 1993, 39, 221–222. [Google Scholar] [CrossRef]
- Avila, J.G.; de Liverant, J.G.; Martínez, A.; Martínez, G.; Muñoz, J.L.; Arciniegas, A.; Romo de Vivar, A. Mode of Action of Buddleja cordata Verbascoside against Staphylococcus aureus. J. Ethnopharmacol. 1999, 66, 75–78. [Google Scholar] [CrossRef]
- Speranza, L.; Franceschelli, S.; Pesce, M.; Menghini, L.; Patruno, A.; Vinciguerra, I.; De Lutiis, M.A.; Felaco, M.; Felaco, P.; Grilli, A. Anti-Inflammatory Properties of the Plant Verbascum Mallophorum. J. Biol. Regul. Homeost. Agents 2009, 23, 189–195. [Google Scholar] [PubMed]
- Wang, H.; Xu, Y.; Yan, J.; Zhao, X.; Sun, X.; Zhang, Y.; Guo, J.; Zhu, C. Acteoside Protects Human Neuroblastoma SH-SY5Y Cells against Beta-Amyloid-Induced Cell Injury. Brain Res. 2009, 1283, 139–147. [Google Scholar] [CrossRef]
- Wang, C.; Cai, X.; Wang, R.; Zhai, S.; Zhang, Y.; Hu, W.; Zhang, Y.; Wang, D. Neuroprotective Effects of Verbascoside against Alzheimer’s Disease via the Relief of Endoplasmic Reticulum Stress in Aβ-Exposed U251 Cells and APP/PS1 Mice. J. Neuroinflamm. 2020, 17, 309. [Google Scholar] [CrossRef] [PubMed]
- Aimaiti, M.; Wumaier, A.; Aisa, Y.; Zhang, Y.; Xirepu, X.; Aibaidula, Y.; Lei, X.; Chen, Q.; Feng, X.; Mi, N. Acteoside Exerts Neuroprotection Effects in the Model of Parkinson’s Disease via Inducing Autophagy: Network Pharmacology and Experimental Study. Eur. J. Pharmacol. 2021, 903, 174136. [Google Scholar] [CrossRef]
- Gao, L.; Wang, D.; Ren, J.; Tan, X.; Chen, J.; Kong, Z.; Nie, Y.; Yan, M. Acteoside Ameliorates Learning and Memory Impairment in APP/PS1 Transgenic Mice by Increasing Aβ Degradation and Inhibiting Tau Hyperphosphorylation. Phytother. Res. 2023. online ahead of print. [Google Scholar] [CrossRef]
- Li, R.; Tao, M.; Wu, T.; Zhuo, Z.; Xu, T.; Pan, S.; Xu, X. A Promising Strategy for Investigating the Anti-Aging Effect of Natural Compounds: A Case Study of Caffeoylquinic Acids. Food Funct. 2021, 12, 8583–8593. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| tR (min) | Ions (m/z) in Positive Ionization | M.W. | Compound | mg Sideridiol equiv./g Dry Extract | |
|---|---|---|---|---|---|
| 1 | 1.73 | 413 [M + Na]+ | 390 | Ajugoside # | nq |
| 2 | 9.10 | 321 [M + H]+/343 [M + Na]+/ 663 [2M + Na]+ | 320 | Sideroxol # | 6.08 |
| 3 | 13.41 | 385 [M + Na]+/747 [2M + Na]+ | 362 | Unknown | 14.12 |
| 4 | 14.63 | 327 [M + Na]+/631 [2M + Na]+ | 304 | Unknown | nq |
| 5 | 16.41 | 327 [M + Na]+/631 [2M + Na]+ | 304 | Unknown | 8.36 |
| 6 | 16.76 | 327 [M + Na]+/631 [2M + Na]+ | 304 | Unknown | 2.09 |
| 7 | 17.62 | 327 [M + Na]+, 631 [2M + Na]+ | 304 | Sideridiol # | 111.59 |
| 8 | 25.53 | 427 [M + Na]+/809 [2M + H]+/831 [2M + Na]+ | 404 | Unknown | 47.44 |
| 9 | 33.50 | 369 [M + Na]+/715 [2M + Na]+ | 346 | Siderol # | 272.89 |
| 10 | 40.82 | 469 [M + Na]+/915 [2M + Na]+ | 446 | 3,7,18-triacetyl-foliol 1 | 27.92 |
| 11 | 56.02 | 411 [M + Na]+/799 [2M + Na]+ | 388 | Unknown | nq |
| tR (min) | Ions (m/z) in Negative Ionization | M.W. | Compound | mg Rutin equiv./g Dry Extract | |
|---|---|---|---|---|---|
| 1 | 3.24 | 191/383/533 | - | Unknown | 4.26 |
| 2 | 6.38 | 523 [M − H]−/583 [M + Hac − H]−/1047 [2M − H]− | 524 | Melittoside # | 4.37 |
| 3 | 17.92 | 191 (quinic acid)/353 [M − H]−/707 [2M − H]−/729 [2M − 2H + Na]− | 354 | Chlorogenic acid # | 10.35 |
| 4 | 23.91 | 417 [M − H]− | 418 | 7-O-acetyl-8-epi-loganic acid # | nq |
| 5 | 26.79 | 392 [M − 2H]2−/785 [M − H]− | 786 | Unknown | 3.23 |
| 6 | 27.11 | 377 [M − 2H]2−/755 [M − H]− | 756 | Forsythoside B ή Lavandulofolioside | 9.55 |
| 7 | 28.06 | 311 [M − 2H]2−/623 [M − H]−/1248 [2M − H]− | 624 | Verbascoside # | 15.61 |
| 8 | 29.01 | 625 [M − H]− | 626 | All-Glc-HYP | 2.69 |
| 9 | 32.36 | 769 [M − H]− | 770 | Allysonoside | 3.43 |
| 10 | 34.03 | 637 [M1 − H]−/651 [M2 − H]−/623 [M3 − H]− | 638 (M1) and 652 (M2) and 624 (M3) | M1: Leucoseptoside isomer/M2: AcO-All-Glc-ISC or AcO-All-Glc-LUT/M3: isoverbascoside # | 9.10 |
| 11 | 35.71 | 639 [M − H]−/1279 [2M − H]− | 640 | All-Glc-HYP-Me | nq |
| 12 | 36.18 | 667 [M − H]− | 668 | AcO-All-Glc-HYP | 8.79 |
| 13 | 42.43 | 651 [M − H]−/1303 [2M − H]− | 652 | AcO-All-Glc-ISC or AcO-All-Glc-LUT | 9.62 |
| 14 | 43.26 | 681 [M − H]−/1364 [2M − H]− | 682 | AcO-All-Glc-HYP-Me | 6.54 |
| 15 | 47.72 | 709 [M − H]− | 710 | (AcO)2-All-Glc-HYP | 2.64 |
| 16 | 49.25 | 693 [M1 − H]−/723 [M2 − H]− | 694 (M1) and 724 (M2) | M1: (AcO)2-All-Glc-ISC/M2: (AcO)2-All-Glc-HYP-Me | nq |
| Developmental Time a | Fertility b | Egg Lethality c | Pharyngeal Pumping d | Defecation e | |
|---|---|---|---|---|---|
| DMSO | 51.27 ± 0.12 | 269.8 ± 11.81 | 0 | 279.4 ± 3.06 | 47.45 ± 0.71 |
| 10 μg/mL SCP (EtOAc) | 52 ± 0.08 | 257.6 ± 13.70 | 0 | 288.2 ± 2.76 * | 41.13 ± 0.57 *** |
| Developmental Time a | Fertility b | Egg Lethality c | Pharyngeal Pumping d | Defecation e | |
|---|---|---|---|---|---|
| DMSO | 52.15 ± 0.07 | 265.2 ± 8.81 | 0 | 311.64 ± 2.72 | 45.67 ± 0.50 |
| 5 μg/mL SCP (MeOH) | 51.7 ± 0.08 | 276.3 ± 7.222 | 0 | 312.8 ± 2.52 | 43.36 ± 0.49 *** |
| Developmental Time a | Fertility b | Egg Lethality c | Pharyngeal Pumping d | Defecation e | |
|---|---|---|---|---|---|
| Control | 51.08 ± 0.14 | 279.4 ± 7.81 | 0 | 292.9 ± 1.87 | 43.94 ± 0.74 |
| 5 μg/mL Sideridiol | 51.41 ± 0.16 | 277.0 ± 8.62 | 0 | 289.4 ± 2.41 | 44.30 ± 0.43 |
| Developmental Time a | Fertility b | Egg Lethality c | Pharyngeal Pumping d | Defecation e | |
|---|---|---|---|---|---|
| Control | 51.67 ± 0.47 | 305.80 ± 18.12 | 0.75 ± 0.59 | 280.7 ± 3.92 | 42.73 ± 0.62 |
| 5 μg/mL Verbascoside | 52.67 ± 0.47 | 299.80 ± 13.67 | 1.12 ± 0.41 | 289.2 ± 4.04 | 47.40 ± 0.39 **** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gioran, A.; Paikopoulos, Y.; Panagiotidou, E.; Rizou, A.E.I.; Nasi, G.I.; Dimaki, V.D.; Vraila, K.D.; Bezantakou, D.S.; Spatharas, P.M.; Papandreou, N.C.; et al. Beneficial Effects of Sideritis clandestina Extracts and Sideridiol against Amyloid β Toxicity. Antioxidants 2024, 13, 261. https://doi.org/10.3390/antiox13030261
Gioran A, Paikopoulos Y, Panagiotidou E, Rizou AEI, Nasi GI, Dimaki VD, Vraila KD, Bezantakou DS, Spatharas PM, Papandreou NC, et al. Beneficial Effects of Sideritis clandestina Extracts and Sideridiol against Amyloid β Toxicity. Antioxidants. 2024; 13(3):261. https://doi.org/10.3390/antiox13030261
Chicago/Turabian StyleGioran, Anna, Yiorgos Paikopoulos, Eleni Panagiotidou, Aikaterini E. I. Rizou, Georgia I. Nasi, Virginia D. Dimaki, Konstantina D. Vraila, Dimitra S. Bezantakou, Panagiotis M. Spatharas, Nikos C. Papandreou, and et al. 2024. "Beneficial Effects of Sideritis clandestina Extracts and Sideridiol against Amyloid β Toxicity" Antioxidants 13, no. 3: 261. https://doi.org/10.3390/antiox13030261