
A Novel Disease (Water Bubble Disease) of the Giant Freshwater Prawn Macrobrachium rosenbergii Caused by Citrobacter freundii: Antibiotic Treatment and Effects on the Antioxidant Enzyme Activity and Immune Responses


Abstract
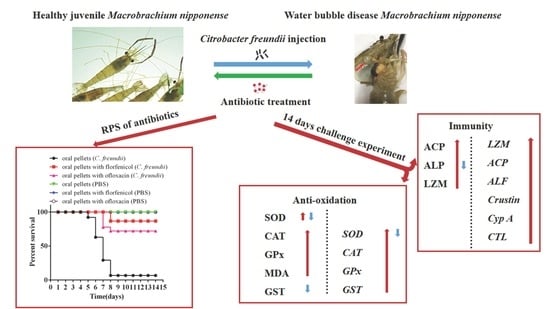
:
1. Introduction
2. Materials and Methods
2.1. Strain Isolation and Identification
2.2. Gene Sequencing and Phylogenetic Analysis
2.3. Effects of Temperature, pH, and Salinity on the Growth of Citrobacter freundii
2.4. Antibiotic Susceptibility Test
2.5. Experimental Challenge
2.6. Median Lethal Dose Determination
2.7. Antibiotic Treatment
2.8. Immune and Anti-Oxidization Parameter Changes in Hemolymph and Hepatopancreas
2.9. Expressions of Immune-Related Genes in Hepatopancreas
2.10. Statistical Analysis
3. Results
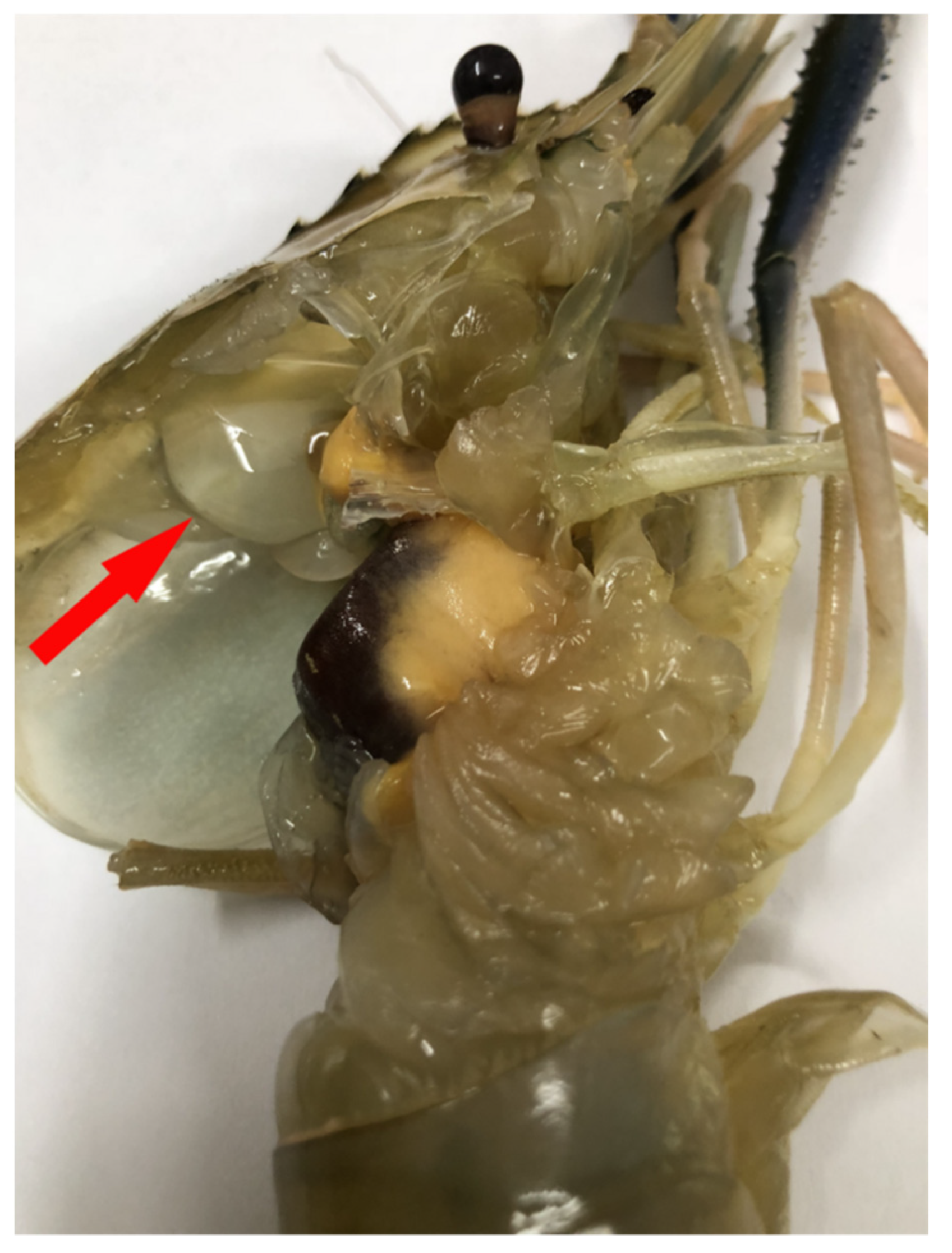
3.1. Clinical Symptoms of WBD in M. rosenbergii
3.2. Molecular Identification and Biochemical Characterization
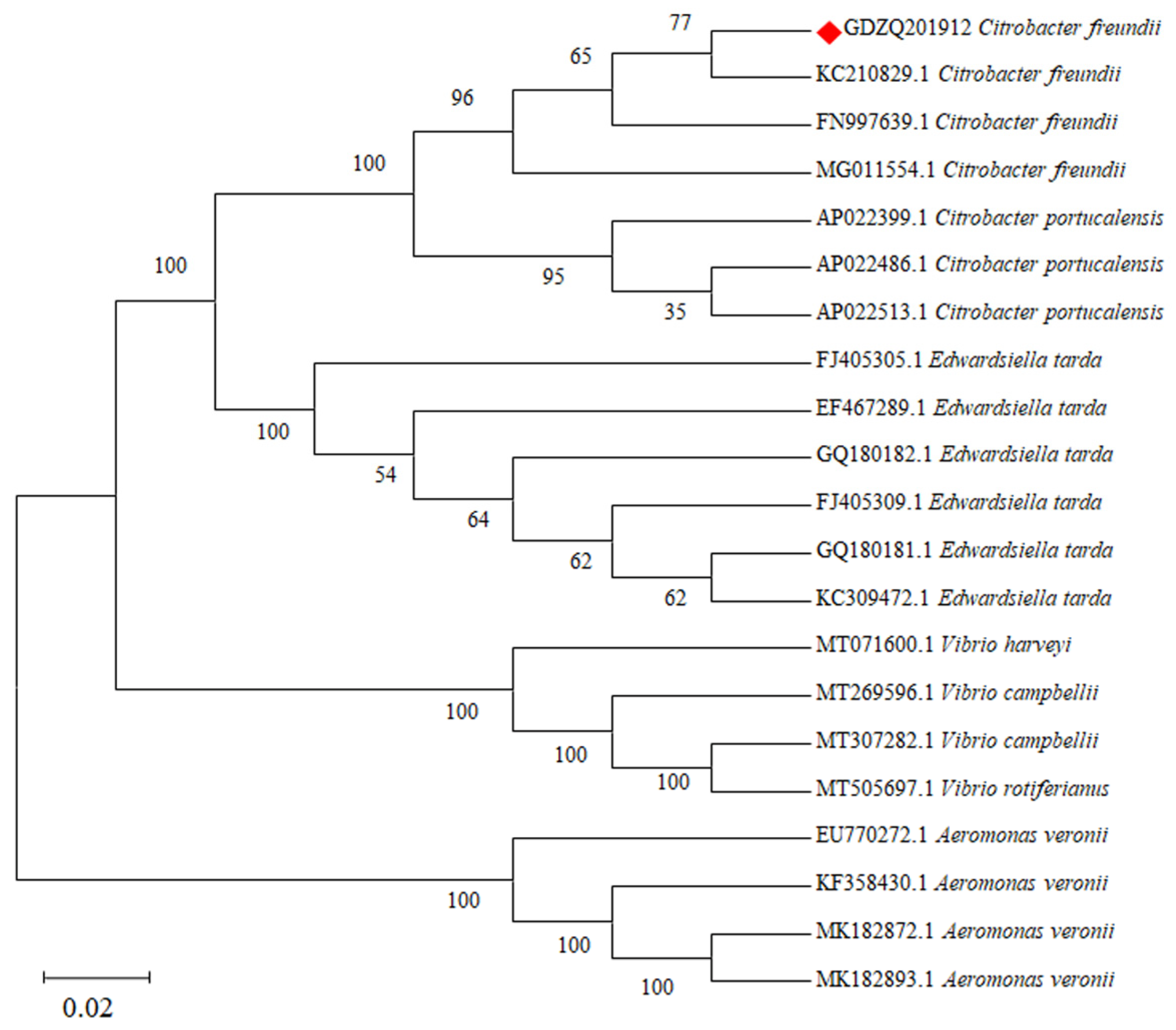
3.3. Phylogenetic Analysis
3.4. Effects of Temperature, pH, and Salinity on the Growth of C. freundii
3.5. Experimental Challenge
3.6. LD50 of Strain GDZQ201912 in M. rosenbergii
3.7. Antibiogram Tests of Strain GDZQ201912
3.8. Antibiotic Treatment
3.9. Immune and Antioxidant Enzyme Activity Changes in Hemolymph
3.10. Immune and Antioxidant Enzyme Activity Changes in Hepatopancreas
3.11. Expression of Immune-Related Genes in Hepatopancreas after the Challenge with C. freundii
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nhan, D.T.; Wille, M.; Sorgeloos, P. Comparison of reproductive performance and offspring quality of giant freshwater prawn (Macrobrachium rosenbergii) broodstock from different regions. Aquaculture 2009, 298, 36–42. [Google Scholar] [CrossRef]
- New, M. Freshwater prawn farming: Global status, recent research and a glance at the future. Aquac. Res. 2005, 36, 210–230. [Google Scholar] [CrossRef]
- Kumaresan, V.; Palanisamy, R.; Pasupuleti, M.; Arockiaraj, J. Impacts of environmental and biological stressors on immune system of Macrobrachium rosenbergii. Rev. Aquacult. 2017, 9, 283–307. [Google Scholar] [CrossRef]
- FAO Data form “Fisheries and Aquaculture Software Fishstatj-Software for Fishery and Aquaculture Statistical Time Series”. FAO Fisheries Division 2021, Rome, Italy. Available online: http://www.fao.org/fishery/statistics/software/fishstatj/en (accessed on 18 November 2021).
- Tiruvayipati, S.; Bhassu, S. Host, pathogen and the environment: The case of Macrobrachium rosenbergii, Vibrio parahaemolyticus and magnesium. Gut. Pathog. 2016, 8, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.J.; Miao, Z.; Li, X.X.; Chen, N.; Gu, W.W.; Liu, X.D.; Yang, H.; Wei, W.H.; Zhang, X.J. Pathogenicity of non-O1/O139 Vibrio cholerae and its induced immune response in Macrobrachium rosenbergii. Fish. Shellfish. Immunol. 2019, 92, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y.; Wang, L.F.; Zhuang, H.; Li, X.X.; Gao, X.J.; An, Z.H.; Liu, X.D.; Yang, H.; Wei, W.Z.; Zhang, X.J. Excessive use of enrofloxacin leads to growth inhibition of juvenile giant freshwater prawn Macrobrachium rosenbergii. Ecotoxicol. Environ. Saf. 2019, 169, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.J.; Yang, X.Q.; He, W.R.; Xiong, Y.A.; Liu, J.; Dai, Z.M. Involvement of tetraspanin 8 in the innate immune response of the giant prawn, Macrobrachium rosenbergii. Fish. Shellfish. Immunol. 2019, 86, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Tao, B.H.; Shi, H.R.; Huang, J.W.; Wang, G. Studied on yellow and black gills of Macrobrachium rosenbergii caused by Pseudoonas. Acta Sci. Nat. Univ. Sunyatseni 2000, 1, 255–259. [Google Scholar]
- Cheng, W.; Chen, J. Isolation and characterization of an Enterococcus-like bacterium causing muscle necrosis and mortality in Macrobrachium rosenbergii in Taiwan. Dis. Aquat. Organ. 1998, 34, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramalingam, K.; Ramarani, S. Effect of Pseudomonas aeruginosa on the giant freshwater prawn, Macrobrachium rosenbergii histopathological and electron microscopic study. J. Environ. Biol. 2007, 28, 627–635. [Google Scholar] [PubMed]
- Jayaprakash, N.S.; Rejish, K.V.J.; Philip, R.; Bright, S.I.S. Vibrios associated with Macrobrachium rosenbergii (De Man, 1879) larvae from three hatcheries on the Indian southwest coast. Aquac. Res. 2006, 37, 351–358. [Google Scholar] [CrossRef]
- Sung, H.H.; Hwang, S.F.; Tasi, F.M. Responses of giant freshwater prawn (Macrobrachium rosenbergii) to challenge by two strains of Aeromonas spp. J. Invertebr. Pathol. 2000, 76, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; He, X.; An, Z.; Sun, W.; Chen, N.; Gao, X.; Li, X.X.; Zhang, X. Citrobacter freundii infection in red swamp crayfish (Procambarus clarkii) and host immune-related gene expression profiles. Aquaculture 2020, 515, 734499. [Google Scholar] [CrossRef]
- Li, Y.; Zhan, F.; Li, F.; Lu, Z.; Shi, F.; Xu, Z.; Yang, Y.C.; Zhao, L.J.; Qin, Z.D.; Lin, L. Immune function of cytosolic manganese superoxide dismutase from Macrobrachium rosenbergii in response to bacterial infection. Aquaculture 2021, 541, 736771. [Google Scholar] [CrossRef]
- Butprom, S.; Phumkhachorn, P.; Rattanachaikunsopon, P. Effect of Lactobacillus plantarum C014 on innate immune response and disease resistance against Aeromonas hydrophila in hybrid catfish. Sci. World. J. 2013, 2013, 392523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behera, B.K.; Paria, P.; Das, A.; Das, B.K. Molecular identification and pathogenicity study of virulent Citrobacter freundii associated with mortality of farmed Labeo rohita (Hamilton 1822), in India. Aquaculture 2022, 547, 737437. [Google Scholar] [CrossRef]
- Li, Y.; Yan, L.; Kong, X.; Chen, J.; Zhang, H. Cloning, expression, and characterization of a novel superoxide dismutase from deep-sea sea cucumber. Int. J. Bio. Macromol. 2020, 163, 1875–1883. [Google Scholar] [CrossRef] [PubMed]
- Cetinkale, O.; Belce, A.; Konukoglu, D.; Senyuva, C.; Gumustas, M.K.; Tas, T. Evaluation of lipid peroxidation and total antioxidant status in plasma of rats following thermal injury. Burns 1997, 23, 114–116. [Google Scholar] [CrossRef]
- Lopes, P.A.; Pinheiro, T.; Santos, M.C.; da Luz Mathias, M.; Collares-Pereira, M.J.; Viegas-Crespo, A.M. Response of antioxidant enzymes in freshwater fish populations (Leuciscus alburnoides complex) to inorganic pollutants exposure. Sci. Total Environ. 2001, 280, 153–163. [Google Scholar] [CrossRef]
- Yin, F.; Gong, H.; Ke, Q.; Li, A. Stress, antioxidant defence and mucosal immune responses of the large yellow croaker Pseudosciaena crocea challenged with Cryptocaryon irritans. Fish. Shellfish. Immunol. 2015, 47, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.W.; Xu, W.T.; Wang, X.W.; Mu, Y.; Zhao, X.F.; Yu, X.Q.; Wang, J.X. A novel C-type lectin with two CRD domains from Chinese shrimp Fenneropenaeus chinensis functions as a pattern recognition protein. Mol. Immunol. 2009, 46, 1626–1637. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.X.; Zhang, M.L.; Wang, X.W. C-Type Lectin Maintains the Homeostasis of Intestinal Microbiota and Mediates Biofilm Formation by Intestinal Bacteria in Shrimp. J. Immunol. 2021, 206, 1140–1150. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, D.; Wang, L.; Gai, Y.; Zhou, Z.; Zhang, H.; Song, L. The molecular characterization of a cyclophilin A from Chinese mitten crab Eriocheir sinensis and the antifungal activity of its recombinant protein. Electron. J. Biotechn. 2013, 16, 2. [Google Scholar] [CrossRef]
- Wang, X.W.; Vasta, G.R.; Wang, J.X. The functional relevance of shrimp C-type lectins in host-pathogen interactions. Dev. Comp. Immunol. 2020, 109, 103708. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.L.; Hancock, R.E. Cationic host defense (antimicrobial) peptides. Curr. Opin. Immunol. 2006, 18, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.; Brown, K.L.; Mookherjee, N. Host defence peptides from invertebrates-emerging antimicrobial strategies. Immunobiology 2006, 211, 315–322. [Google Scholar] [CrossRef]
- Tassanakajon, A.; Amparyup, P.; Somboonwiwat, K.; Supungul, P. Cationic antimicrobial peptides in penaeid shrimp. Mar. Biotechnol. 2011, 13, 639–657. [Google Scholar] [CrossRef] [PubMed]
- Destoumieux, D.; Munoz, M.; Bulet, P.; Bachere, E. Penaeidins, a family of antimicrobial peptides from penaeid shrimp (Crustacea, Decapoda). Cell. Mol. Life. Sci. 2000, 57, 1260–1271. [Google Scholar] [CrossRef]
- Liu, B.; Ge, X.; He, Y.; Xie, J.; Xu, P.; He, Y.; Zhaou, Q.; Pan, L.; Chen, R. Effects of anthraquinones extracted from Rheum officinale Bail on the growth, non-specific immune response of Macrobrachium rosenbergii. Aquaculture 2010, 310, 13–19. [Google Scholar] [CrossRef]
- Romero, J.; Feijoó, C.G.; Navarrete, P. Antibiotics in aquaculture-use, abuse and alternatives. HEV. Aquacult. 2012, 159, 159–198. [Google Scholar]
- Lee, I.M.; Hammond, R.W.; Davis, R.E.; Gundersen, D.E. Universal amplification and analysis of pathogen 16S rDNA for classification and identification of mycoplasmalike organisms. Phytopathology 1993, 83, 834–842. [Google Scholar] [CrossRef]
- Lalitha, M.K. Manual on antimicrobial susceptibility testing. In Performance Standards for Antimicrobial Testing: Twelfth Informational Supplement; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2004; Volume 56238, pp. 454–456. [Google Scholar]
- Wayne, P.A. Clinical and laboratory standards institute. Performance standards for antimicrobial susceptibility testing. Inf. Suppl. 2011, 31, 100–121. [Google Scholar]
- Kärber, G. Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Arch. Exptl. Pathol. Pharmakol. 1931, 162, 480–483. [Google Scholar] [CrossRef]
- Pan, L.; Yang, Y.; Peng, Y.; Li, D.; Khan, T.A.; Chen, P.; Yan, P.; Hu, S.B.; Ding, X.Z.; Sun, Y.J.; et al. The novel pathogenic Citrobacter freundii (CFC202) isolated from diseased crucian carp (Carassius auratus) and its ghost vaccine as a new prophylactic strategy against infection. Aquaculture 2021, 533, 736190. [Google Scholar] [CrossRef]
- Chen, M.; Chen, X.Q.; Tian, L.X.; Liu, Y.J.; Niu, J. Improvement of growth, intestinal short-chain fatty acids, non-specific immunity and ammonia resistance in Pacific white shrimp (Litopenaeus vannamei) fed dietary water-soluble chitosan and mixed probiotics. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2020, 236, 108791. [Google Scholar] [CrossRef]
- Frederiksen, W. Citrobacter. In Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley: Hoboken, NJ, USA, 2015; pp. 1–23. [Google Scholar]
- Baldissera, M.D.; Souza, C.F.; Junior, G.B.; Moreira, K.L.S.; da Veiga, M.L.; da Rocha, M.; Baldisserotto, B. Citrobacter freundii impairs the phosphoryl transfer network in the gills of Rhamdia quelen: Impairment of bioenergetics homeostasis. Microb. Pathog. 2018, 117, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Lü, A.; Hu, X.; Xue, J.; Zhu, J.; Wang, Y.; Zhou, G. Gene expression profiling in the skin of zebrafish infected with Citrobacter freundii. Fish. Shellfish. Immunol. 2012, 32, 273–283. [Google Scholar] [CrossRef]
- Sun, H.Y.; Cao, X.H.; Jiang, Y.F.; Ni, L.Y.; Mo, Z.Q.; Qin, Q.W.; Li, Y.W.; Dan, X.M. Outbreak of a novel disease associated with Citrobacter freundii infection in freshwater cultured stingray, Potamotrygon motoro. Aquaculture 2018, 492, 35–39. [Google Scholar] [CrossRef]
- Huang, X.D.; Zhou, H.H.; An, J.; Yang, X.L.; Cao, H.P. Isolation, identification and antibiotic susceptibility of pathogenic Citrobacter freundii from Eriocheir sinensis. J. South. Agric. 2019, 50, 1613–1619. [Google Scholar]
- Xiong, F.; Qin, L.; Hao, Y.T.; Zhao, D.; Li, W.X.; Zou, H.; Li, M.; Wu, S.G.; Wang, G.T. Gut microbiota modulation and immunity response induced by Citrobacter freundii strain GC01 in grass carp (Ctenopharyngodon idellus). Aquaculture 2020, 521, 735015. [Google Scholar] [CrossRef]
- Mauel, M.J.; Miller, D.L.; Frazier, K.S.; Hines, M.E. Bacterial pathogens isolated from cultured bullfrogs (Rana Castesbeiana). J. Vet. Diag. Inv. 2002, 14, 431–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novak, S.S.; Seigel, R.A. Gram-negative septicemia in American alligators (Alligator mississippiensis). J. Wildl. Dis. 1986, 22, 484–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bover-Cid, S.; Holzapfel, W.H. Improved screening procedure for biogenic amine production by lactic acid bacteria. Int. J. Food. Microbiol. 1999, 53, 33–41. [Google Scholar] [CrossRef]
- Chuang, Y.M.; Tseng, S.P.; Teng, L.J.; Ho, Y.C.; Hsueh, P.R. Emergence of cefotaxime resistance in Citrobacter freundii causing necrotizing fasciitis and osteomyelitis. J. Inf. 2006, 53, e161–e163. [Google Scholar] [CrossRef] [PubMed]
- Brock, T.D. Robert Koch: A Life in Medicine and Bacteriology; Zondervan: Grand Rapids, MI, USA, 1999. [Google Scholar]
- Lü, A.; Hu, X.; Zheng, L.; Zhu, A.; Cao, C.; Jiang, J. Isolation and characterization of Citrobacter spp. from the intestine of grass carp Ctenopharyngodon idellus. Aquaculture 2011, 313, 156–160. [Google Scholar] [CrossRef]
- Kumar, P.; Meghvansi, M.K.; Kamboj, D.V. Phenotypic characterization and whole-genome analysis of a novel bacteriophage HCF1 infecting Citrobacter amalonaticus and C. freundii. Front. Microbiol. 2021, 12, 15. [Google Scholar] [CrossRef] [PubMed]
- Tajima, M.; Takenouchi, Y.; Sugawara, S.; Inoue, M.; Mitsuhashi, S. Purification and properties of chromosomally mediated β-lactamase from Citrobacter freundii GN7391. Microbiology 1980, 121, 449–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, M.V.A.D.; Cangussú, Í.M.; Carvalho, A.L.S.D.; Brito, I.L.P.; Costa, R.A. Drug resistance, AmpC-β-lactamase and extended-spectrum β-lactamase-producing Enterobacteriaceae isolated from fish and shrimp. Rev. Inst. Med. Trop. 2017, 59, e70. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.M.; Huang, M.; Wu, H.J.; Guo, M.K.; Wu, L.T. Identification of CFE-2, a new plasmid-encoded AmpC β-lactamase from a clinical isolate of Citrobacter freundii. Int. J. Antimicrob. Ag. 2018, 52, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.S.; Sultana, M.; Hossain, M.A. Complete genome arrangement revealed the emergence of a poultry origin superbug Citrobacter portucalensis strain NR-12. J. Glob. Antimicrob. Resist. 2019, 18, 126–129. [Google Scholar] [CrossRef] [PubMed]
- Toranzo, A.E.; Cutrin, J.M.; Roberson, B.S.; Nunez, S.; Abell, J.M.; Hetrick, F.M.; Baya, A.M. Comparison of the taxonomy, serology, drug resistance transfer, and virulence of Citrobacter freundii strains from mammals and poikilothermic hosts. App. Eenviron. Microb. 1994, 60, 1789–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duman, M.; Saticioglu, I.B.; Buyukekiz, A.G.; Balta, F.; Altun, S. Molecular characterization and antimicrobial resistance profile of atypical Citrobacter gillenii and Citrobacter sp. isolated from diseased rainbow trout (Oncorhynchus mykiss). J. Glob. Antimicrob. Resist. 2017, 10, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Ture, M.; Kutlu, I. Isolation of Citrobacter freundii from Rainbow Trout (Oncorhynchus mykiss) in Freshwater Cage. J. Limnol. Freshw. Fish. Res. 2018, 4, 85–89. [Google Scholar] [CrossRef]
- Zhao, C.Y.; Fu, H.T.; Sun, S.M.; Qiao, H.; Zhang, W.Y.; Jin, S.B.; Gong, Y.S. Experimental inoculation of oriental river prawn Macrobrachium nipponense with white spot syndrome virus (WSSV). Dis. Aquat. Organ. 2017, 126, 125–134. [Google Scholar] [CrossRef]
- Jiravanichpaisal, P.; Roos, S.; Edsman, L.; Liu, H.; Söderhäll, K. A highly virulent pathogen, Aeromonas hydrophila, from the freshwater crayfish Pacifastacus leniusculus. J. Invertebr. Pathol. 2009, 101, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Khimmakthong, U.; Sukkarun, P. The spread of Vibrio parahaemolyticus in tissues of the Pacific white shrimp Litopenaeus vannamei analyzed by PCR and histopathology. Microb. Pathog. 2017, 113, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Jiang, Z.; Ao, S.; Gao, X.; Zhu, X.; Zhang, Z.; Zhang, X. Multi-biomarker assessment in the giant freshwater prawn Macrobrachium rosenbergii after deltamethrin exposure. Ecotoxicol. Environ. Saf. 2021, 214, 112067. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zhang, J.; Dong, H.; Wang, Y.; Liu, Q.; Li, H. Oxidative stress response of the black tiger shrimp Penaeus monodon to Vibrio parahaemolyticus challenge. Fish. Shellfish. Immunol. 2015, 46, 354–365. [Google Scholar] [CrossRef] [PubMed]
- Li, C.C.; Yeh, S.T.; Chen, J.C. The immune response of white shrimp Litopenaeus vannamei following Vibrio alginolyticus injection. Fish. Shellfish. Immunol. 2008, 25, 853–860. [Google Scholar] [CrossRef]
- Neves, C.A.; Santos, E.A.; Bainy, A.C.D. Reduced superoxide dismutase activity in Palaemonetes argentinus (Decapoda, Palemonidae) infected by Probopyrus ringueleti (Isopoda, Bopyridae). Dis. Aquat. Organ. 2000, 39, 155–158. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.; Tung, Y.H.; Liu, C.H.; Chen, J.C. Molecular cloning and characterisation of copper/zinc superoxide dismutase (Cu, Zn-SOD) from the giant freshwater prawn Macrobrachium rosenbergii. Fish. Shellfish. Immunol. 2006, 21, 102–112. [Google Scholar] [CrossRef]
- Xiu, Y.; Wu, T.; Du, J.; Yao, W.; Li, W.; Ding, Z.; Ren, Q.; Gu, W.; Meng, Q.; Wang, W. Molecular characterization and expression analysis of extracellular copper/zinc superoxide dismutase (ecCuZnSOD) from oriental river prawn, Macrobrachium nipponense. Aquaculture 2013, 380, 23–28. [Google Scholar] [CrossRef]
- Cheng, W.; Tung, Y.H.; Liu, C.H.; Chen, J.C. Molecular cloning and characterisation of cytosolic manganese superoxide dismutase (cytMn-SOD) from the giant freshwater prawn Macrobrachium rosenbergii. Fish. Shellfish. Immunol. 2006, 20, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Ni, D.; Song, L.; Gao, Q.; Wu, L.; Yu, Y.; Zhao, J.; Qiu, L.; Zhang, H.; Shi, F. The cDNA cloning and mRNA expression of cytoplasmic Cu, Zn superoxide dismutase (SOD) gene in scallop Chlamys farreri. Fish. Shellfish. Immunol. 2007, 23, 1032–1042. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Sun, H.; Chen, A.; Ning, X.; Wu, H.; Qin, S.; Xue, Q.; Zhao, J. Identification and characterization of an intracellular Cu, Zn-superoxide dismutase (icCu/Zn-SOD) gene from clam Venerupis philippinarum. Fish. Shellfish. Immunol. 2010, 28, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Zhu, H.; Ye, M.; Ma, Y. Macrobrachium rosenbergii Cu/Zn superoxide dismutase (Cu/Zn SOD) expressed in Saccharomyces cerevisiae and evaluation of the immune function to Vibrio parahaemolyticus. Fish. Shellfish. Immunol. 2019, 90, 363–375. [Google Scholar] [CrossRef]
- Mathew, S.; Kumar, K.A.; Anandan, R.; Nair, P.G.V.; Devadasan, K. Changes in tissue defence system in white spot syndrome virus (WSSV) infected Penaeus monodon. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 145, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Zhu, H.; Liu, P.; Chen, J.; Xiu, Y.; Yao, W.; Wu, T.; Ren, Q.; Meng, Q.; Gu, W.; et al. Immune responses and gene expression in hepatopancreas from Macrobrachium rosenbergii challenged by a novel pathogen spiroplasma MR-1008. Fish. Shellfish. Immunol. 2013, 34, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Ahn, I.Y.; Cheon, J.; Park, H. Molecular cloning and thermal stress-induced expression of a pi-class glutathione S-transferase (GST) in the Antarctic bivalve Laternula elliptica. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2009, 152, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Ren, Q.; Sun, R.R.; Zhao, X.F.; Wang, J.X. A selenium-dependent glutathione peroxidase (Se-GPx) and two glutathione S-transferases (GSTs) from Chinese shrimp (Fenneropenaeus chinensis). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2009, 149, 613–623. [Google Scholar] [CrossRef]
- Liu, C.H.; Tseng, M.C.; Cheng, W. Identification and cloning of the antioxidant enzyme, glutathione peroxidase, of white shrimp, Litopenaeus vannamei, and its expression following Vibrio alginolyticus infection. Fish. Shellfish. Immunol. 2007, 23, 34–45. [Google Scholar] [CrossRef]
- Hikima, S.; Hikima, J.I.; Rojtinnakorn, J.; Hirono, I.; Aoki, T. Characterization and function of kuruma shrimp lysozyme possessing lytic activity against Vibrio species. Gene 2003, 316, 187–195. [Google Scholar] [CrossRef]
- Sotelo-Mundo, R.R.; Islas-Osuna, M.A.; De-la-Re-Vega, E.; Hernández-López, J.; Vargas-Albores, F.; Yepiz-Plascencia, G. cDNA cloning of the lysozyme of the white shrimp Penaeus vannamei. Fish. Shellfish. Immunol. 2003, 15, 325–331. [Google Scholar] [CrossRef]
- Franchini, A.; Ottaviani, E. Fine structure and acid phosphatase localization of hemocytes in the freshwater snail Viviparus ater (Gastropoda, Prosobranchia). J. Invertebr. Pathol. 1990, 55, 28–34. [Google Scholar] [CrossRef]
- Sung, H.H.; Sun, R.Y.A.N. Intrahaemocytic activity of lysosomal enzymes in Penaeus monodon and Macrobrachium rosenbergii. Fish. Shellfish. Immunol. 1999, 9, 505. [Google Scholar] [CrossRef]
- Sun, C.B.; Wang, G.; Chan, S.F. Effects of artificial infection of Litopenaeus vannamei by Micrococcus lysodeikticus and WSSV on the activity of immunity related enzymes. Fish. Shellfish. Immunol. 2015, 46, 778–786. [Google Scholar] [CrossRef]
- Wang, X.; Wang, L.; Zhang, H.; Ji, Q.; Song, L.; Qiu, L.; Zhou, Z.; Wang, M.Q.; Wang, L. Immune response and energy metabolism of Chlamys farreri under Vibrio anguillarum challenge and high temperature exposure. Fish. Shellfish. Immunol. 2012, 33, 1016–1026. [Google Scholar] [CrossRef] [PubMed]
- Pang, H.; Wang, G.; Zhou, S.; Wang, J.; Zhao, J.; Hoare, R.; Monaghan, S.J.; Wang, Z.L.; Sun, C.B. Survival and immune response of white shrimp Litopenaeus vannamei following single and concurrent infections with WSSV and Vibrio parahaemolyticus. Fish. Shellfish. Immunol. 2019, 92, 712–718. [Google Scholar] [CrossRef]
- Li, Y.; Han, Z.; Xu, W.; Li, X.; Zhao, Y.; Wei, H.; Li, X.; Chen, Q. Antioxidant and immune responses of the Oriental river prawn Macrobrachium nipponense to the isopod parasite Tachaea chinensis. Fish. Shellfish. Immunol. 2020, 101, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Lan, J.F.; Sun, J.J.; Jia, W.M.; Zhao, X.F.; Wang, J.X. A novel crustin from Marsupenaeus japonicus promotes hemocyte phagocytosis. Dev. Comp. Immunol. 2015, 49, 313–322. [Google Scholar] [CrossRef]
- Amparyup, P.; Kondo, H.; Hirono, I.; Aoki, T.; Tassanakajon, A. Molecular cloning, genomic organisation and recombinant expression of a crustin like antimicrobial peptide from black tiger shrimp Penaeus monodon. Mol. Immunol. 2008, 45, 1085–1093. [Google Scholar] [CrossRef]
- Shockey, J.E.; O’Leary, N.A.; de la Vega, E.; Browdy, C.L.; Baatz, J.E.; Gross, P.S. The role of crustins in Litopenaeus vannamei in response to infection with shrimp pathogens: An in vivo approach. Dev. Comp. Immunol. 2009, 33, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Ma, C.; Zhu, P.; Yang, Y.; Lei, A.; Chen, X.; Liang, W.; Chen, M.; Xiong, J.; Li, C. A new crustin is involved in the innate immune response of shrimp Litopenaeus vannamei. Fish. Shellfish. Immunol. 2019, 94, 398–406. [Google Scholar] [CrossRef]
- Li, S.; Guo, S.; Li, F.; Xiang, J. Characterization and function analysis of an anti-lipopolysaccharide factor (ALF) from the Chinese shrimp Fenneropenaeus chinensis. Dev. Comp. Immunol. 2014, 46, 349–355. [Google Scholar] [CrossRef] [PubMed]
- De la Vega, E.; O’Leary, N.A.; Shockey, J.E.; Robalino, J.; Payne, C.; Browdy, C.L.; Warr, G.W.; Gross, P.S. Anti-lipopolysaccharide factor in Litopenaeus vannamei (LvALF): A broad spectrum antimicrobial peptide essential for shrimp immunity against bacterial and fungal infection. Mol. Immunol. 2008, 45, 1916–1925. [Google Scholar] [CrossRef]
- Liu, Y.; Song, L.; Sun, Y.; Liu, T.; Hou, F.; Liu, X. Comparison of immune response in Pacific white shrimp, Litopenaeus vannamei, after knock down of Toll and IMD gene in vivo. Dev. Comp. Immunol. 2016, 60, 41–52. [Google Scholar] [CrossRef]
- Muhammad, F.; Zhi-Feng, Z.; Ming-Yu, S.; Shafi, M. cDNA Cloning and expression of cyclophilin A (LvCypA) in white leg shrimp, Litopenaeus vannamei. Pak. J. Zoöl. 2017, 49, 935–941. [Google Scholar] [CrossRef]
- Qiu, L.; Jiang, S.; Huang, J.; Wang, W.; Zhu, C.; Su, T. Molecular cloning and mRNA expression of cyclophilin A gene in black tiger shrimp (Penaeus monodon). Fish. Shellfish. Immunol. 2009, 26, 115–121. [Google Scholar] [CrossRef]
- Song, X.; Wang, L.; Song, L.; Zhao, J.; Zhang, H.; Zheng, P.; Qiu, L.; Liu, X.; Wu, L. A cyclophilin A inducible expressed in gonad of zhikong scallop Chlamys farreri. Mol. Biol. Rep. 2009, 36, 1637–1645. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Mu, C.; Zhao, J.; Wang, C. Molecular cloning and characterization of two isoforms of cyclophilin A gene from Venerupis philippinarum. Fish. Shellfish. Immunol. 2011, 31, 1218–1223. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Jiang, S.; Huang, J.; Zhou, F.; Yang, Q.; Jiang, S.; Yang, L. C-type lectin response to bacterial infection and ammonia nitrogen stress in tiger shrimp (Penaeus monodon). Fish. Shellfish. Immunol. 2019, 90, 188–198. [Google Scholar] [CrossRef]
- Liu, Y.C.; Li, F.H.; Dong, B.; Wang, B.; Luan, W.; Zhang, X.J.; Zhang, L.S.; Xiang, J.H. Molecular cloning, characterization and expression analysis of a putative C-type lectin (Fclectin) gene in Chinese shrimp Fenneropenaeus chinensis. Mol. Immunol. 2007, 44, 598–607. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.D.; Fu, L.D.; Jia, Y.P.; Du, X.J.; Wang, Q.; Wang, Y.H.; Zhao, X.F.; Yu, X.Q.; Wang, J.X. A hepatopancreas-specific C-type lectin from the Chinese shrimp Fenneropenaeus chinensis exhibits antimicrobial activity. Mol. Immunol. 2008, 45, 348–361. [Google Scholar] [CrossRef]
- Wang, X.W.; Zhang, X.W.; Xu, W.T.; Zhao, X.F.; Wang, J.X. A novel C-type lectin (FcLec4) facilitates the clearance of Vibrio anguillarum in vivo in Chinese white shrimp. Dev. Comp. Immunol. 2009, 33, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Xu, W.T.; Zhang, X.W.; Zhao, X.F.; Yu, X.Q.; Wang, J.X. A C-type lectin is involved in the innate immune response of Chinese white shrimp. Fish. Shellfish. Immunol. 2009, 27, 556–562. [Google Scholar] [CrossRef]
- Wei, X.; Liu, X.; Yang, J.; Fang, J.; Qiao, H.; Zhang, Y.; Yang, J. Two C-type lectins from shrimp Litopenaeus vannamei that might be involved in immune response against bacteria and virus. Fish. Shellfish. Immunol. 2012, 32, 132–140. [Google Scholar] [CrossRef]
- Li, M.; Li, C.; Ma, C.; Li, H.; Zuo, H.; Weng, S.; Chen, X.; Zeng, D.; He, J.; Xu, X. Identification of a C-type lectin with antiviral and antibacterial activity from pacific white shrimp Litopenaeus vannamei. Dev. Comp. Immunol. 2014, 46, 231–240. [Google Scholar] [CrossRef]
- Bi, J.; Ning, M.; Xie, X.; Fan, W.; Huang, Y.; Gu, W.; Wang, W.; Wang, L.; Meng, Q. A typical C-type lectin, perlucin-like protein, is involved in the innate immune defense of whiteleg shrimp Litopenaeus vannamei. Fish. Shellfish. Immunol. 2020, 103, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Xiu, Y.; Hou, L.; Liu, X.; Wang, Y.; Gu, W.; Meng, Q.; Wang, W. Isolation and characterization of two novel C-type lectins from the oriental river prawn, Macrobrachium nipponense. Fish. Shellfish. Immunol. 2015, 46, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Zhai, Q.; Wang, J.; Li, J.; Li, J. Comparison of florfenicol pharmacokinetics in Exopalaemon carinicauda at different temperatures and administration routes. J. Vet. Pharmacol. Ther. 2019, 42, 230–238. [Google Scholar] [CrossRef]
- Barani, A.; Fallah, A.A. Occurrence of tetracyclines, sulfonamides, fluoroquinolones and florfenicol in farmed rainbow trout in Iran. Food. Agr. Immunol. 2015, 26, 420–429. [Google Scholar] [CrossRef]
- Caipang, C.M.A.; Lazado, C.C.; Brinchmann, M.F.; Berg, I.; Kiron, V. In vivo modulation of immune response and antioxidant defense in Atlantic cod, Gadus morhua following oral administration of oxolinic acid and florfenicol. Comp. Biochem. Physi. C 2009, 150, 459–464. [Google Scholar] [CrossRef]
- Van de Riet, J.M.; Potter, R.A.; Christie-Fougere, M.; Burns, B.G. Simultaneous determination of residues of chloramphenicol, thiamphenicol, florfenicol, and florfenicol amine in farmed aquatic species by liquid chromatography/mass spectrometry. J. AOAC. Int. 2003, 86, 510–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, W.; Li, G.; Zhou, S.; Li, X.; Hu, L.; Zhou, J. Pharmacokinetics and Tissue Distribution of Thiamphenicol and Florfenicol in Pacific White Shrimp Litopenaeus vannameiin Freshwater following Oral Administration. J. Aqua. Anim. Health. 2013, 25, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Burridge, L.; Weis, J.S.; Cabello, F.; Pizarro, J.; Bostick, K. Chemical use in salmon aquaculture: A review of current practices and possible environmental effects. Aquaculture 2010, 306, 7–23. [Google Scholar] [CrossRef]
- Parmar, P.; Yusufzai, S.K.; Parmar, H.V.; Nanjiyani, R.P.; Chavda, V.M. Therapeutic potentiality of florfenicol against vibriosis in Litopenaeus vannamei. J. Entomol. Zool. Stud. 2018, 6, 463–467. [Google Scholar]
- Tipmongkolsilp, N.; Limpanon, Y.; Patamalai, B.; Lusanandana, P.; Wongtavatchai, J. Oral medication with florfenicol for black tiger shrimps Penaeus monodon. Thai J. Vet. Med. 2006, 36, 39–47. [Google Scholar]
- Yasunaga, N.; Yasumoto, S. Therapeutic effect of florfenicol on experimentally induced pseudotuberculosis in Yellowtail. Fish. Pathol. 1988, 23, 1–5. [Google Scholar] [CrossRef]
- Fukui, H.; Fujihara, Y.; Kano, T. In vitro and in vivo antibacterial activities of florfenicol, a new fluorinated analog of thiamphenicol, against fish pathogens. Fish. Pathol. 1987, 22, 201–207. [Google Scholar] [CrossRef]
- Inglis, V.; Richards, R.H.; Varma, K.J.; Sutherland, I.H.; Brokken, E.S. Florfenicol in Atlantic salmon, Salmo salar L., parr: Tolerance and assessment of efficacy against furunculosis. J. Fish Dis. 1991, 14, 343–351. [Google Scholar] [CrossRef]
- Van Hai, N.; Buller, N.; Fotedar, R. The use of customised probiotics in the cultivation of western king prawns (Penaeus latisulcatus Kishinouye, 1896). Fish. Shellfish. Immunol. 2009, 27, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Shivu, M.M.; Rajeeva, B.C.; Girisha, S.K.; Karunasagar, I.; Krohne, G.; Karunasagar, I. Molecular characterization of Vibrio harveyi bacteriophages isolated from aquaculture environments along the coast of India. Environ. Microbiol. 2007, 9, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Randrianarivelo, R.; Danthu, P.; Benoit, C.; Ruez, P.; Raherimandimby, M.; Sarter, S. Novel alternative to antibiotics in shrimp hatchery: Effects of the essential oil of Cinnamosma fragrans on survival and bacterial concentration of Penaeus monodon larvae. J. Appl. Microbiol. 2010, 109, 642–650. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Dry Weight (%) |
|---|---|
| Rice bran | 30.0 |
| Soybean meal (44%) | 25.0 |
| Wheat midds | 17.75 |
| Menhaden meal (72%) | 16.0 |
| Meat, bone, and blood meal | 11.0 |
| Mineral premix a | 0.1 |
| Vitamin premix b | 0.1 |
| Choline | 0.05 |
| Proximate analysis (%) | |
| Moisture | 12.3 |
| Crude protein | 36.2 |
| Crude fat c | 6.3 |
| Crude fiber | 4.4 |
| Ash | 10.7 |
| Nitrogen free extract (by difference) | 30.1 |
| Target Genes | Sequences (5′–3′) | Accessions |
|---|---|---|
| β-actin | Forward: GAGACCTTCAACACCCCCGC | AF221096.1 |
| Reverse: TAGGTGGTCTCGTGGATGCC | ||
| ALF | Forward: ATCTGGCGTCGTTACCAAAAC | JQ364961.1 |
| Reverse: GAAATGAAACCTGATGATCGTC | ||
| Crustin | Forward: AACGACTTCAAGTGCTTCGGGTCT | JQ413342.1 |
| Reverse: AAGCTTAGTGGTTTGCAGACGTGC | ||
| CTL | Forward: ATGTTGACCTTAATGGCCAC | KX495215.1 |
| Reverse: CTTTTCTGTGGGCGTTTCTTC | ||
| Cu/Zn-SOD | Forward: TCGCCTAACGAGGAGGTTC | DQ121374.1 |
| Reverse: CGGCTTCATCAGGATTTTGAG | ||
| CypA | Forward: CTAATGCTGGACCCAACACC | EL696406.1 |
| Reverse: CCTCCACT CCAAT TCTAGCTGTAA | ||
| LZM | Forward: TGCCATCAACCACCACAACT | AY257549.2 |
| Reverse: CCCCTTTCCCTTCCACTTCT | ||
| CAT | Forward: AGCGAGATTGGCAAGAAGACACC | HQ668089.1 |
| Reverse: AAGGATGGTGACCTGGTGCGTGG | ||
| GPx | Forward: TTCGCCCAGGGAACAATTT | EL696567.1 |
| Reverse: CCTTTTCACTGAGAATTACCCAG | ||
| GST | Forward: GTTGTGCAGCATTGAGGTTTAT | HF570114.1 |
| Reverse: GTATCCTACACCATGTGCTCTG | ||
| ACP | Forward: GTTTACACTCGCCTTATCCTCCG | JX975267.1 |
| Reverse: CTTTGTGCATGAACATGACCCTG |
| Characteristics | Strain GDZQ201912 | Citrobacter freundii * |
|---|---|---|
| Gram reaction | − | − |
| Shape | Rod | Rod |
| Motility | + | + |
| Indole | − | V |
| Citrate | + | V |
| Methyl red | + | + |
| H2S | + | V |
| Ornithine | − | − |
| Malonate | − | V |
| Sucrose | + | + |
| Melibiose | + | + |
| Raffinose | + | V |
| Dulcitol | + | V |
| Adonitol | − | − |
| Characteristics | Strain GDZQ201912 | Citrobacter freundii * |
|---|---|---|
| cis-Aconitate | + | + |
| trans-Aconitate | + | + |
| Adonitol | − | − |
| L-alanine | + | + |
| γ-Aminobutyrate | + | + |
| D-arabitol | − | − |
| Benzoate | − | − |
| Caprate | − | − |
| D-cellobiose | + | + |
| Dulcitol | + | + |
| D-galactose | + | + |
| N-acetyl-β-D-galactosamine | + | + |
| D-galacturonic acid | + | + |
| L-galactose | + | + |
| Esculin | − | − |
| D-fucose | + | + |
| D-glucose | + | + |
| Gentiobiose | + | + |
| Gentisate | + | + |
| L-glutamate | + | + |
| Glycerol | + | + |
| Inositol | + | + |
| L-lactate | + | + |
| Lactose | + | + |
| Lactulose | + | + |
| Maltitol | + | + |
| Melibiose | + | + |
| Phenylacetate | − | + |
| L-proline | + | + |
| Putrescine | − | + |
| Raffinose | + | + |
| D-sorbitol | + | + |
| Sucrose | + | + |
| D-turanose | − | − |
| L-tyrosine | − | + |
| Stachyose | − | − |
| D-fructose | − | − |
| L-fructose | + | + |
| D-mannose | + | + |
| L-rhamnose monohydrate | + | + |
| dextrin | − | − |
| D-glucosamic acid | + | + |
| D-salicin | + | + |
| N-acetylglucosamine | + | + |
| D-mannitol | + | + |
| N-acetylneuraminic acid | + | + |
| D-glucuronamide | − | − |
| D-serine | + | + |
| D-aspartic acid | + | + |
| L-arginine | − | − |
| Methyl pyruvate | + | + |
| Methyl D3phenyllactate | − | − |
| D-malic acid | + | + |
| L-malic acid | + | + |
| Number | Antibiotics | Drug Content (µg/Pill) | Diameter of Inhibition Zone | Results |
|---|---|---|---|---|
| 1 | penicillin | 10 | 6.00 ± 0.00 | ± |
| 2 | doxycycline | 30 | 18.36 ± 0.16 | + + |
| 3 | florfenicol | 30 | 28.57 ± 0.19 | + + + |
| 4 | norfloxacin | 10 | 28.85 ± 0.21 | + + + |
| 5 | ofloxacin | 5 | 29.91 ± 0.13 | + + + |
| 6 | amoxicillin | 20 | 14.23 ± 0.11 | + + |
| 7 | ampicillin | 10 | 6.00 ± 0.00 | ± |
| 8 | cefepime | 30 | 41.12 ± 0.23 | + + + |
| 9 | ceftriaxone | 30 | 36.78 ± 0.15 | + + + |
| 10 | neomycin | 30 | 19.63 ± 0.12 | + + + |
| 11 | gentamicin | 10 | 17.51 ± 0.18 | + + + |
| 12 | streptomycin | 10 | 10.03 ± 0.10 | + + |
| 13 | clindamycin | 2 | 6.00 ± 0.00 | ± |
| 14 | lincomycin | 2 | 6.00 ± 0.00 | ± |
| 15 | tetracycline | 30 | 26.56 ± 0.17 | + + + |
| 16 | novobiocin | 30 | 6.00 ± 0.00 | ± |
| 17 | Polymyxin B | 300 | 19.21 ± 0.14 | + + + |
| 18 | rifampin | 5 | 10.11 ± 0.12 | + + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, C.; Wen, H.; Huang, S.; Weng, S.; He, J. A Novel Disease (Water Bubble Disease) of the Giant Freshwater Prawn Macrobrachium rosenbergii Caused by Citrobacter freundii: Antibiotic Treatment and Effects on the Antioxidant Enzyme Activity and Immune Responses. Antioxidants 2022, 11, 1491. https://doi.org/10.3390/antiox11081491
Zhao C, Wen H, Huang S, Weng S, He J. A Novel Disease (Water Bubble Disease) of the Giant Freshwater Prawn Macrobrachium rosenbergii Caused by Citrobacter freundii: Antibiotic Treatment and Effects on the Antioxidant Enzyme Activity and Immune Responses. Antioxidants. 2022; 11(8):1491. https://doi.org/10.3390/antiox11081491
Chicago/Turabian StyleZhao, Caiyuan, Huagen Wen, Shengsheng Huang, Shaoping Weng, and Jianguo He. 2022. "A Novel Disease (Water Bubble Disease) of the Giant Freshwater Prawn Macrobrachium rosenbergii Caused by Citrobacter freundii: Antibiotic Treatment and Effects on the Antioxidant Enzyme Activity and Immune Responses" Antioxidants 11, no. 8: 1491. https://doi.org/10.3390/antiox11081491