
The Quality of Scutellaria baicalensis Georgi Is Effectively Affected by Lithology and Soil’s Rare Earth Elements (REEs) Concentration

 ,
,
Abstract
:1. Introduction
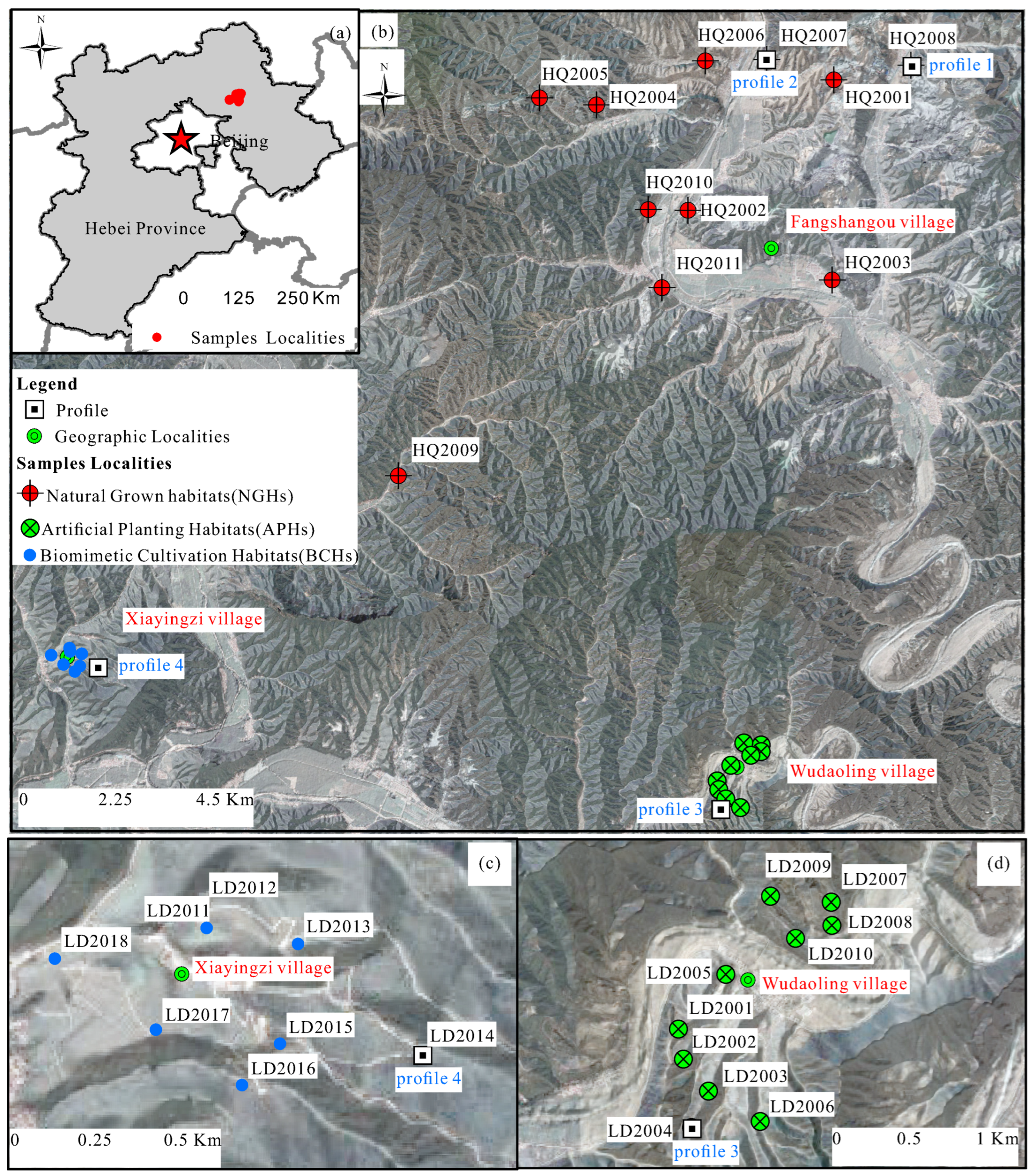
2. Physical Geography and Distribution of Scutellaria baicalensis
- (1)
- It was documented that NGHs were concentrated in southern Chengde, such as Luanping County, Xinglong County and Kuanchen County, China. Based on our field survey, NGHs in Hongqi, Luanping County, were selected for sampling (Figure 2a–c). The lithology was dominated by hornblende-gneiss, which were strongly weathered and had intensive fractures along weathering profiles, resulting in thick saprolite and soil with a thickness of 1 m.
- (2)
- Sample plots of APHs were concentrated in Wudaoling village, Luanping County, whose parent materials were loess. Scutellaria baicalensis was planted in artificial terraced fields without fertilization (Figure 2d,e).
- (3)
- Sample plots of BCHs were chosen in a Chinese herbal medicine plantation in Xiayingzi Villiage, Luanping County, whose parent materials were also loess, mingled with a small amount of weathering residues of andesitic volcanic and pyroclastic rocks. Scutellaria baicalensis was planted along turnover hillside fields under field management by biomimetic cultivation and adding a certain amount of REE-rich micro-fertilizers to replenish the soil nutrient contents (Figure 2f).
3. Materials and Methods
3.1. Sampling Strategy
3.2. Analytical Methods
3.3. Parameters on the REE Distribution Characteristics
4. Results
4.1. REE Concentrations in the Weathering Crust
4.1.1. REE Concentrations of Rhizosphere Soils
4.1.2. REE Concentrations in the Weathering Profile
4.2. REE Concentrations in the Roots of Scutellaria baicalensis
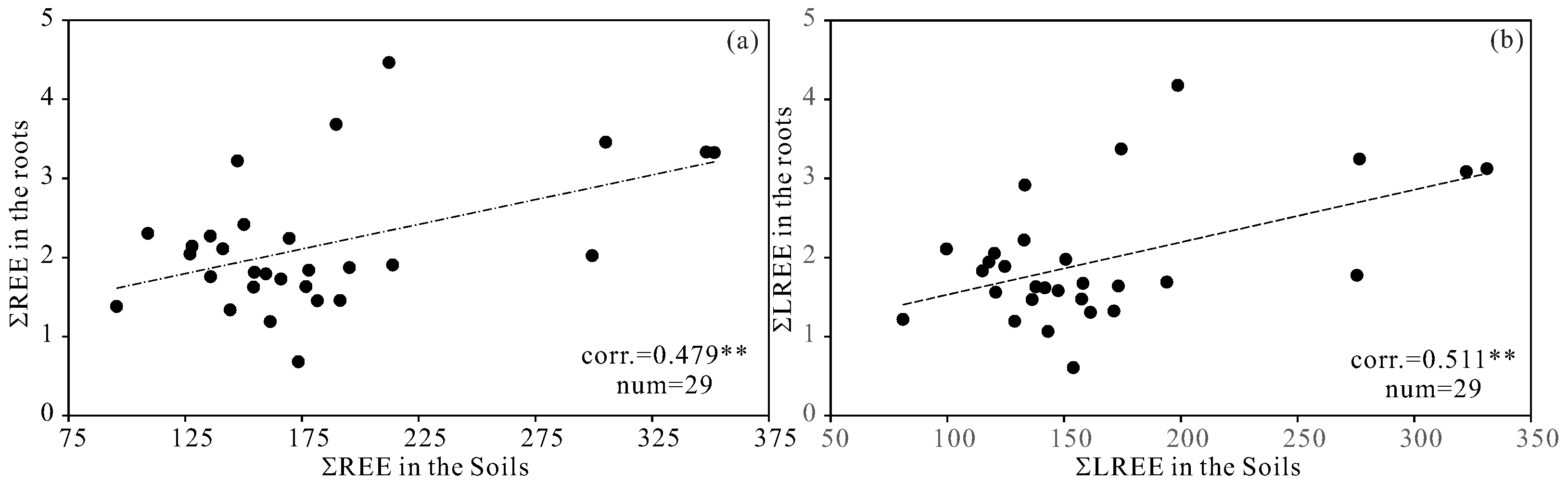
4.3. Accumulation of REE in the Roots of Scutellaria baicalensis
4.4. Effective Constituent Content of Scutellaria baicalensis
5. Discussions
5.1. Lithological Influences on the Enrichments and Fractionations of REEs in the Soils
5.2. Enrichments and Fractionations of REE in the Roots of Scutellaria baicalensis
5.3. Relationships between REE and Effective Constituents of Scutellaria baicalensis
- (1)
- Hornblende-gneiss locations were confirmed to be the natural top-geoherbs habitats of Rehe Scutellaria baicalensis for their relatively higher REEs and other micronutrient elements concentrations and soil property. In order to protect the chemical type of top-geoherbs, it was significant to avoid excessive digging of Scutellaria baicalensis to maintain their natural propagation and replacement.
- (2)
- Luanping County and Kuancheng Kuancheng Manchu Nationality Autonomous County was also the main gneiss location in the Luanhe watershed in Chengde City. It can be considered for the large-scale cultivation of Scutellaria baicalensis by biomimetic cultivation.
- (3)
- Scientific and reasonable rare earth micro-fertilizers application was optional for optimizing replanting patterns. The conventional principle that high micronutrient concentrations inhibit their absorption while low micronutrient concentrations favor their absorption should not be neglected.
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hu, S. Geoherbs in China; Heilongjiang Science and Technology Press: Haerbin, China, 1989. [Google Scholar]
- Xie, W. Discussion about geoherbs. J. Tradit. Chin. Med. 1990, 40, 43–46. [Google Scholar]
- Huang, L.; Zhang, R. Biological essential about geoherbs. Chin. Pharm. J. 1997, 32, 563–566. [Google Scholar]
- Huang, L.; Guo, L.; Ma, C.; Gao, W.; Yuan, Q. Top-geoherbs of traditional Chinese medicine: Common traits, quality characteristics and formation. Front. Med. 2011, 5, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.G.; Duan, G.L.; Chen, B.D.; Peng, X.H.; Chen, Z.; Sun, G.X. Minerals Weathering and Elemental cycling in the System of Siols-microorganisms-plants. Sci. Sin. 2014, 44, 1107–1116. [Google Scholar]
- Reimann, C.; Englmaier, P.; Fabian, K.; Gough, L.; Lamothe, P.; Smith, D. Biogeochemical plant–soil interaction: Variable element composition in leaves of four plant species collected along a south–north transect at the southern tip of Norway. Sci. Total Environ. 2015, 506, 480–495. [Google Scholar] [CrossRef]
- Yan, H.; Duan, J.A.; Qian, D.W.; Xiu, S.L.; Song, B.S.; He, Z.Q. Correlation Analysis and Evaluation of Inorganic Elements in Angelica sinensis and Its Correspondence Soil from Different Regions. J. Chin. Med. Mater. 2011, 34, 512–516. [Google Scholar]
- Zhong, X.J.; Tan, Y.F. Research progress of soil factor influences on the quality of genuine medicinal materials. J. South. Agric. 2012, 43, 1708–1711. [Google Scholar]
- Yao, Q.H.; Yan, S.A.; Zhang, B.L.; Su, H.G.; Pan, H.Y.; Lin, Q. Effects of Soil Types in Tea Garden on Distribution and Composition of Rare Earth Elements in Tieguanyin. J. Trop. Subtrop. Bot. 2018, 26, 644–650. [Google Scholar]
- Tao, Y.; Shen, L.; Feng, C.; Qu, J.; Ju, H.; Yang, R.; Zhang, Y. Distribution of rare earth elements (REEs) and their roles in plants growth: A review. Environ. Pollut. 2022, 298, 118540. [Google Scholar] [CrossRef]
- Qi, J.S.; Xu, H.B.; Zhou, J.Y.; Lu, X.H.; Yang, X.L.; Guan, J.H. Studies on the Amount of trace elements and Four Characteristics in Prescriptions of Chinese Medicine. J. Anal. Sci. 1998, 14, 283–287. [Google Scholar]
- Zhang, Z.J.; Song, B. Effect Analyses on Micronutrient or REE Fertilization to Ginseng. Liaoning Agric. Sci. 1998, 1, 14–16. [Google Scholar]
- Liu, C.E.; Ren, S.Y.; Yin, W.Y.; He, M.Y. Application Effect and Technique analyses of REE to the Eucommia ulmoides. Hunan Agric. Sci. 1999, 27, 29–31. [Google Scholar]
- Chen, H.; Liu, H.L.; Dong, Y.Y. Study on the Relationship between Rare Earth Elements (REES)and Traditional Chinese Medicine(TCM). Guangdong Trace Elem. Sci. 2001, 8, 1–8. [Google Scholar]
- Fu, W.; Li, X.; Feng, Y.; Feng, M.; Peng, Z.; Yu, H.; Lin, H. Chemical weathering of S-type granite and formation of rare earth element (REE)-rich regolith in South China: Critical control of lithology. Chem. Geol. 2019, 520, 33–51. [Google Scholar] [CrossRef]
- Brantley, S.L.; Goldhaber, M.B.; Ragnarsdottir, K.V. Crossing disciplines and scales to understand the critical zone. Elements 2007, 3, 307–314. [Google Scholar] [CrossRef]
- Hewawasam, T.; von Blanckenburg, F.; Bouchez, J.; Dixon, J.L.; Schuessler, J.A.; Maekeler, R. Slow advance of the weathering front during deep, supply-limited saprolite formation in the tropical Highlands of Sri Lanka. Geochim. Cosmochim. Acta 2013, 118, 202–230. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.X.; Pei, M.; Zhao, C.D.; Hna, W.; Wang, H.Y.; Wang, Q.L.; Yang, F.; Zhang, F.G.; Wang, C.W.; Liu, F. Epigenetic geochemical dynamics and driving mechanisms of distribution patterns of chemical elements in soil, Southwest China. Earth Sci. Front. 2019, 26, 159–191. [Google Scholar]
- Chinese Pharmacopoeia Commission. The Pharmacopoeia of the People’s Republic of China; China Medical Science Press: Beijing, China, 2015. [Google Scholar]
- Xie, Y.; Bi, J.H.; Peng, G.Y. Comparative Study on Detoxication between Dao-di Herb and Non Dao-di Herb of Huangqin. Guid. J. Tradit. Chin. Med. Pharm. 2015, 21, 35–38. [Google Scholar]
- Liu, Y.; Li, L.T.; Ji, X.Q.; Kong, L.J.; Gao, Z.H.; Pang, J.; Chen, H.N.; Wang, S.M. Effect of inorganic element in soil on contents of inorganic elements and baicalin in Scutellaria baicalensis from different regions. Chin. Tradit. Herb. Drugs 2017, 48, 1225–1228. [Google Scholar]
- Xin, X.J.; Li, Y.C.; Liu, G.J. Scutellaria baicalensis Georgi Conformation Simulation Cultivation Technology in Daxing’An Mountains. For. Sci. Technol. Inf. 2016, 48, 20–21. [Google Scholar]
- Liu, Y.Y.; Wu, P.P.; Yang, L.B.; Wang, L.M. Analysis of the Climate Characteridtics of thunderstorms in Chengde from 1973 to 2012. Meteorol. J. Inn. Mong. 2018, 2, 15–17. [Google Scholar]
- Chen, D.; Chen, G. Practical Geochemistry of Rare Earth Elements; Geological Press: Beijing, China, 1990. [Google Scholar]
- Yuan, M.; Guo, M.N.; Liu, W.S.; Liu, C.; van der Ent, A.; Morel, J.L.; Huot, H.; Zhao, W.Y.; Wei, X.G.; Qiu, R.L.; et al. The accumulation and fractionation of Rare Earth Elements in hydroponically grown Phytolacca americana L. Plant Soil 2017, 421, 67–82. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, Z.H.; Geng, X.; He, F.P.; Zu, Y.C.; Wang, L. REE Biogeochemistry of Soil-Vegetation System in Dabaoshan Polymetallic Mine. Earth Sci. J. China Univ. Geosci. 2014, 39, 733–740. [Google Scholar]
- Miao, L.; Xu, R.; Ma, Y.; Zhu, Z.; Wang, J.; Cai, R.; Chen, Y. Geochemistry and biogeochemistry of rare earth elements in a surface environment (soil and plant) in South China. Environ. Geol. 2008, 56, 225–235. [Google Scholar] [CrossRef]
- Ryan, S.E.; Snoeck, C.; Crowley, Q.G.; Babechuk, M.G. 87Sr/86Sr and trace element mapping of geosphere-hydrosphere-biosphere interactions: A case study in Ireland. Appl. Geochem. 2018, 92, 209–224. [Google Scholar] [CrossRef]
- Chen, D.; Zou, Z.; Ren, D. Preliminary application of plant exploration in search for Thallium mineral deposits. Bull. Mineral. Petrol. Geochem. 2000, 19, 397–400. [Google Scholar]
- Hu, R.; Beguiristain, T.; De Junet, A.; Leyval, C. Bioavailability and transfer of elevated Sm concentration to alfalfa in spiked soils. Environ. Sci. Pollut. Res. 2020, 27, 44333–44341. [Google Scholar] [CrossRef]
- Huang, L. Molecular Pharmacognosy; Peking University Medical Press: Beijing, China, 2006. [Google Scholar]
- Xiao, Y.Q.; Jiang, Y.; Li, L.; Zhang, W.T.; Li, M.P.; Zhang, W.S. Relationship between active components in Scutellaria baicalensis and environmental factors in mountainous region of Western Beijing. Chin. Tradit. Herb. Drugs 2009, 40, 1291–1296. [Google Scholar]
- Hu, Z.; Haneklaus, S.; Sparovek, G.; Schnug, E. Rare earth elements in soils. Commun. Soil Sci. Plant Anal. 2006, 37, 1381–1420. [Google Scholar] [CrossRef]
- Loell, M.; Reiher, W.; Felix-Henningsen, P. Contents and bioavailability of rare earth elements in agricultural soils in Hesse (Germany). J. Plant Nutr. Soil Sci. 2011, 174, 644–654. [Google Scholar] [CrossRef]
- Pye, K. Mineralogical and textural controls on the weathering of granitoid rocks. Catena 1986, 13, 47–57. [Google Scholar] [CrossRef]
- Worthington, S.R.; Davies, G.J.; Alexander, E.C., Jr. Enhancement of bedrock permeability by weathering. Earth-Sci. Rev. 2016, 160, 188–202. [Google Scholar] [CrossRef]
- Zhang, L.; Li, X.; Li, D.; Han, Z.; Zhang, G. Rare earth elements distribution and its correlation with microelements and particle-size of basalt-deriver soils in Leizhou Peninsula. Acta Pedol. Sin. 2011, 48, 1–9. [Google Scholar]
- Brioschi, L.; Steinmann, M.; Lucot, E.; Pierret, M.C.; Stille, P.; Prunier, J.; Badot, P.M. Transfer of rare earth elements (REE) from natural soil to plant systems: Implications for the environmental availability of anthropogenic REE. Plant Soil 2013, 366, 143–163. [Google Scholar] [CrossRef]
- Braun, J.J.; Riotte, J.; Battacharya, S.; Violette, A.; Prunier, J.; Bouvier, V.; Candaudap, F.; Maréchal, J.C.; Ruiz, L.; Panda, S.R.; et al. REY-Th-U solute dynamics in the critical zone: Combined influence of chemical weathering, atmospheric deposit leaching, and vegetation cycling (Mule Hole Watershed, South India). Geochem. Geophys. Geosys. 2017, 18, 4409–4425. [Google Scholar] [CrossRef]
- Chang, C.; Song, C.; Beckford, H.O.; Wang, S.; Ji, H. Behaviors of REEs during pedogenetic processes in the karst areas of Southwest China. J. Asian Earth Sci. 2019, 185, 104023. [Google Scholar] [CrossRef]
- Holdren, G.R., Jr.; Berner, R.A. Mechanism of feldspar weathering—I. Experimental studies. Geochim. Cosmochim. Acta 1979, 43, 1161–1171. [Google Scholar] [CrossRef]
- Heckman, K.; Rasmussen, C. Lithologic controls on regolith weathering and mass flux in forested ecosystems of the southwestern USA. Geoderma 2011, 164, 99–111. [Google Scholar] [CrossRef]
- Fu, S.Z.; Yan, C.L.; Wu, S.Q.; Yang, X.K. Content and Distribution Characteristics of Rare Earth Elements In Typical Soils Of Guizhou Province. Acta Pedol. Sin. 2000, 37, 109–115. [Google Scholar]
- Miao, L.; Xu, R.S.; Xu, J.H. Geochemical Characteristics of Rare Earth Elements(REEs) in the Soil-Plant System in West Guangdong Province. Acta Pedol. Sin. 2007, 44, 54–62. [Google Scholar]
- Mihajlovic, J.; Bauriegel, A.; Stärk, H.J.; Roßkopf, N.; Zeitz, J.; Milbert, G.; Rinklebe, J. Rare earth elements in soil profiles of various ecosystems across Germany. Appl. Geochem. 2019, 102, 197–217. [Google Scholar] [CrossRef]
- Coppin, F.; Berger, G.; Bauer, A.; Castet, S.; Loubet, M. Sorption of lanthanides on smectite and kaolinite. Chem. Geol. 2002, 182, 57–68. [Google Scholar] [CrossRef]
- Tertre, E.; Berger, G.; Simoni, E.; Castet, S.; Giffaut, E.; Loubet, M.; Catalette, H. Europium retention onto clay minerals from 25 to 150 C: Experimental measurements, spectroscopic features and sorption modelling. Geochim. Cosmochim. Acta 2006, 70, 4563–4578. [Google Scholar] [CrossRef] [Green Version]
- Galán, E.; Fernández-Caliani, J.; Miras, A.; Aparicio, P.; Márquez, M. Residence and fractionation of rare earth elements during kaolinization of alkaline peraluminous granites in NW Spain. Clay Miner. 2007, 42, 341–352. [Google Scholar] [CrossRef]
- Borst, A.M.; Smith, M.P.; Finch, A.A.; Estrade, G.; Villanova-de Benavent, C.; Nason, P.; Marquis, E.; Horsburgh, N.J.; Goodenough, K.M.; Xu, C.; et al. Adsorption of rare earth elements in regolith-hosted clay deposits. Nat. Commun. 2020, 11, 4386. [Google Scholar] [CrossRef]
- Andrade, G.R.P.; Cuadros, J.; Barbosa, J.M.P.; Vidal-Torrado, P. Clay minerals control rare earth elements (REE) fractionation in Brazilian mangrove soils. Catena 2022, 209, 105855. [Google Scholar] [CrossRef]
- Land, M.; Òhlander, B.; Ingri, J.; Thunberg, J. Solid speciation and fractionation of rare earth elements in a spodosol profile from northern Sweden as revealed by sequential extraction. Chem. Geol. 1999, 160, 121–138. [Google Scholar] [CrossRef]
- Laveuf, C.; Cornu, S. A review on the potentiality of rare earth elements to trace pedogenetic processes. Geoderma 2009, 154, 1–12. [Google Scholar] [CrossRef]
- Steinmann, M.; Stille, P. Rare earth element behavior and Pb, Sr, Nd isotope systematics in a heavy metal contaminated soil. Appl. Geochem. 1997, 12, 607–623. [Google Scholar] [CrossRef]
- Granados-Correa, F.; Vilchis-Granados, J.; Jiménez-Reyes, M.; Quiroz-Granados, L. Adsorption behaviour of La (III) and Eu (III) ions from aqueous solutions by hydroxyapatite: Kinetic, isotherm, and thermodynamic studies. J. Chem. 2013, 2013, 751696. [Google Scholar] [CrossRef] [Green Version]
- Yusoff, Z.M.; Ngwenya, B.T.; Parsons, I. Mobility and fractionation of REEs during deep weathering of geochemically contrasting granites in a tropical setting, Malaysia. Chem. Geol. 2013, 349, 71–86. [Google Scholar] [CrossRef]
- Yang, M.; Liang, X.; Ma, L.; Huang, J.; He, H.; Zhu, J. Adsorption of REEs on kaolinite and halloysite: A link to the REE distribution on clays in the weathering crust of granite. Chem. Geol. 2019, 525, 210–217. [Google Scholar] [CrossRef]
- Jin, P.P.; Ou, C.H.; Ma, Z.G.; Li, D.; Ren, Y.J.; Zhao, Y.F. Evolution of montmorillonite and its related clay minerals and their effects on shale gas development. Geophys. Prospect. Pet. 2018, 57, 344–355. [Google Scholar]
- Alshameri, A.; He, H.; Xin, C.; Zhu, J.; Wei, X.; Zhu, R.; Wang, H. Understanding the role of natural clay minerals as effective adsorbents and alternative source of rare earth elements: Adsorption operative parameters. Hydrometallurgy 2019, 185, 149–161. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, C.; Xiong, D.; Zhou, Y.; Sun, X. Oxide geochemical characteristics and paleoclimate records of “Wushan loess”. Geol. China 2013, 40, 352–360. [Google Scholar]
- Cui, Y.; Luo, C.G.; Xu, L.; Zhang, H.; Deng, M.G.; Gu, H.N.; Meng, Y.; Qin, C.J.; Wen, H.J. Weathering Origin and Enrichment of Lithium in Clay Rocks of the Jiujialu Formation, Central Guizhou Province, Southwest China. Bull. Mineral. Geochem. 2018, 37, 696–704. [Google Scholar]
- Quinn, K.A.; Byrne, R.H.; Schijf, J. Sorption of yttrium and rare earth elements by amorphous ferric hydroxide: Influence of pH and ionic strength. Mar. Chem. 2006, 99, 128–150. [Google Scholar] [CrossRef]
- Huang, J.; Tan, W.; Liang, X.; He, H.; Ma, L.; Bao, Z.; Zhu, J. REE fractionation controlled by REE speciation during formation of the Renju regolith-hosted REE deposits in Guangdong Province, South China. Ore Geol. Rev. 2021, 134, 104172. [Google Scholar] [CrossRef]
- Lee, Y.; Park, E.; Kim, W.; Choi, Y.; Park, J. A case of pelvic paragonimiasis combined with myoma uteri and pelvic inflammatory disease. Korean J. Parasitol. 1993, 31, 295–297. [Google Scholar] [CrossRef]
- Takahashi, Y.; Minai, Y.; Ambe, S.; Makide, Y.; Ambe, F.; Tominaga, T. Simultaneous determination of stability constants of humate complexes with various metal ions using multitracer technique. Sci. Total Environ. 1997, 198, 61–71. [Google Scholar] [CrossRef]
- Luo, Y.R.; Byrne, R.H. Carbonate complexation of yttrium and the rare earth elements in natural waters. Geochim. Cosmochim. Acta 2004, 68, 691–699. [Google Scholar] [CrossRef]
- Sonke, J.E.; Salters, V.J. Lanthanide–humic substances complexation. I. Experimental evidence for a lanthanide contraction effect. Geochim. Cosmochim. Acta 2006, 70, 1495–1506. [Google Scholar] [CrossRef]
- Zhao, Z.Z.; Xu, D.R.; Bi, H.; Tang, S.X. Vertical Distribution Pattern of REE Contents in Latosol in the Eastern Areas of Hainan Island. Geotecton. Metallog. 2006, 30, 401–407. [Google Scholar]
- Pourret, O.; Davranche, M.; Gruau, G.; Dia, A. Competition between humic acid and carbonates for rare earth elements complexation. J. Colloid Interface Sci. 2007, 305, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Liu, C. The effect of humic acid on the adsorption of REEs on kaolin. Colloids Surf. A Physicochem. Eng. Asp. 2006, 290, 112–117. [Google Scholar] [CrossRef]
- Li, Z. Study on Herbal Textual Research and Distribution and Change of Authentic Region of Huang Qin. Ph.D. Thesis, China Academy of Chinese Medical Sciences, Beijing, China, 2010. [Google Scholar]
- Wyttenbach, A.; Furrer, V.; Schleppi, P.; Tobler, L. Rare earth elements in soil and in soil-grown plants. Plant Soil 1998, 199, 267–273. [Google Scholar] [CrossRef]
- Ozaki, T.; Enomoto, S. Uptake of rare earth elements by Dryopteris erythrosora (autumn fern). Riken Rev. 2001, 1, 84–87. [Google Scholar]
- Cao, X.; Ding, Z.; Hu, X.; Wang, X. Effects of soil pH value on the bioavailability and fractionation of rare earth elements in wheat seedling (Triticum aestivum L.). Huanjing Kexue 2002, 23, 97–102. [Google Scholar]
- Tyler, G. Rare earth elements in soil and plant systems-A review. Plant Soil 2004, 267, 191–206. [Google Scholar] [CrossRef]
- Semhi, K.; Chaudhuri, S.; Clauer, N. Fractionation of rare-earth elements in plants during experimental growth in varied clay substrates. Appl. Geochem. 2009, 24, 447–453. [Google Scholar] [CrossRef]
- Gao, A.G.; Chen, Z.H.; Liu, Y.G.; Sun, H.Q.; Yang, S.Y. REE geochemical characteristics of surficial sediments in chukchi sea. Sci. China Ser. D 2003, 33, 148–154. [Google Scholar]
- Jones, D. Trivalent metal (Cr, Y, Rh, La, Pr, Gd) sorption in two acid soils and its consequences for bioremediation. Eur. J. Soil Sci. 1997, 48, 697–702. [Google Scholar] [CrossRef]
- Iyengar, S.; Martens, D.; Miller, W. Distribution and plant availability of soil zinc fractions. Soil Sci. Soc. Am. J. 1981, 45, 735–739. [Google Scholar] [CrossRef]
- LeClaire, J.P.; Chang, A.; Levesque, C.; Sposito, G. Trace Metal Chemistry in Arid-Zone Field Soils Amended with Sewage Sludge: IV. Correlations between Zinc Uptake and Extracted Soil Zinc Fractions. Soil Sci. Soc. Am. J. 1984, 48, 509–513. [Google Scholar] [CrossRef]
- Sims, J.T. Soil pH effects on the distribution and plant availability of manganese, copper, and zinc. Soil Sci. Soc. Am. J. 1986, 50, 367–373. [Google Scholar] [CrossRef]
- Zhu, W.H.; Yang, Y.G.; Bi, H.; Liu, Q. Progress in Geochemical Research of Rare Earth Element in Soils. Bull. Mineral. Petrol. Geochem. 2003, 23, 259–264. [Google Scholar]
- Ji, H.B.; Wang, L.J.; Dong, Y.S.; Wang, S.J.; Luo, J.M.; Sun, Y.Y. An Overview on the Study Of Biogeochemical Cycle for Rare Earth Elements(REEs). Prog. Geogr. 2004, 23, 51–61. [Google Scholar]
- Pang, X.; Xing, X.Y.; Wang, D.H.; Pei, A. Change of Rare-Earth Elements (REEs) Forms Using Them as Fertilizers. Agro-Environ. Prot. 2001, 20, 319–321. [Google Scholar]
- Wang, X.; Liu, D. Integration of cerium chemical forms and subcellular distribution to understand cerium tolerance mechanism in the rice seedlings. Environ. Sci. Pollut. Res. 2017, 24, 16336–16343. [Google Scholar] [CrossRef]
- Ding, S.; Liang, T.; Yan, J.; Zhang, Z.; Huang, Z.; Xie, Y. Fractionations of rare earth elements in plants and their conceptive model. Sci. China Ser. C Life Sci. 2007, 50, 47–55. [Google Scholar] [CrossRef]
- Wang, L.F. Studies on the Photosynthetic Characterizations and Distributions of Rear Earth Elements in Fern Dicranopteris Dichotoma. Ph.D. Thesis, Institute of Botany, Chinese Academy of Sciences, Beijing, China, 2005. [Google Scholar]
- Liang, M.X.; Chen, Z.B.; Chen, Z.Q.; Ren, T.J.; Qu, X. Effects of Vegetation Restoration on REEs Migration and Control in Red Soil Erosion Area in Southern China. Chin. Rare Earths 2022, 43, 64–72. [Google Scholar]
- Shan, X.; Wang, H.; Zhang, S.; Zhou, H.; Zheng, Y.; Yu, H.; Wen, B. Accumulation and uptake of light rare earth elements in a hyperaccumulator Dicropteris dichotoma. Plant Sci. 2003, 165, 1343–1353. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, Z.; Bai, L. Rare earth element migration in gullies with different Dicranopteris dichotoma covers in the Huangnikeng gully group, Changting County, Southeast China. Chemosphere 2016, 164, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.M.; Yusoff, I.; Abu Bakar, N.K.; Abu Bakar, A.F.; Alias, Y.; Mispan, M.S. Accumulation, Uptake and Bioavailability of Rare Earth Elements (Rees) in Soil Grown Plants from Ex-Mining Area in Perak, Malaysia. Appl. Ecol. Environ. Res. 2017, 15, 117–133. [Google Scholar] [CrossRef]
- Yuan, Y.; Huang, H.L.; Liu, X.W. Influences of inorganic fertilizer on the effective constituent content. Jiangxi For. Sci. Technol. 2000, 2, 29–30. [Google Scholar]
- Zhou, C.Z. Taxonomy and Systematic Research of Asarum Plants and Genuine Medicine. Ph.D. Thesis, Beijing University of Chinese Medicine, Beijing, China, 1994. [Google Scholar]
- Lei, W.; Shui, X.; Zhou, Y.; Tang, S.; Sun, M. Effects of praseodymium on flavonoids production and its biochemical mechanism of Scutellaria viscidula hairy roots in vitro. Pak. J. Bot. 2011, 43, 2387–2390. [Google Scholar]
- Zhou, J.; Fang, L.; Li, X.; Guo, L.; Huang, L. Jasmonic acid (JA) acts as a signal molecule in LaCl3-induced baicalin synthesis in Scutellaria baicalensis seedlings. Biol. Trace Elem. Res. 2012, 148, 392–395. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhang, Y.; Jia, X.Y.; Sun, B.X.; Zhang, R.R.; Ma, C.G.; Liang, J.P. Nutrient uptake rules of Polygala tenuifolia and its relationship with accumulation of bioactive components. J. Plant Nutr. Fertil. 2019, 25, 1230–1238. [Google Scholar]
- Fang, Y.M.; Cui, M.Y.; Liu, J.; Pei, T.L.; Wei, Y.K.; Zhao, Q. Study advance in biosynthesis of flavone from Scutellaria. China J. Chin. Mater. Med. 2020, 43, 4819–4826. [Google Scholar]
- Guo, M.; Wu, Z.L.; Wang, C.G.; Gao, X.Y. Synthesis and anti-tumor activity of baicalin-metal complex. Acta Pharm. Sin. 2014, 49, 337–345. [Google Scholar]
- Wang, Z.L.; Wang, S.; Kuang, Y.; Hu, Z.M.; Qiao, X.; Ye, M. A comprehensive review on phytochemistry, pharmacology, and flavonoid biosynthesis of Scutellaria baicalensis. Pharm. Biol. 2018, 56, 465–484. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.Y.; Sun, X.M.; Jia, F.C.; Wang, Y.L.; Li, D.J.; Li, J. The eco-geochemical characteristics of germanium and its relationship with the genuine medicinal material Scutellaria baicalensis in Chengde, Hebei Province. Geol. China 2020, 47, 1646–1667. [Google Scholar]
- Bai, Y.C.; Wei, X.F.; Chen, L.; Wan, R.M.; Hou, Z.X. Multivariate Analysis of Fruit Leaf Mineral Elements, Soil Fertility Factors and Fruit Quality of Vaccinium uliginosum L. Sci. Agric. Sin. 2018, 51, 170–181. [Google Scholar]
- Guo, B.S. Recent Research Advance of Rare Earth in the Field of Biology. Chin. Rare Earths 1999, 20, 64–68. [Google Scholar]
- Hong, F.S.; Fang, N.H.; Gu, Y.H.; Zhao, G.W. Effect of Cerium Nitrate on Seed Vigor and Activitiesof Enzymes during Germination of Rice. Chin. Rare Earths 1999, 20, 45–47. [Google Scholar]
- Jiang, L.; Zhang, W.C.; Wei, Z.L. Effects of micronutrient elements on cell growth and formation of flavonol glycosides of ginkgo callus. Hubei Agric. Sci. 1999, 38, 45–47. [Google Scholar]
- Yuan, X.F.; Wang, Q.; Zhao, B.; Wang, Y.C. Promotion of Cell Growth and Flavonoids Production in Saussurea medusa Cell Suspension Cultures by Rare Earth Elements. Chin. J. Process. Eng. 2004, 4, 325–329. [Google Scholar]
- Wang, J.N.; Huang, Y.H.; Mou, Z.M.; Wang, Y.W. Research Progress on Flavonoid of the Plant Secondary Metabolites. Sci. Seric. 2007, 33, 499–505. [Google Scholar]
- Wang, L.; Cheng, M.; Chu, Y.; Li, X.; Chen, D.D.; Huang, X.; Zhou, Q. Responses of plant calmodulin to endocytosis induced by rare earth elements. Chemosphere 2016, 154, 408–415. [Google Scholar] [CrossRef]
- Wang, L.; Li, J.; Zhou, Q.; Yang, G.; Ding, X.L.; Li, X.; Cai, C.X.; Zhang, Z.; Wei, H.Y.; Lu, T.H.; et al. Rare earth elements activate endocytosis in plant cells. Proc. Natl. Acad. Sci. USA 2014, 111, 12936–12941. [Google Scholar] [CrossRef] [Green Version]
- Ben, Y.; Cheng, M.; Wang, L.; Zhou, Q.; Yang, Z.; Huang, X. Low-dose lanthanum activates endocytosis, aggravating accumulation of lanthanum or/and lead and disrupting homeostasis of essential elements in the leaf cells of four edible plants. Ecotoxicol. Environ. Saf. 2021, 221, 112429. [Google Scholar] [CrossRef] [PubMed]
- Ge, Z.Q.; Li, J.C.; Yuan, Y.J.; Hu, Z.D. Effects of Ce (4+) on DNA Content and PAL Activity of Cell in Suspension Culture of Taxus Chinensis Var. Mairei. Chin. Rare Earths 2000, 21, 35–37. [Google Scholar]
- Fang, Y.; Lv, W.M.; Hu, D.D.; Min, S.Z.; Cao, Z.Q. Interactions between baicalein and La(III), Nd(III), Sm(III), Yb(III) and Y(III). J. Shaanxi Norm. Univ. (Nat. Sci. Ed.) 1993, 21, 44–48. [Google Scholar]
- Hu, Z.; Richter, H.; Sparovek, G.; Schnug, E. Physiological and biochemical effects of rare earth elements on plants and their agricultural significance: A review. J. Plant Nutr. 2004, 27, 183–220. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, Q.; Zhu, M.; Wang, L.; Zhou, Q.; Yang, Z.; Huang, X. Endocytosis in microcystis aeruginosa accelerates the synthesis of microcystins in the presence of lanthanum (III). Harmful Algae 2020, 93, 101791. [Google Scholar] [CrossRef]
- Tang, L.; Lin, J.H.; Nian, G.X.; Li, Y.M.; Lin, Y.L.; Wang, S.; Liu, D.H. Effects of Cu, Zn and Se on contents of total flavonlid, chlorogenic acid and Se in the flower of Chrysanthemum morifolium Ramat. Plant Nutr. Fertil. Sci. 2009, 15, 1475–1480. [Google Scholar]
- Yang, S.H.; Liu, X.F.; Guo, D.A.; Zheng, J.H. Effect of Rare-earth Element Eu (3+) on Callus Growth and Flavonoids Content in Glycyrrhiza uralensis. J. Chin. Med. Mater. 2005, 28, 533–534. [Google Scholar]
- Ynag, S.H.; Liu, X.F.; Guo, D.A.; Zheng, J.H. Effects of Different Additives on Accumulation of Flavonoids in Glycyrrhiza uralensis Callus. Chin. Pharm. J. 2006, 41, 96–99. [Google Scholar]
- Wang, L.; Lu, A.; Lu, T.; Ding, X.; Huang, X. Interaction between lanthanum ion and horseradish peroxidase in vitro. Biochimie 2010, 92, 41–50. [Google Scholar] [CrossRef]
- Batistič, O.; Kudla, J. Analysis of calcium signaling pathways in plants. Biochim. Biophys. Acta (BBA) Gen. Subj. 2012, 1820, 1283–1293. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Habitats | Median (25–75%) (g·g) | Average (g·g) | Skewness | CV/% | p Value |
|---|---|---|---|---|---|---|
| REE | NGHs (n = 11) | 174 (127–212) | 173 | 0.754 | 33.9 | 0.541 |
| APHs (n = 10) | 149 (140–156) | 149 | 0.231 | 6.84 | 0.753 | |
| BCHs (n = 8) | 187 (177–337) | 238 | 0.726 | 34.4 | 0.007 | |
| LREE | NGHs (n = 11) | 154 (115–194) | 157 | 0.765 | 35.1 | 0.673 |
| APHs (n = 10) | 133 (124–139) | 133 | 0.120 | 6.88 | 0.791 | |
| BCHs (n = 8) | 166 (158–311) | 216 | 0.761 | 36.8 | 0.009 | |
| HREE | NGHs (n = 11) | 15.2 (12.1–19.9) | 16.2 | 0.131 | 29.7 | 0.802 |
| APHs (n = 10) | 16.7 (15.3–18.1) | 16.6 | −0.158 | 8.84 | 0.701 | |
| BCHs (n = 8) | 20.2 (19.5–24.7) | 21.7 | 1.49 | 16.3 | 0.008 | |
| LREE\HREE | NGHs (n = 11) | 9.76 (7.85–11.5) | 9.85 | 0.156 | 25.2 | 0.814 |
| APHs (n = 10) | 7.86 (7.64–8.30) | 8.01 | 1.89 | 7.00 | 0.016 | |
| BCHs (n = 8) | 8.36 (7.96–11.7) | 9.84 | 1.73 | 29.8 | 0.005 | |
| La\Yb | NGHs (n = 11) | 11.4 (8.49–15.9) | 12.4 | 1.03 | 46.0 | 0.315 |
| APHs (n = 10) | 8.82 (8.34–10.1) | 9.27 | 1.37 | 12.7 | 0.042 | |
| BCHs (n = 8) | 9.53 (8.87–17.6) | 13.4 | 1.96 | 59.7 | 0.001 | |
| Eu | NGHs (n = 11) | 0.900 (0.710–0.970) | 0.869 | −0.168 | 14.4 | 0.385 |
| APHs (n = 10) | 0.719 (0.704–0.791) | 0.743 | 1.31 | 8.05 | 0.057 | |
| BCHs (n = 8) | 0.686 (0.652–0.793) | 0.699 | 0.032 | 18.9 | 0.512 | |
| Ce | NGHs (n = 11) | 0.950 (0.890–0.970) | 0.919 | −2.09 | 8.15 | 0.002 |
| APHs (n = 10) | 0.910 (0.870–0.935) | 0.892 | −2.20 | 7.89 | 0.003 | |
| BCHs (n = 8) | 0.950 (0.935–0.990) | 0.959 | 0.490 | 5.18 | 0.510 |
| Habitats | Profile | Layer | Depth (cm) | REE (g·g) | LREE (g·g) | HREE (g·g) | LREE/HREE | La/Yb | Eu | Ce |
|---|---|---|---|---|---|---|---|---|---|---|
| NGHs (hornblende gneiss) | Profile 1 | Soil | 0–20 | 169 | 150 | 18.2 | 8.28 | 9.52 | 0.734 | 0.872 |
| regolith | 20–30 | 259 | 235 | 23.8 | 9.88 | 11.6 | 0.596 | 0.895 | ||
| regolith | 50–70 | 259 | 237 | 21.2 | 11.2 | 13.8 | 0.572 | 0.889 | ||
| regolith | 260–270 | 255 | 231 | 24.6 | 9.37 | 9.9 | 0.650 | 0.902 | ||
| regolith | 280–300 | 243 | 220 | 23.2 | 9.47 | 10.5 | 0.587 | 0.842 | ||
| regolith | 310–320 | 289 | 259 | 30.3 | 8.54 | 9.24 | 0.538 | 0.888 | ||
| regolith | 340–360 | 242 | 221 | 21 | 10.5 | 12.9 | 0.612 | 0.883 | ||
| bedrock | 540–560 | 233 | 208 | 24.4 | 8.56 | 9.48 | 0.563 | 0.875 | ||
| Profile 2 | soil | 0–10 | 238 | 210 | 28.3 | 7.4 | 8.19 | 0.729 | 0.852 | |
| soil | 20–40 | 134 | 117 | 17.7 | 6.6 | 6.31 | 0.879 | 0.901 | ||
| regolith | 60–80 | 238 | 218 | 20.4 | 10.6 | 14.3 | 0.619 | 0.827 | ||
| regolith | 110–120 | 146 | 126 | 20.7 | 6.08 | 5.83 | 0.756 | 0.883 | ||
| regolith | 210–220 | 151 | 132 | 18.6 | 7.11 | 7.36 | 0.693 | 0.867 | ||
| regolith | 230–240 | 172 | 149 | 23.5 | 6.32 | 6.11 | 0.700 | 0.918 | ||
| regolith | 290–300 | 142 | 126 | 16.6 | 7.55 | 8.01 | 0.621 | 0.890 | ||
| regolith | 510–520 | 180 | 163 | 16.9 | 9.67 | 12.4 | 0.660 | 0.783 | ||
| regolith | 560–580 | 223 | 202 | 20.6 | 9.81 | 10.6 | 0.620 | 0.889 | ||
| Bedrock | 600 | 264 | 252 | 12.2 | 20.6 | 35.8 | 0.724 | 0.921 | ||
| APHs (loess) | profile 3 | soil | 0–20 | 119 | 107 | 12.6 | 8.43 | 9.87 | 0.841 | 0.990 |
| loess | 20–40 | 139 | 124 | 15.6 | 7.90 | 9.32 | 0.835 | 0.982 | ||
| BCHs (loess) | profile 4 | soil | 0–20 | 136 | 121 | 14.8 | 8.17 | 9.23 | 0.775 | 0.981 |
| loess | 20–40 | 133 | 118 | 14.6 | 8.11 | 9.26 | 0.799 | 0.968 |
| Parameters | Habitats | Median (25–75%) (mg·kg) | Average (mg·kg) | Skewness | CV/% | p Value |
|---|---|---|---|---|---|---|
| REE | NGHs (n = 11) | 2.02 (1.79–2.3) | 2.21 | 1.14 | 47.0 | 0.066 |
| APHs (n = 10) | 1.78 (1.55–2.31) | 1.94 | 1.01 | 30.3 | 0.484 | |
| BCHs (n = 8) | 2.04 (1.5–3.33) | 2.34 | 0.386 | 38.0 | 0.042 | |
| LREE | NGHs (n = 11) | 1.78 (1.62–2.11) | 2.00 | 1.24 | 49.2 | 0.056 |
| APHs (n = 10) | 1.6 (1.4–2.09) | 1.76 | 0.998 | 30.7 | 0.491 | |
| BCHs (n = 8) | 1.82 (1.36–3.12) | 2.15 | 0.441 | 39.8 | 0.039 | |
| HREE | NGHs (n = 11) | 0.212 (0.175–0.246) | 0.209 | −0.541 | 30.3 | 0.724 |
| APHs (n = 10) | 0.187 (0.146–0.217) | 0.187 | 1.07 | 28.1 | 0.376 | |
| BCHs (n = 8) | 0.183 (0.148–0.235) | 0.189 | 0.378 | 25.7 | 0.580 | |
| LREE\HREE | NGHs (n = 11) | 8.64 (7.55–10.8) | 9.27 | 1.48 | 23.7 | 0.063 |
| APHs (n = 10) | 9.27 (8.57–9.89) | 9.38 | 0.840 | 11.0 | 0.536 | |
| BCHs (n = 8) | 10.2 (9.13–14.7) | 11.27 | 0.556 | 26.2 | 0.283 | |
| La\Yb | NGHs (n = 11) | 15.2(12.4–20.9) | 17.26 | 1.07 | 43.4 | 0.291 |
| APHs (n = 10) | 16 (14.6–18.4) | 16.56 | 1.04 | 15.0 | 0.246 | |
| BCHs (n = 8) | 19.8 (16.5–32.9) | 24.58 | 1.10 | 43.6 | 0.168 | |
| Eu | NGHs (n = 11) | 1.28 (1.08–1.57) | 1.32 | −0.109 | 22.8 | 0.985 |
| APHs (n = 10) | 0.937 (0.854–1.14) | 0.998 | 0.787 | 16.0 | 0.185 | |
| BCHs (n = 8) | 1.06 (0.972–1.13) | 1.05 | −0.357 | 11.6 | 0.972 | |
| Ce | NGHs (n = 11) | 0.9 (0.730–0.950) | 0.845 | −1.00 | 15.7 | 0.192 |
| APHs (n = 10) | 0.935 (0.858–0.963) | 0.897 | −1.55 | 12.5 | 0.009 | |
| BCHs (n = 8) | 0.935 (0.835–1.04) | 0.933 | −0.241 | 16.7 | 0.933 |
| Flavonoids Compounds | Habitats | Average (mg·g) | Ranges (mg·g) | Skewness | p Value |
|---|---|---|---|---|---|
| Baicalin | NGHs (n = 11) | 27.5 | 12.8–51.7 | 1.42 | 0.325 |
| APHs (n = 10) | 8.28 | 4.22–13.2 | 0.546 | 0.911 | |
| BCHs (n = 8) | 11.0 | 4.04–17.1 | −0.411 | 0.273 | |
| Oroxylin A glycoside | NGHs (n = 11) | 1.85 | 0.270–4.03 | 1.06 | 0.326 |
| APHs (n = 10) | 0.895 | 0.030–1.83 | 0.136 | 0.601 | |
| BCHs (n = 8) | 0.724 | 0.080–1.62 | 0.633 | 0.726 | |
| Wogonoside | NGHs (n = 11) | 5.24 | 1.96–12.2 | 1.79 | 0.099 |
| APHs (n = 10) | 1.81 | 0.080–3.02 | −0.588 | 0.396 | |
| BCHs (n = 8) | 2.69 | 1.03–4.56 | 0.031 | 0.585 | |
| Baicalein | NGHs (n = 11) | 2.43 | 0.820–4.68 | 0.765 | 0.605 |
| APHs (n = 10) | 1.15 | 0.550–2.09 | 1.42 | 0.356 | |
| BCHs (n = 8) | 1.09 | 0.330–2.43 | 1.22 | 0.357 | |
| wogonin | NGHs (n = 11) | 0.966 | 0.310–1.75 | 0.476 | 0.508 |
| APHs (n = 10) | 0.468 | 0.200–0.810 | 0.862 | 0.598 | |
| BCHs (n = 8) | 0.478 | 0.090–1.19 | 1.38 | 0.301 | |
| Oroxylin A | NGHs (n = 11) | 0.386 | 0.200–0.560 | −0.036 | 0.61 |
| APHs (n = 10) | 0.23 | 0.060–0.440 | 0.378 | 0.561 | |
| BCHs (n = 8) | 0.184 | 0.030–0.570 | 1.92 | 0.031 |
| Micronutrient Elements | Habitats | Median (25∼75%) (mg·kg) | Average (mg·kg) | p Value |
|---|---|---|---|---|
| Cu | NGHs (n = 11) | 11 (9.08–14.5) | 12.2 | 0.277 |
| APHs (n = 10) | 10.1 (7.59–11) | 9.44 | 0.190 | |
| BCHs (n = 8) | 9.09 (6.85–11.8) | 9.24 | 0.310 | |
| Zn | NGHs (n = 11) | 15.9 (14.1–16.9) | 15.3 | 0.806 |
| APHs (n = 10) | 11.5 (10.53–13.2) | 11.8 | 0.373 | |
| BCHs (n = 8) | 12.4 (11.58–17) | 13.6 | 0.084 | |
| Sr | NGHs (n = 11) | 48 (28.35–51.3) | 48.3 | 0.039 |
| APHs (n = 10) | 22.2 (20.8–25.7) | 22.9 | 0.873 | |
| BCHs (n = 8) | 28.4 (22.26–34.7) | 28.4 | 0.479 | |
| Ge | NGHs (n = 11) | 1.21 (0.963–1.45) | 1.28 | 0.004 |
| APHs (n = 10) | 0.837 (0.777–0.995) | 0.910 | 0.095 | |
| BCHs (n = 8) | 1.15 (0.829–1.57) | 1.17 | 0.929 | |
| Se | NGHs (n = 11) | 0.064 (0.045–0.069) | 0.0592 | 0.818 |
| APHs (n = 10) | 0.06 (0.045–0.063) | 0.0550 | 0.686 | |
| BCHs (n = 8) | 0.045 (0.04–0.069) | 0.0510 | 0.025 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Z.; Shen, W.; Fang, W.; Zhang, H.; Chen, Z.; Xiong, L.; An, T. The Quality of Scutellaria baicalensis Georgi Is Effectively Affected by Lithology and Soil’s Rare Earth Elements (REEs) Concentration. Appl. Sci. 2023, 13, 3086. https://doi.org/10.3390/app13053086
Sun Z, Shen W, Fang W, Zhang H, Chen Z, Xiong L, An T. The Quality of Scutellaria baicalensis Georgi Is Effectively Affected by Lithology and Soil’s Rare Earth Elements (REEs) Concentration. Applied Sciences. 2023; 13(5):3086. https://doi.org/10.3390/app13053086
Chicago/Turabian StyleSun, Zijian, Wei Shen, Weixuan Fang, Huiqiong Zhang, Ziran Chen, Lianghui Xiong, and Tianhao An. 2023. "The Quality of Scutellaria baicalensis Georgi Is Effectively Affected by Lithology and Soil’s Rare Earth Elements (REEs) Concentration" Applied Sciences 13, no. 5: 3086. https://doi.org/10.3390/app13053086