

Effects of Vertically Heterogeneous Soil Salinity on Genetic Polymorphism and Productivity of the Widespread Halophyte Bassia prostrata
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
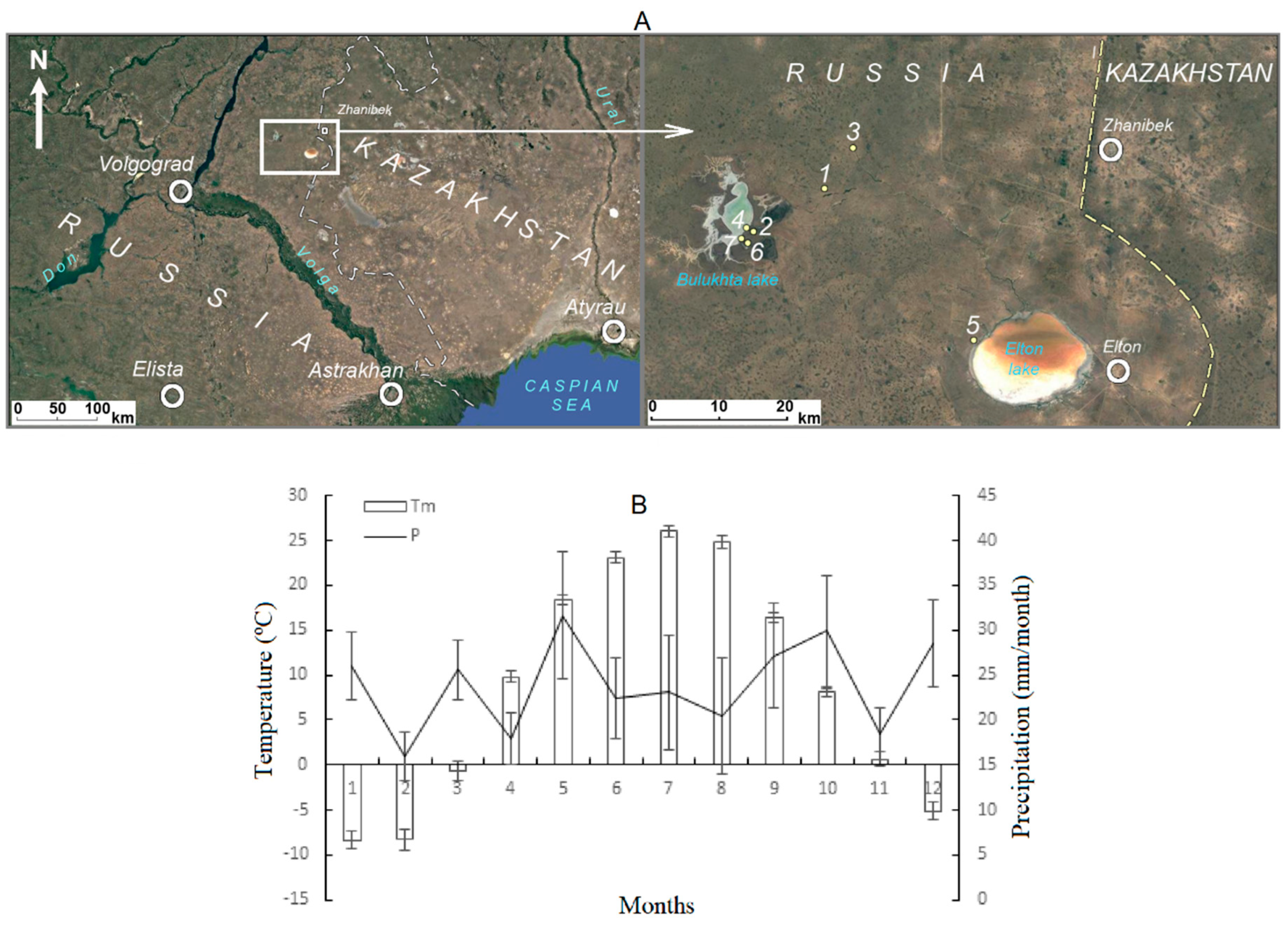
2.1. Study Area
2.2. Plant Sampling
2.3. Soil Sampling and Analysis
2.4. Plant Biomass and Water Content
2.5. Population Genetic Analysis
2.6. Statistical Analysis
3. Results
3.1. Soil Characteristics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitats, No | Soil Layers | Anions, cmol(eq)/kg | Cations, cmol(eq)/kg | Ssalt, % | Salinity Level | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| HCO3− | Cl− | SO42− | Ca2+ | Mg2+ | Na+ | K+ | ||||
| 1 | 0–20 | 1.04 ± 0.08 | 0.26 ± 0.02 | 0.78 ± 0.04 | 0.78 ± 0.02 | 0.52 ± 0.04 | 0.56 ± 0.01 | 0.22 ± 0.04 | 0.15 | non-saline |
| 20–40 | 1.25 ± 0.11 | 0.17 ± 0.01 | 0.52 ± 0.03 | 0.52 ± 0.01 | 0.26 ± 0.03 | 1.45 ± 0.12 | 0.02 ± 0.01 | 0.16 | non-saline | |
| 40–60 | 2.18 ± 0.17 | 0.26 ± 0.03 | 0.52 ± 0.03 | 0.52 ± 0.02 | 0.78 ± 0.08 | 1.74 ± 019 | 0.03 ± 0.01 | 0.23 | slight | |
| 2 | 0–20 | 0.10 ± 0.02 | 0.17 ± 0.01 | 0.52 ± 0.03 | 0.26 ± 0.01 | 0.26 ± 0.01 | 0.24 ± 0.01 | 0.03 ± 0.01 | 0.05 | non-saline |
| 20–40 | 0.31 ± 0.02 | 0.34 ± 0.04 | 0.26 ± 0.01 | 0.26 ± 0.02 | 0.26 ± 0.01 | 0.32 ± 0.04 | 0.07 ± 0.01 | 0.06 | non-saline | |
| 40–60 | 0.94 ± 0.13 | 1.89 ± 0.14 | 1.82 ± 0.23 | 0.78 ± 0.66 | 0.78 ± 0.32 | 3.15 ± 0.07 | 0.04 ± 0.01 | 0.31 | slight | |
| 3 | 0–20 | 1.35 ± 0.08 | 0.34 ± 0.07 | 0.52 ± 0.01 | 0.52 ± 0.02 | 0.26 ± 0.01 | 1.31 ± 0.04 | 0.12 ± 0.01 | 0.17 | slight |
| 20–40 | 1.77 ± 0.21 | 5.41 ± 0.43 | 1.04 ± 0.08 | 0.52 ± 0.01 | 0.78 ± 0.03 | 7.01 ± 0.16 | 0.01 ± 0.01 | 0.50 | moderate | |
| 40–60 | 0.52 ± 0.02 | 10.22 ± 0.93 | 18.98 ± 0.21 | 9.88 ± 0.10 | 8.32 ± 0.72 | 11.48 ± 0.81 | 0.04 ± 0.01 | 1.87 | extreme | |
| 4 | 0–20 | 0.94 ± 0.10 | 0.26 ± 0.03 | 1.56 ± 0.33 | 2.08 ± 0.50 | 0.26 ± 0.09 | 0.14 ± 0.07 | 0.27 ± 0.05 | 0.20 | slight |
| 20–40 | 0.57 ± 0.04 | 0.17 ± 0.01 | 15.60 ± 0.42 | 12.09 ± 0.74 | 3.12 ± 0.41 | 1.84 ± 0.09 | 0.15 ± 0.01 | 1.11 | moderate | |
| 40–60 | 0.52 ± 0.03 | 0.69 ± 0.08 | 18.72 ± 1.53 | 9.88 ± 0.51 | 3.64 ± 0.33 | 6.27 ± 0.71 | 0.14 ± 0.02 | 1.35 | high | |
| 5 | 0–20 | 0.73 ± 0.17 | 0.49 ± 0.0.1 | 0.35 ± 0.07 | 0.43 ± 0.08 | 0.43 ± 0.11 | 0.74 ± 0.26 | 0.06 ± 0.02 | 0.11 | non-saline |
| 20–40 | 1.25 ± 0.76 | 7.90 ± 0.06 | 2.60 ± 0.32 | 1.04 ± 0.09 | 0.52 ± 0.07 | 10.28 ± 0.93 | 0.01 ± 0.01 | 0.75 | moderate | |
| 40–60 | 0.31 ± 0.02 | 7.39 ± 0.07 | 23.92 ± 1.30 | 10.14 ± 0.12 | 6.76 ± 0.54 | 14.77 ± 1.10 | 0.05 ± 0.01 | 2.06 | extreme | |
| 6 | 0–20 | 0.78 ± 0.13 | 0.39 ± 0.04 | 0.91 ± 0.11 | 0.52 ± 0.01 | 0.52 ± 0.01 | 0.78 ± 0.07 | 0.26 ± 0.09 | 0.15 | non-saline |
| 20–40 | 1.77 ± 0.17 | 3.18 ± 1.05 | 4.03 ± 0.98 | 1.04 ± 0.21 | 0.39 ± 0.11 | 7.50 ± 1.94 | 0.04 ± 0.01 | 0.76 | moderate | |
| 40–60 | 0.42 ± 0.03 | 7.65 ± 1.11 | 22.36 ± 1.74 | 11.70 ± 1.03 | 8.06 ± 0.92 | 10.67 ± 1.05 | 0.10 ± 0.03 | 1.95 | extreme | |
| 7 | 0–20 | 1.51 ± 0.04 | 1.42 ± 0.74 | 1.69 ± 0.32 | 0.52 ± 0.01 | 0.39 ± 0.11 | 3.51 ± 1.01 | 0.20 ± 0.11 | 0.25 | slight |
| 20–40 | 0.73 ± 0.04 | 9.02 ± 1.08 | 8.58 ± 0.91 | 3.12 ± 0.07 | 2.08 ± 0.34 | 13.14 ± 1.11 | 0.09 ± 0.01 | 1.17 | high | |
| 40–60 | 0.62 ± 0.03 | 16.06 ± 1.77 | 23.66 ± 1.73 | 10.66 ± 0.85 | 6.50 ± 0.77 | 23.25 ± 2.76 | 0.04 ± 0.01 | 2.57 | extreme | |
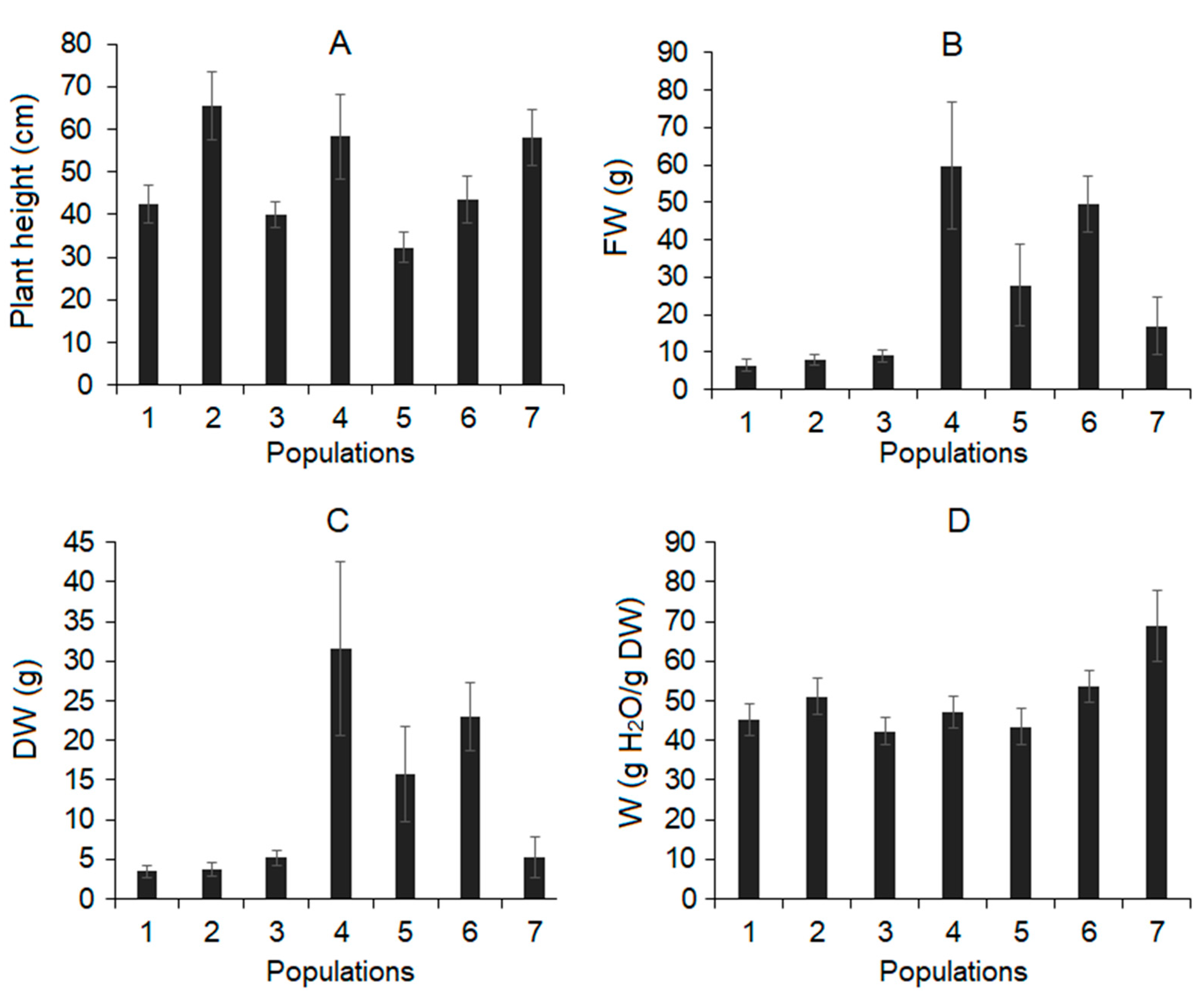
3.2. Plant Growth and Water Content

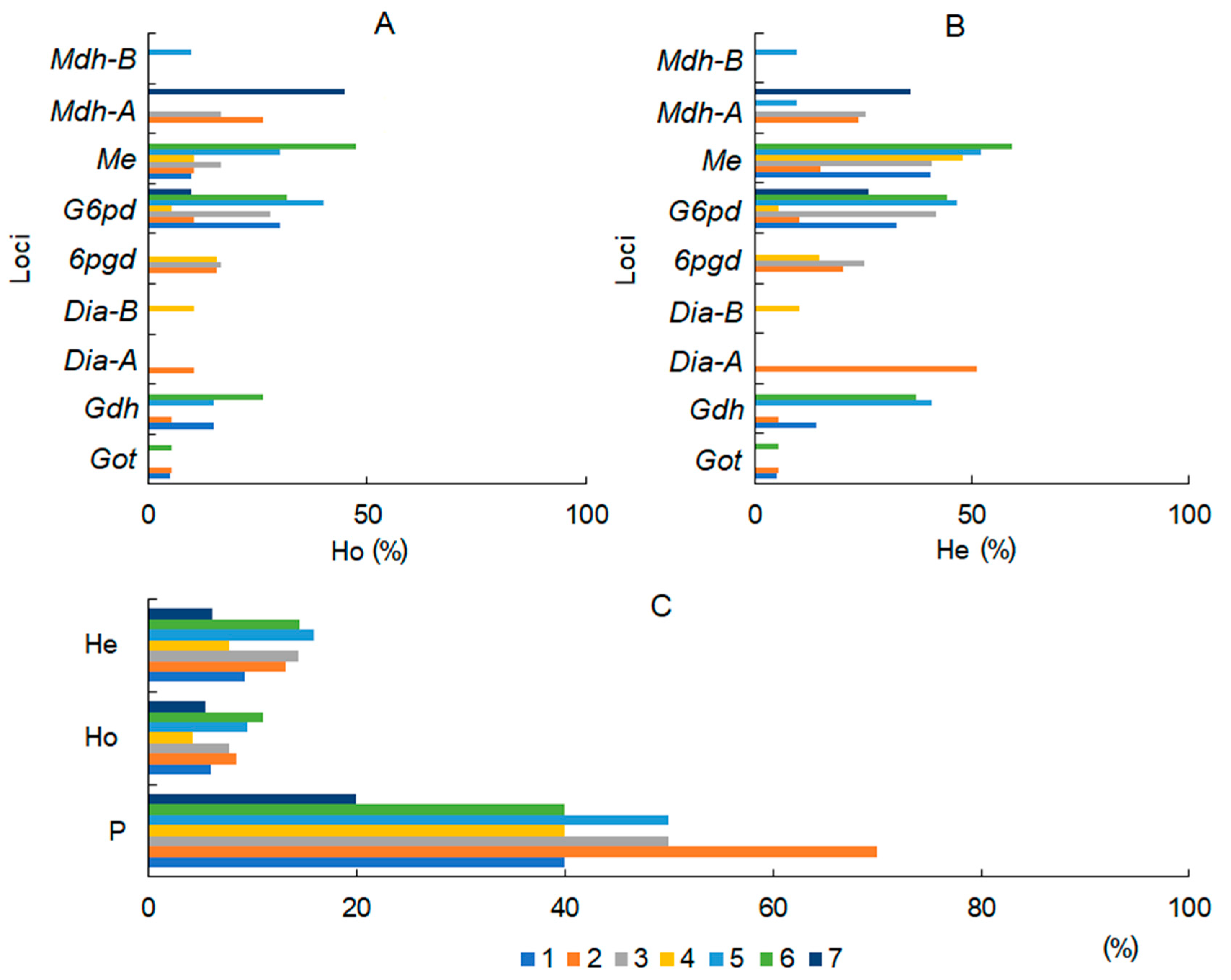
3.3. Population Genetic Diversity
3.4. Plant–Soil Interaction
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, J.; Ji, M.; Xie, Y.; Wang, S.; He, Y.; Ran, J. Global semi-arid climate change over last 60 years. Clim. Dyn. 2016, 46, 1131–1150. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Dantas, B.F.; Moura, M.S.B.; Pelacani, C.R.; Angelotti, F.; Taura, T.A.; Oliveira, G.M.; Bispo, J.S.; Matias, J.R.; Silva, F.F.S.; Pritchard, H.W.; et al. Rainfall, not soil temperature, will limit the seed germination of dry forest species with climate change. Oecologia 2020, 192, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Welegedara, N.P.Y.; Grant, R.F.; Quideau, S.A.; Landhäusser, S.M.; Merlin, M.; Lloret, E. Modelling plant water relations and net primary productivity as affected by reclamation cover depth in reclaimed forestlands of northern Alberta. Plant Soil 2020, 446, 627–654. [Google Scholar] [CrossRef]
- Kakeh, J.; Gorji, M.; Mohammadi, M.H.; Asadi, H.; Khormali, F.; Sohrabi, M.; Cerdà, A. Biological soil crusts determine soil properties and salt dynamics under arid climatic condition in Qara Qir, Iran. Sci. Total Environ. 2020, 732, 139168. [Google Scholar] [CrossRef] [PubMed]
- Pankova, E.; Vorob’eva, L.; Gadjiev, I.; Gorohova, I.N.; Elizarova, T.N.; Koroluk, T.V.; Lopatovskaya, O.G.; Novikova, A.F.; Reshetov, G.G.; Skripnikova, M.I.; et al. Saline Soils in Russia; Akademkniga: Moscow, Russia, 2006; ISBN 5-94628-198-4. [Google Scholar]
- Flowers, T.; Galal, H.; Bromham, L. Evolution of halophytes: Multiple origins of salt tolerance in land plants. Funct. Plant Biol. 2010, 37, 604–612. [Google Scholar] [CrossRef]
- Yamanaka, N.; Toderich, K. Photobooks of Drylands Vol.4 Salinization in Drylands; Imai Print Co. Ltd.: Tottori, Japan, 2020; ISBN 978-4-86611-188-9. Available online: https://catalogue.unccd.int/1484_Salinization_in_Dryland_Yamanaka_Toderich_2020.pdf (accessed on 31 March 2020)(In Japanese and English).
- Nikalje, G.C.; Yadav, K.; Penna, S. Halophyte Responses and Tolerance to Abiotic Stresses. In Ecophysiology, Abiotic Stress Responses and Utilization of Halophytes; Hasanuzzaman, M., Nahar, K., Öztürk, M., Eds.; Springer: Singapore, 2019; pp. 1–23. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: Adaptation in halophytes. Ann. Bot. 2015, 155, 327–331. [Google Scholar] [CrossRef] [Green Version]
- Ventura, Y.; Eshel, A.; Pasternak, D.; Sagi, M. The development of halophyte-based agriculture: Past and present. Ann. Bot. 2015, 115, 529–540. [Google Scholar] [CrossRef] [Green Version]
- Tigano, A.; Friesen, V.L. Genomics of local adaptation with gene flow. Mol. Ecol. 2016, 25, 2144–2164. [Google Scholar] [CrossRef]
- Thalmann, M.; Santelia, D. Starch as a determinant of plant fitness under abiotic stress. New Phytol. 2017, 214, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Sun, K.; Hunt, K.; Hauser, B.A. Ovule abortion in Arabidopsis triggered by stress. Plant Physiol. 2004, 135, 2358–2367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prentice, H.; Lonn, M.; Lager, H.; Rosen, E.; van der Maarel, E. Changes in allozyme frequencies in Festuca ovina populations after a 9-year nutrient/water experiment. J. Ecol. 2000, 88, 331–347. [Google Scholar] [CrossRef]
- Mandak, B.; Bimova, K.; Plackova, I. Genetic structure of experimental populations and reproductive fitness in a heterocarpic plant Atriplex tatarica (Chenopodiaceae). Am. J. Bot. 2006, 93, 1640–1649. [Google Scholar] [CrossRef] [PubMed]
- Nevo, E.; Brown, A.; Zohary, D.; Storch, N.; Beiles, A. Microgeographic edaphic differentiation an allozyme polymorphysms of wild barley (Hordeum spontaneum, Poaceae). Plant Syst. Evol. 1981, 138, 287–292. [Google Scholar] [CrossRef]
- Nevo, E.; Krugman, T.; Beiles, A. Edaphic natural selection of allozyme polymorphisms in Aegilops peregrina at a Galilee microsite in Israel. Heredity 1994, 72, 109–112. [Google Scholar] [CrossRef]
- Prentice, H.; Lonn, M.; Lefkovitch, L.; Runyeon, H. Associations between allele frequencies in Festuca ovina and habitat variation in the alvar grass-lands on the Baltic island of Oland. J. Ecol. 1995, 83, 391–401. [Google Scholar] [CrossRef]
- Sun, X.-S.; Chen, Y.-H.; Zhuo, N.; Cui, Y.; Luo, F.-L.; Zhang, M.-X. Effects of salinity and concomitant species on growth of Phragmites australis populations at different levels of genetic diversity. Sci. Total Environ. 2021, 780, 146516. [Google Scholar] [CrossRef]
- Aavik, T.; Helm, A. Restoration of plant species and genetic diversity depends on landscape-scale. Restor. Ecol. 2018, 26, S92–S102. [Google Scholar] [CrossRef]
- Song, X.; Gao, X.; Wu, P.; Zhao, X.; Zhang, W.; Zou, Y.; Siddique, K.H.M. Drought responses of profile plant-available water and fine-root distributions in apple (Malus pumila Mill.) orchards in a loessial, semiarid, hilly area of China. Sci. Total Environ. 2020, 723, 137739. [Google Scholar] [CrossRef]
- Shuyskaya, E.V.; Toderich, K.N.; Voinitska-Poltorak, A. Genetic variation of Kochia prostrata (L.) Schrad. in the arid zone of Uzbekistan. Probl. Osvoeniya Pustyn. 2001, 3, 24–31. [Google Scholar]
- Gintzburger, G.; Toderich, K.N.; Mardonov, B.K.; Makhmudov, M.M. Rangelands of the Arid and Semi-Arid Zones in Uzbekistan; Centre de Cooperation Internationale en Resherche Agronomique pour le Development (CIRAD): Monpellier, France, 2003. [Google Scholar]
- Akhzari, D.; Sepehry, A.; Pessarakli, M.; Barani, H. Studying the effects of salinity stress on the growth of various halophytic plant species (Agropyron elongatum, Kochia prostrata and Puccinellia distans). World Appl. Sci. J. 2012, 16, 998–1003. [Google Scholar]
- Toderich, K.N.; Shuyskaya, E.V.; Taha, F.K.; Matsuo, N.; Ismail, S.; Aralova, D.B.; Radjabov, T.F. Integrating Agroforestry and Pastures for Soil Salinity Management in Dryland Ecosystems in Aral Sea Basin. In Developments in Soil Salinity Assessment and Reclamation; Shahid, S., Abdelfattah, M., Taha, F., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 579–602. [Google Scholar]
- Shuyskaya, E.; Rakhmankulova, Z.; Prokofieva, M.; Saidova, L.; Toderich, K.; Voronin, P. Intensity and duration of salinity required to form adaptive response in C4 halophyte Kochia prostrata (L.) Shrad. Front. Plant Sci. 2022, 13, 955880. [Google Scholar] [CrossRef] [PubMed]
- Shuyskaya, E.V.; Nukhimovskaya, Y.D.; Lebedeva, M.P.; Churilina, A.E.; Kolesnikov, A.V. Effect of soil conditions on the level of genetic diversity in the xerohalophyte Kochia prostrata (L.) Schrad. (Chenopodiaceae). Russ. J. Ecol. 2020, 51, 118–126. [Google Scholar] [CrossRef]
- Konyushkova, M.; Kozlov, D. Automated analysis of the distribution of dark-colored chernozemlike soils in the Northern Caspian region based on satellite imaging data: The example of the Dzhanybek station. Arid Ecosyst. 2011, 1, 65–73. [Google Scholar] [CrossRef]
- Balyan, G. Kochia Prostrata in Kyrgyzstan; Kyrgyzstan: Frunze, Kyrgyzstan, 1972. [Google Scholar]
- Waldron, B.; Eun, J.; ZoBell, D.; Olson, K. Forage kochia (Kochia prostrata) for fall and winter grazing. Small Rumin. Res. 2010, 91, 47–55. [Google Scholar] [CrossRef]
- Vorob’eva, L.A. Chemical Analysis of Soils; Mosk Gos Univ: Moscow, Russia, 1998. [Google Scholar]
- Spooner, D.; van Treuren, R.; de Vicente, M. Molecular Markers for Genebank Management; IPGRI Technical Bulletin No. 10; International Plant Genetic Resources Institute: Rome, Italy, 2005. [Google Scholar]
- Marden, J.H. Nature’s inordinate fondness for metabolic enzymes: Why metabolic enzyme loci are so frequently targets of selection. Mol. Ecol. 2013, 22, 5743–5764. [Google Scholar] [CrossRef] [PubMed]
- Soltis, D.; Soltis, P. Isozymes in Plant Biology; Springer: London, UK, 1990. [Google Scholar]
- Harrison, R.D.; Chatterton, N.J.; Waldron, B.L.; Davenport, B.W.; Palazzo, A.J.; Horton, W.H.; Asay, K.H. Forage Kochia: Its Compatibility and Potential Aggressiveness on Intermountain Rangelands; Research Report 162; Utah Agricultural Experiment Station, Utah State University: Logan, UT, USA, 2000. [Google Scholar]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. Available online: https://www.jstor.org/stable/25150520 (accessed on 22 November 2022). [CrossRef]
- Karimi, G.; Ghorbanli, M.; Heidari, H.; Khavari Nejad, R.; Assareh, M. The effects of NaCl on growth, water relations, osmolytes and ion content in Kochia Prostrata. Biol. Plant. 2005, 49, 301–304. [Google Scholar] [CrossRef]
- Grigore, M.; Boscaiu, M.; Llinares, J.; Vocente, O. Mitigation of salt stress-induced inhibition of Plantago crassifolia reproductive development by supplemental calcium or magnesium. Not. Bot. Horti Agrobot. Cluj-Napoca 2012, 40, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Tian, W.; Wang, C.; Gao, Q.; Li, L.; Luan, S. Calcium spikes, waves and oscillations in plant development and biotic interactions. Nat. Plants 2020, 6, 750–759. [Google Scholar] [CrossRef]
- Shaul, O. Magnesium transport and function in plants: The tip of iceberg. Biometals 2002, 15, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.; Cakmak, I.; Wang, S.; Zhang, F.; Guo, S. Synergistic and antagonistic interactions between potassium and magnesium in higher plants. Crop. J. 2021, 9, 249–256. [Google Scholar] [CrossRef]
- Orlovsky, N.S.; Japakova, U.N.; Shulgina, I.; Volis, S. Comparative study of seed germination and growth of Kochia prostrata and Kochia scoparia (Chenopodiaceae) under salinity. J. Arid Environ. 2011, 75, 532–537. [Google Scholar] [CrossRef]
- Loveless, M.D.; Hamrick, J.L. Ecological determinants of genetic structure in plant populations. Ann. Rev. Syst. 1984, 15, 65–95. [Google Scholar] [CrossRef]
- North, A.; Pennanen, J.; Ovaskainen, O.; Laine, A.-L. Local adaptation in a changing world: The roles of gene-flow, mutation, and sexual reproduction. Evolution 2010, 65, 79–89. [Google Scholar] [CrossRef]
- Rakhmankulova, Z.F. Plant respiration and global climatic changes. Russ. J. Plant Physiol. 2022, 69, 109. [Google Scholar] [CrossRef]
- Kandoi, D.; Tripathy, B.C. Overexpression of chloroplastic Zea mays NADP-malic enzyme (ZmNADP-ME) confers tolerance to salt stress in Arabidopsis thaliana. Photosynth. Res. 2022. Available online: https://www.researchsquare.com/article/rs-2143178/v1 (accessed on 22 November 2022).
- Forester, B.R.; Jones, M.R.; Joost, S.; Landguth, E.L.; Lasky, J.R. Detecting spatial genetic signatures of local adaptation in heterogeneous landscapes. Mol. Ecol. 2016, 25, 104–120. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shuyskaya, E.; Toderich, K.; Kolesnikov, A.; Prokofieva, M.; Lebedeva, M. Effects of Vertically Heterogeneous Soil Salinity on Genetic Polymorphism and Productivity of the Widespread Halophyte Bassia prostrata. Life 2023, 13, 56. https://doi.org/10.3390/life13010056
Shuyskaya E, Toderich K, Kolesnikov A, Prokofieva M, Lebedeva M. Effects of Vertically Heterogeneous Soil Salinity on Genetic Polymorphism and Productivity of the Widespread Halophyte Bassia prostrata. Life. 2023; 13(1):56. https://doi.org/10.3390/life13010056
Chicago/Turabian StyleShuyskaya, Elena, Kristina Toderich, Alexander Kolesnikov, Maria Prokofieva, and Marina Lebedeva. 2023. "Effects of Vertically Heterogeneous Soil Salinity on Genetic Polymorphism and Productivity of the Widespread Halophyte Bassia prostrata" Life 13, no. 1: 56. https://doi.org/10.3390/life13010056