
The Coding Mitogenome of the Freshwater Crayfish Pontastacus leptodactylus (Decapoda:Astacidea:Astacidae) from Lake Vegoritida, Greece and Its Taxonomic Classification

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
3. Results
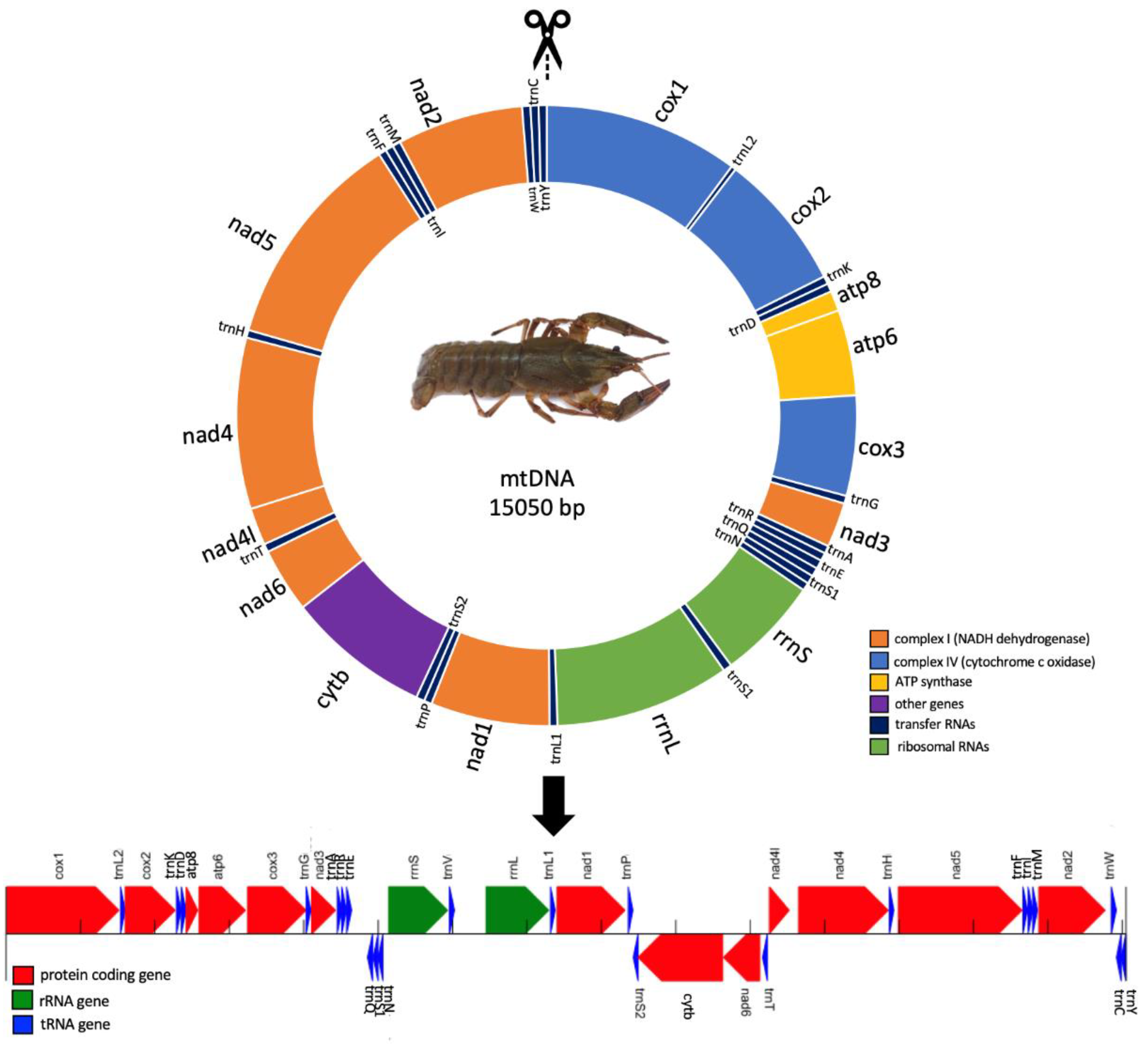
3.1. Genome Composition
3.2. Protein-Coding Genes, tRNAs, and rRNAs
3.3. Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gherardi, F.; Souty-Grosset, C. Pontastacus leptodactylus (Amended Version of 2016 Assessment). The IUCN Red List of Threatened Species 2017: E.T153745A120103207. Available online: https://www.iucnredlist.org/species/153745/120103207 (accessed on 8 December 2022).
- Diler, O.; Bolat, Y. Isolation of Acremonium species from crayfish, Astacus leptodactylus in Egirdir Lake. Bull. Eur. Assoc. Fish Pathol. 2001, 21, 164–168. [Google Scholar]
- Souty-Grosset C’Reynolds, J.D. Current ideas on methodological approaches in European crayfish conservation and restocking procedures. Knowl. Manag. Aquat. Ecosyst. 2009, 394–395, 1. [Google Scholar] [CrossRef]
- Harlioğlu, A.G.; Harlioğlu, M.M. The status of freshwater crayfish (Astacus leptodactylus Eschscholtz) fisheries in Turkey. Rev. Fish. Sci. 2009, 17, 187–189. [Google Scholar] [CrossRef]
- Alvanou, M.V.; Papadopoulos, D.K.; Lattos, A.; Georgoulis, I.; Feidantsis, K.; Apostolidis, A.P.; Michaelidis, B.; Giantsis, I. Biology, distribution, conservation status and stocking perspective of freshwater crayfish in Greece: An updated review. Aquac. Res. 2022, 53, 5115–5128. [Google Scholar] [CrossRef]
- Holdich, D.M.; Haffner, P.; Noël, P.Y.; Carral, J.; Füreder, L.; Gherardi, F.; Machino, Y.; Madec, J.; Pöckl, M.; Śmietana, P.; et al. Species files. In Atlas of Crayfish in Europe; Souty-Grosset, C., Holdich, D.M., Noël, P.V., Haffner, P., Eds.; Muséum Nationale d’Histoire Naturelle: Paris, France, 2006; pp. 50–129. [Google Scholar]
- Holdich, D.M.; Harlioğlu, M.M.; Firkins, I. Salinity Adaptations of Crayfish in British Waters with Particular Reference to Austropotamobius pallipes, Astacus leptodactylus and Pacifastacus leniusculus. Estuar. Coast. Shelf Sci. 1997, 44, 147–154. [Google Scholar] [CrossRef]
- Kokko, H.; Koistinen, L.; Harlioğlu, M.M.; Makkonen, J.; Aydın, H.; Jussila, J. Recovering Turkish narrow clawed crayfish (Astacus leptodactylus) populations carry Aphanomyces astaci. Knowl. Manag. Aquat. Ecosyst. 2012, 404, 12. [Google Scholar] [CrossRef] [Green Version]
- Perdikaris, C.; Konstantinidis, E.; Georgiadis, C.; Kouba, A. Freshwater crayfish distribution update and maps for Greece: Combining literature and citizen-science data. Knowl. Manag. Aquat. Ecosyst. 2017, 418, 51. [Google Scholar] [CrossRef] [Green Version]
- Laggis, A.; Baxevanis, A.D.; Charalampidou, A.; Maniatsi, S.; Triantafyllidis, A.; Abatzopoulos, T.J. Microevolution of the noble crayfish (Astacus astacus) in the Southern Balkan Peninsula. BMC Evol. Biol. 2017, 17, 122. [Google Scholar] [CrossRef] [Green Version]
- Gross, R.; Kõiv, K.; Pukk, L.; Kaldre, K. Development and characterization of novel tetranucleotide microsatellite markers in the noble crayfish (Astacus astacus) suitable for highly multiplexing and for detecting hybrids between the noble crayfish and narrow-clawed crayfish (A. leptodactylus). Aquaculture 2017, 472, 50–56. [Google Scholar] [CrossRef]
- Holdich, D.M.; Reynolds, J.D.; Souty-Grosset, C.; Sibley, P.J. A review of the ever increasing threat to European crayfish from non-indigenous crayfish species. Knowl. Manag. Aquat. Ecosyst. 2009, 394-395, 11. [Google Scholar] [CrossRef] [Green Version]
- Akhan, S.; Bektas, Y.; Berber, S.; Kalayci, G. Population structure and genetic analysis of narrow-clawed crayfish (Astacus leptodactylus) populations in Turkey. Genetica 2014, 142, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Kȯksal, G. Astacus leptodactylus in Europe. In Freshwater Crayfish. Biology, Management and Exploitation; Holdich, D.M., Lowery, R.S., Eds.; Croom Helm: London, UK, 1988; pp. 365–400. [Google Scholar]
- Karaman, M.S. Studie der Astacidae (Crustacea, Decapoda) II. Teil. Hydrobiologia 1963, 22, 111–132. [Google Scholar] [CrossRef]
- Strarobogatov, Y.I. Taxonomy and geographical distribution of crayfishes of Asia and East Europe (Crustacea Decapoda Astacoidei). Arthropoda Sel. 1995, 4, 3–25. [Google Scholar]
- Śmietana, P.; Schuz, H.K.; Keszka, S.; Schulz, R.; Schulz, H.K.; Keszka, S.; Schulz, R. A proposal for accepting Pontastacus as a genus of European crayfish within the family Astacidae based on a revision of the West and East European taxonomic literature. Bull. Français Pêche Piscic. 2006, 380–381, 1041–1052. [Google Scholar] [CrossRef] [Green Version]
- Kostyuk, V.S.; Garbar, A.V.; Mezhzherin, S.V. Karyotypes and morphological variability of crayfish Pontastacus leptodactylus and P. angulosus (Malacostraca, Decapoda). Vestn. Zool. 2013, 47, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Crandall, K.A.; De Grave, S. An updated classification of the freshwater crayfishes (Decapoda: Astacidea) of the world, with a complete species list. J. Crust. Biol. 2017, 37, 615–653. [Google Scholar] [CrossRef] [Green Version]
- Brodski, S.Y. On the systematics of palaearctic crayfishes (Crustacea, Astacidae). Freshw. Crayfish 1983, 5, 464–469. [Google Scholar]
- Grandjean, F.; Gouin, N.; Keith, P.; Noėl, P.; Persat, H.; Reynolds, J.; Schulz, H.; Smietana, P.; Souty-Grosset, C. Systematics and phylogeny of freshwater crayfish, with particular reference to historical biogeography of Europe. In Atlas of Crayfish in Europe; Souty-Grosset, C., Holdich, D.M., Noël, P.V., Haffner, P., Eds.; Muséum Nationale d’Histoire Naturelle: Paris, France, 2006; pp. 11–23. [Google Scholar]
- Wolstenholme, D.R. Animal mitochondrial DNA: Structure and evolution. Int. Rev. Cytol. 1992, 141, 173–216. [Google Scholar]
- Maguire, I.; Podnar, M.; Jelic, M.S.Ï.; Tambuk, A.; Schrimpf, A.; Schulz, H. Two distinct evolutionary lineages of the Astacus leptodactylus species-complex (Decapoda: Astacidae) inferred by phylogenetic analyses. Invertebr. Syst. 2014, 28, 117–123. [Google Scholar] [CrossRef]
- Khoshkholgh, M.R.; Nazari, S. Genetic variation in popula- tions of the narrow-clawed crayfish (Astacus leptodactylus) as assessed by PCR-RFLP of mitochondrial COI gene. Mol. Biol. Res. Commun. 2015, 4, 225–237. [Google Scholar]
- Khoshkholgh, M.R.; Nazari, S. Application of microsatellite markers for the delineation of the narrow-clawed crayfish (Astacus leptodactylus) populations from Caspian Sea Basin. WJFMS 2016, 8, 142–150. [Google Scholar]
- Blaha, M.; Patoka, J.; Japoshvili, B.; Let, M.; Buřič, M.; Kouba, A.; Mumladze, L. Genetic diversity, phylogenetic position and morphometric analysis of Astacus colchicus (Decapoda, Astacidae): A new insight into Eastern European crayfish fauna. Integr. Zool. 2021, 16, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.H.; Gan, H.M.; Lee, Y.P.; Grandjean, F.; Croft, L.J.; Austin, C.M. A giant genome for a giant crayfish (Cherax quadricarinatus) with insights into cox1 pseudogenes in decapod genomes. Front. Gen. 2020, 11, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, R.A.; Cruickshank, R.H. The seven deadly sins of DNA barcoding. Mol. Ecol. Resour. 2013, 13, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Buhay, J.E.; Whiting, M.F.; Crandall, K.A. Many species in one: DNA barcoding overestimates the number of species when nuclear mitochondrial pseudogenes are coamplified. PNAS 2008, 105, 13486–13491. [Google Scholar] [CrossRef] [Green Version]
- Hazkani-Covo, E.; Zeller, R.M.; Martin, W. Molecular poltergeists: Mitochondrial DNA copies (numts) in sequenced nuclear genomes. PLoS Genet. 2010, 6, e1e000834. [Google Scholar] [CrossRef]
- Munasinghe, D.H.N.; Murphy, N.P.; Austin, C.M. Utility of mitochondrial DNA sequences from four gene regions for systematic studies of Australian freshwater crayfish of the genus Cherax (Decapoda: Parastacidae). J. Crustacean Biol. 2003, 23, 402–417. [Google Scholar] [CrossRef] [Green Version]
- Khoshkholgh, M.; Nazari, S. The genetic diversity and differentiation of narrow-clawed crayfish Pontastacus leptodactylus (Eschscholtz, 1823) (Decapoda:Astacidea:Astacidae) in the Caspian Sea Basin, Iran as determined with mitochondrial and microsatellite DNA markers. J. Crust. Biol. 2019, 39, 112–120. [Google Scholar] [CrossRef]
- Jia, X.N.; Xu, S.X.; Bai, J.; Wang, Y.F.; Nie, Z.H.; Zhu, C.C.; Wang, Y.; Cai, Y.X.; Zou, J.X.; Zhou, X.M. The complete mitochondrial genome of Somanniathelphusa boyangensis and phylogenetic analysis of Genus Somanniathelphusa (Crustacea: Decapoda: Parathelphusidae). PLoS ONE 2018, 13, 0192601. [Google Scholar] [CrossRef] [Green Version]
- Cameron, S.L.; Lambkin, C.L.; Barker, S.C.; Whiting, M.F. A mitochondrial genome phylogeny of Diptera: Whole genome sequence data accurately resolve relationships over broad timescales with high precision. Syst. Entomol. 2007, 32, 40–59. [Google Scholar] [CrossRef]
- Kõressaar, T.; Lepamets, M.; Kaplinski, L.; Raime, K.; Andreson, R.; Remm, M. Primer3_masker: Integrating masking of template sequence with primer design software. Bioinformatics 2018, 34, 1937–1938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGAX: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de Novo Metazoan Mitochondrial Genome Annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Boore, J.L.; Brown, W.M. Big Trees from Little Genomes: Mitochondrial Gene Order as a Phylogenetic Tool. Curr. Opin. Genet. Dev. 1998, 8, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Gouin, N.; Grandjean, F.; Souty-Grosset, C. Disentangling the impact of demographic factors on population differentiation of an endangered freshwater crayfish (Austropotamobius pallipes) using population density and microsatellite data. Freshw. Biol. 2011, 56, 2105–2118. [Google Scholar] [CrossRef]
- Gouin, N.; Souty-Grosset, C.; Ropiquet, A.; Grandjean, F. High dispersal ability of Austropotamobius pallipes revealed by microsatellite markers in a French brook. Bull. Français Pêche Piscic. 2002, 367, 681–689. [Google Scholar] [CrossRef]
- Fetzner, J.W.; Crandall, K.A. Genetic variability within and among populations of the golden crayfish (Orconectes luteus): A comparison using amplified fragment length polymorphism (AFLPs) and mitochondrial 16s gene sequences. Freshw. Crayfish 1999, 12, 396–412. [Google Scholar]
- Palumbi, S.R. Genetic divergence, reproductive isolation, and marine speciation. Ecol. Evol. Syst. 1994, 25, 547–572. [Google Scholar] [CrossRef]
- Perdikaris, C.; Georgiadis, C. Co-occurrence of narrow-clawed crayfish (Astacus leptodactylus sensu lato) and noble crayfish (Astacus astacus L.) in the southwestern Balkans: The case of Lake Pamvotida (NW Greece). North West. J. Zool. 2017, 13, 18–26. [Google Scholar]
- Largiader, C.R.; Herger, F.; Lörtscher, M.; Scholl, A. Assessment of natural and artificial propagation of the white-clawed crayfish (Austropotamobius pallipes species complex) in the Alpine region with nuclear and mitochondrial markers. Mol. Ecol. 2000, 9, 25–37. [Google Scholar] [CrossRef]
- Schulz, R. Status of the noble crayfish Astacus astacus (L.) in Germany: Monitoring protocol and the use of RAPD markers to assess the genetic structure of populations. Bull. Français Pêche Piscic. 2000, 356, 123–138. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.J.; Cordes, J.F. DNA marker technologies and their applications in aquaculture genetics. Aquaculture 2004, 238, 1–37. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Set | Name | Sequence | Tm (°C) | Product Length (bp) | Genes Included |
|---|---|---|---|---|---|
| 1 | MtAst1F MtAst1R | 5′ CAAATCATAAAGATATTGGAAC 3′ 5′ AATGTTGRGGRAAGAATGT 3′ | 49 | 1259 | coxI partial |
| 2 | MtAst2F MtAst2R | 5′ CAGTKGGRGGTTTAACAGGAG 3′ 5′ GTTATCRRGTGATTATTCTGAAC 3′ | 54 | 1255 | coxI (partial) trnL2 coxII trnK (partial) |
| 3 | MtAst3F MtAst3R | 5′ CATTCTTGAACTGTACCTTC 3′ 5′ CTACTAAATGATATGCATGGTG 3′ | 52 | 1201 | coxII (partial) trnK trnD atp8 atp6 coxIII (partial) |
| 4 | MtAst4.2F MtAst4.2R | 5′ GCTGTTGCAATTATTCAGTC 3′ 5′ CCAAAGGTATAAGAAGMGTA 3′ | 50 | 1394 | atp6 (partial) coxIII trnG nad3 (partial) |
| 5 | MtAst5F MtAst5R | 5′ GGCTTCCAACCAAAAGGTC 3′ 5′ AGTWTAACCGCGACTGCTG 3′ | 49 | ~1500–2000 | D-loop |
| 6 | MtAst6F MtAst6R | 5′ GAATTTAACCGCTCAAGAAC 3′ 5′ CCTAACTATTTCTCTTCCGAG 3′ | 52 | 1287 | trnN rrnS trnV rrnL (partial) |
| 7 | MtAst7F MtAst7R | 5′ AGCATCTCATTTACACCGA 3′ 5′ ABTCBAACATGTCTAAGCATC 3′ | 51 | 1424 | rrnL trnL1 nad1 (partial) |
| 8 | MtAst8F MtAst8R | 5′ TACTTTAGGGATAACAGCGTA 3′ 5′ GGTCAAATTCTTTCACTCCT 3′ | 52 | 1502 | rrnL (partial) trnL1 nad1 trnP trnS2 cytb (partial) |
| 9 | MtAst9F MtAst9R | 5′ TACCTCGGTTTCGTTATGA 3′ 5′ ATACYCCYAATATTGAATCAG 3′ | 51 | 1243 | nad1 (partial) trnP trnS2 cytb (partial) |
| 10 | MtAst10F MtAst10R | 5′ GACCTCARGGTAAGACATATC 3′ 5′ TCTCTCCCTAAYTGATTTCC 3′ | 54 | 1463 | cytb (partial) nad6 trnT nad4l |
| 11 | MtAst11F MtAst11R | 5′ GTCCGCTCRCARGGTAATG 3′ 5′ AAAGGAAGYCAATGAAGAC 3′ | 51 | 1339 | nad4l (partial) nad4 trnH (partial) |
| 12 | MtAst12F MtAst12R | 5′ GTCATGGTTTATGTTCATCTG 3′ 5′ ACTCAAAATTAGCTCCRAGC 3′ | 52 | 1285 | nad4l (partial) nad4 trnH nad5 (partial) |
| 13 | MtAst13F MtAst13R | 5′ CGTGTCRGCATTAGTACATTC 3′ 5′ CCCCAAAYCAAGAATTTGAAG 3′ | 54 | 1377 | nad5 (partial) trnF trnI trnM nad2 (partial) |
| 14 | MtAst14F MtAst14R | 5′ CTACATTGAAGCTGTAGAAGAG 3′ 5′ TTTGACAACTTTGAAGGATG 3′ | 51 | 1213 | trnW (partial) nad2 trnW (partial) |
| 15 | MtAst15F MtAst15R | 5′ TCAGCAGGCCTATCATTT 3′ 5′ CAAAAGCATGAGCAGTTACTAC 3′ | 51 | 668 | nad2 (partial) trnW trnC trnY coxI (partial) |
| Position | Gene | Name | Strand | Start Codon | Stop Codon | Anticodon | Gene Length/bp |
|---|---|---|---|---|---|---|---|
| 1–1537 | Cytochrome c oxidase subunit 1 | cox1 | + | ATT | TTA | 1537 | |
| 1539–1601 | tRNA | trnL2 | + | TAA | 63 | ||
| 1602–2288 | Cytochrome c oxidase subunit 2 | cox2 | + | ATG | TAA | 687 | |
| 2290–2353 | tRNA | trnK | + | TTT | 64 | ||
| 2355–2420 | tRNA | trnD | + | GTC | 66 | ||
| 2421–2579 | ATP synthase F0 subunit 8 | atp8 | + | GTG | TAG | 159 | |
| 2573–3247 | ATP synthase F0 subunit 6 | atp6 | + | ATG | TAA | 675 | |
| 3247–4035 | Cytochrome c oxidase subunit 3 | cox3 | + | ATG | TAA | 789 | |
| 4034–4095 | tRNA | trnG | + | TCC | 62 | ||
| 4096–4449 | NADH dehydrogenase subunit 3 | nad3 | + | ATT | TAA | 354 | |
| 4451–4512 | tRNA | trnA | + | TGC | 62 | ||
| 4511–4575 | tRNA | trnR | + | TCG | 65 | ||
| 4576-4645 | tRNA | trnE | + | TTC | 70 | ||
| ~1500–2000 base pairs | Gap | dloop | |||||
| 4858–4926 | tRNA | trnQ | − | TTG | 69 | ||
| 4935–5001 | tRNA | trnS1 | − | TCT | 67 | ||
| 5002–5065 | tRNA | trnN | − | GTT | 64 | ||
| 5143–5956 | rRNA | rrnS | + | 814 | |||
| 5959–6027 | tRNA | trnV | + | TAC | 69 | ||
| 5993–7371 | rRNA (partial) | rrnL | + | 1379 | |||
| 7318–7380 | tRNA | trnL1 | + | TAG | 63 | ||
| 7405–8343 | NADH dehydrogenase subunit 1 | nad1 | + | ATA | TAA | 939 | |
| 8363–8424 | tRNA | trnP | + | TGG | 62 | ||
| 8429–8495 | tRNA | trnS2 | − | TGA | 67 | ||
| 8496–9630 | Cytochrome b | cytb | − | ATG | ATT | 1135 | |
| 9630–10,139 | NADH dehydrogenase subunit 6 | nad6 | − | ATT | TAA | 510 | |
| 10,167–10,232 | tRNA | trnT | − | TGT | 66 | ||
| 10,235–10,528 | NADH dehydrogenase subunit 4 | nad4L | + | ATG | TAA | 294 | |
| 10,525–11,871 | NADH dehydrogenase subunit 4 | nad4 | + | ATA | TAA | 1347 | |
| 11,871–11,932 | tRNA | trnH | + | GTG | 62 | ||
| 11,933–13,666 | NADH dehydrogenase subunit 5 | nad5 | + | ATG | TAA | 1734 | |
| 13,666–13,727 | tRNA | trnF | + | GAA | 62 | ||
| 13,735–13,798 | tRNA | trnI | + | GAT | 64 | ||
| 13,798–13,862 | tRNA | trnM | + | CAT | 65 | ||
| 13,863–14,855 | NADH dehydrogenase subunit 2 | nad2 | + | GTG | TAA | 993 | |
| 14,855–14,921 | tRNA | trnW | + | TCA | 67 | ||
| 14,923–14,986 | tRNA | trnC | − | GCA | 64 | ||
| 14,987–15,050 | tRNA | trnY | − | GTA | 64 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvanou, M.V.; Apostolidis, A.P.; Lattos, A.; Michaelidis, B.; Giantsis, I.A. The Coding Mitogenome of the Freshwater Crayfish Pontastacus leptodactylus (Decapoda:Astacidea:Astacidae) from Lake Vegoritida, Greece and Its Taxonomic Classification. Genes 2023, 14, 494. https://doi.org/10.3390/genes14020494
Alvanou MV, Apostolidis AP, Lattos A, Michaelidis B, Giantsis IA. The Coding Mitogenome of the Freshwater Crayfish Pontastacus leptodactylus (Decapoda:Astacidea:Astacidae) from Lake Vegoritida, Greece and Its Taxonomic Classification. Genes. 2023; 14(2):494. https://doi.org/10.3390/genes14020494
Chicago/Turabian StyleAlvanou, Maria V., Apostolos P. Apostolidis, Athanasios Lattos, Basile Michaelidis, and Ioannis A. Giantsis. 2023. "The Coding Mitogenome of the Freshwater Crayfish Pontastacus leptodactylus (Decapoda:Astacidea:Astacidae) from Lake Vegoritida, Greece and Its Taxonomic Classification" Genes 14, no. 2: 494. https://doi.org/10.3390/genes14020494