

An Ecological Weed Control Strategy in Paddy Fields: Light Interception from Duckweed Mulching
by
Shuhan Xu
1,†, 
Feng Wang
1,†,
Yuling Ding
1,
Wenchao Liu
1,
Yiyu Lan
1,
Qingqing Jia
2,
Peng Sun
1,‡ and
Zhimin Sha
1,3,*,‡ 1
School of Agriculture and Biology, Shanghai Jiao Tong University, Shanghai 200240, China
2
Songjiang Agricultural Technology Extension Center, Shanghai 201600, China
3
Shanghai Yangtze River Delta Eco-Environmental Change and Management Observation and Research Station, Ministry of Science and Technology, Shanghai 200240, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
‡
These authors also contributed equally to this work.
Agronomy 2024, 14(4), 670; https://doi.org/10.3390/agronomy14040670
Submission received: 4 March 2024
/
Revised: 20 March 2024
/
Accepted: 21 March 2024
/
Published: 26 March 2024
(This article belongs to the Special Issue Free from Herbicides: Ecological Weed Control)
Abstract
:Rice–duckweed coculturing as an advanced technique has proven effective for weed control. However, the complex environmental interactions underlying its effectiveness remain unclear. In this study, a controlled pot experiment was conducted to isolate the shading effect of duckweeds (Landoltia punctata and Spirodela polyrhiza) from their intricate environmental interactions with paddy weeds (Cyperus difformis, Eclipta prostrata, and Ammannia auriculata). The results revealed that as the duckweeds grew, there were notable decreases in total weed density (52.41–77.01%) and biomass (42.33–99.23%) while rice biomass remained unaffected. Duckweed covering allowed less than 8.64% of light to pass through and led to decreases in environmental temperature, pH, and dissolved oxygen concentration in the floodwater, but had little impact on inorganic nitrogen levels. Through screening with random forest analysis and structural equation modelling, it was found that weed density and biomass were predominantly influenced by the reduced light transmittance (54.1% and 77.2%, p ≤ 0.001) resulting from duckweed covering while the decline in dissolved oxygen content partly influenced weed density (18%). In conclusion, this study’s results bring a different perspective, confirming the sunlight-shading effect of duckweed as a critical mechanism significantly inhibiting the germination and growth of weeds. These findings provide a scientific basis for the improved application of ecological weed control methods in paddy fields.
1. Introduction
Rice (Oryza sativa), a stable food for more than 60% of the global population, has historically faced significant challenges due to weed infestations dating back to the early days of cultivation [1]. Weeds in paddy fields hinder rice production by competing for vital resources and space, as well as by serving as hosts for pests and diseases [2,3,4]. It is estimated that weeds contribute to a 37% reduction in rice yields globally, with a further 10% yield loss removing mechanical and chemical inputs [5,6]. Herbicide application has traditionally been the primary method of weed control, but prolonged and excessive use has led to the evolution of herbicide-resistant weeds [7,8]. This, in turn, has led to increased reliance on herbicides, raising concerns about soil and water contamination, as well as about the potential toxicity of herbicides to humans and other organisms [9,10,11,12]. The environmental and ecological risks associated with herbicide use highlight the urgent need for alternative, environmentally friendly weed management strategies to ensure sustainable rice production [13,14].
Covering and mulching serve as alternatives to chemical weeding, effectively inhibiting weed germination and growth [15]. While organic residues and synthetic materials are commonly used as covers and mulches [16,17], living mulches, also known as cover crops or intercrops, can also suppress weeds by limiting resource availability or producing allelopathic effects [18,19]. In flooded rice systems, most living covers or mulches, such as Astragalus sinicus and Lolium spp., are typically planted to improve soil fertility during winter fallow periods and are not readily available for weed control during the rice growing seasons [20]. However, a few floating aquatic plants, such as those from the duckweed (Lemnaceae) family and Azolla spp., are well adapted to paddy field ecosystems and can easily form covering layers during the early stages of the rice growing season [21]. The use of duckweeds and Azolla spp. for weed control in paddy fields was practiced and proven effective at least eighteen years ago, providing a sustainable and environmentally friendly approach to weed management in rice cultivation [22].
The Lemnaceae family comprises a wide range of tiny free-floating monocotyledonous plants found in freshwater habitats such as lakes, ponds, ditches, and paddy fields [21]. Known for their rapid growth, prolific reproduction, and strong adaptability to various environmental conditions, these plants play a significant ecological role [23]. According to the findings of a recent study, the introduction of two duckweed species, Landoltia punctata and Spirodela polyrhiza, into paddy fields led to substantial reductions in weed density (90–98%) and biomass (81–100%) throughout the rice growing season [24]. Furthermore, several studies have reported that the introduction of duckweed has promoted rice grain yields [25,26,27]. Due to the myriad benefits and absence of harm associated with duckweed, co-cultivating rice with duckweed has emerged as a promising ecological weed management strategy.
The effect of duckweed on weed control is believed to be mediated by various environmental factors that govern plant growth, particularly competition for nutrients and hydrothermal conditions [28]. Among these factors, lighting conditions play a crucial role in the photosynthesis and biomass accumulation of both duckweed and weeds, especially during the early stages of rice growth [29]. The formation of a duckweed covering layer in paddy fields would restrict the availability of light for weeds [30]. Despite numerous studies confirming the weed-control effect of duckweed, it remains unclear whether the shading effect alone can account for the weed control observed in paddy fields with duckweed, as a dense duckweed mat floating on floodwater induces a range of environmental changes [31]. In this study, a compartmental pot experiment that simulated the conditions of a paddy field during the early stage of the rice growing season was designed and conducted. Simultaneously, screening and modeling techniques were employed to investigate the relationships between duckweed, key environmental factors, and weeds. Our objective was to elucidate the mechanism by which duckweed controls weeds in paddy fields and to assess the relative contributions of the main environmental factors involved.
2. Materials and Methods
2.1. Experimental Conditions
The pot experiment was conducted in an intelligent greenhouse located at Shanghai Jiao Tong University (31°2′22″ N, 121°27′5″ E), situated in Shanghai, China, during August and September 2020 to simulate field conditions. Pot soil was collected from the arable layer (0–20 cm depth) of a paddy field in Qingpu Modern Agriculture Park (30°58′9″ N, 121°0′34″ E), located in Qingpu District, Shanghai, China. The soil retrieved was classified as silty clay loam and possessed the following properties: pH (soil:water 1:2.5, w/v) of 7.19, electrical conductivity (soil:water 1:5, w/v) of 5.10 ms cm−1, total nitrogen (N) content of 1.193 g kg−1, and available phosphorus (P) content of 0.010 g kg−1.
2.2. Plant Materials
The research was carried out with three common weed species in paddy fields, as well as with representative local paddy weeds in Shanghai, including Cyperus difformis, Eclipta prostrata, and Ammannia auriculata. Seeds of Cyperus difformis were collected from Xiangyu Farm (29°49′42″ N, 121°39′52″ E) in Yinzhou District, Ningbo, Zhejiang Province, China. Meanwhile, seeds of Eclipta prostrata and Ammannia auriculata were provided by the Weed Research Group at the Institute of Eco-environmental Protection Research, Shanghai Academy of Agricultural Sciences. The rice cultivar utilized, Qingjiao 307 from Qingpu District, experienced a seedling cultivation process, including soaking, germination, sowing, and emergence, within the greenhouse before transplantation. Furthermore, two species of duckweed, Landoltia punctata and Spirodela polyrhiza, were collected from ponds in Qingpu District and cultivated in Hoagland solution in the laboratory.
2.3. Experimental Design
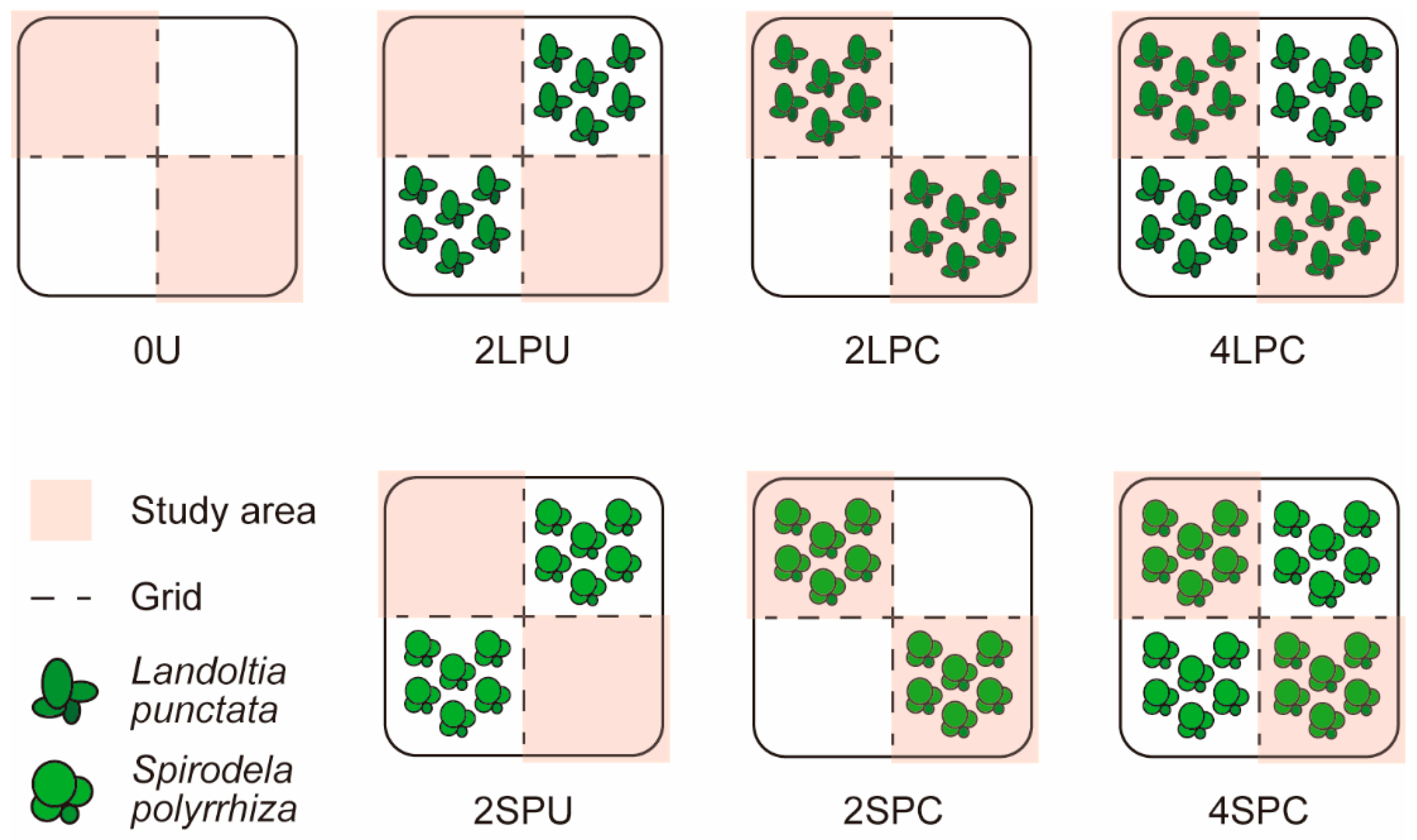
The pot experiment followed a randomized block design consisting of seven treatments with six replications each. The setup, illustrated in Figure 1, involved dividing floodwater in each box-like pot into four equal sections using an 8 cm high cross-shaped grid. Two diagonal portions were then randomly selected as the study area. Duckweed samples were contained within specific sections by the grids, allowing water to flow across them.
Three levels of duckweed coverage were tested: none, two portions occupied, and four portions occupied. In pots with two duckweed-covered sections, the duckweed was positioned diagonally, either in the study area or not in the study area. Since two species of duckweed were studied separately, seven treatments were compared (Table 1), involving no duckweed coverage (0U), non-study areas covered with Landoltia punctata (2LPU), study areas covered with Landoltia punctata (2LPC), the entire pot covered with Landoltia punctata (4LPC), non-study areas covered with Spirodela polyrhiza (2SPU), study areas covered with Spirodela polyrhiza (2SPC), and the entire pot covered with Spirodela polyrhiza (4SPC).
According to our assumptions, shading effect of duckweed would arise in duckweed-covered portions while other environmental effects of duckweed would be apportioned to all portions along with water.
Each pot was square-shaped, measuring 23.2 cm in length and 27.7 cm in height. At the bottom, a layer of 13 cm high soil (8.3 kg) was uniformly mixed with 40 g of organic fertilizer. This was followed by a 4 cm high layer of sterilized soil (2.6 kg, heated at 80 °C for 0.5 h to deactivate weed seeds) mixed with 16 g of organic fertilizer. The organic fertilizer utilized was Nutri Wiz humic acid bio-organic fertilizer (formula 209), containing 4.54% nitrogen, 1.06% phosphorus, and 1.43% potassium. Seeds of 80 Cyperus difformis, 40 Eclipta prostrata, and 40 Ammannia auriculata plants were evenly mixed with 1 cm of sterilized soil (0.64 kg) and then placed on top. Additionally, 25-day-old rice seedlings, growing uniformly, were individually transplanted into the centers of the four portions within each pot. Landoltia punctata and Spirodela polyrhiza were introduced into the corresponding portions, covering approximately 70% of each. The experiment lasted 30 days, during which the positions of the pots within each block were rotated every two days. The water layer depth was maintained at approximately 5 cm throughout the duration of the experiment.
2.4. Sampling and Analyses
Surveys of duckweed growth in duckweed-covered portions and the associated environmental effects in study areas were performed from the first day of the experimental period, occurring every seven days, totaling five times. The duckweeds were sampled in an area of 8 cm2 and then oven-dried at 70 °C for 72 h for measurement of the dry weight. Measurements of light transmittance and water and soil temperature were conducted during 09:00–11:00 a.m. period, with water samples collected simultaneously. Light intensity was measured using a portable photometer. Light transmittance was calculated as the ratio of the light intensity at a depth of 1 cm below the water surface to direct sunlight intensity. Water pH and dissolved oxygen (DO) concentration were measured using a pH meter and DO meter on the day of sampling [32]. We measured the ammonia (NH4+-N) concentration in the water using Nessler’s reagent spectrophotometry, nitrate (NO3−-N) concentration using ultraviolet (UV) spectrophotometry, total N-alkaline concentration using potassium persulfate digestion followed by UV spectrophotometry, and total P concentration using ammonium molybdate spectrophotometry. Weed and rice plants in the study areas were sampled on the thirtieth day of the experimental period. The sampled weed plants were sorted and counted in the laboratory. The plant samples were oven-dried at 70 °C for 72 h and the dry weights were measured.
2.5. Data Analysis
Analysis of variance (ANOVA) followed by Fisher’s least-significant-difference post hoc tests were used to test the significance of differences among treatments (p < 0.05). Random forest algorithm was used to evaluate the relative importance of environmental factors on weed density and biomass. The number of trees grown in random forest was 500 (ntree = 500). Three variables (environmental factors) were considered for each split of the trees (mtry = 3). The relative importance of each variable was measured based on the mean decrease in accuracy in predictions for the out-of-bag samples when a given variable was excluded from the model [33]. The significance of importance metrics for the random forest model was estimated by permuting the response variable (permutations = 100). Structural equation modelling (SEM) was conducted to quantify the direct effects of duckweed on environmental factors and the indirect impacts on weed density and biomass. Based on the results of the random forest analysis, the SEM model only included dominant environmental factors (p < 0.01). A conceptual model was predefined based on experimental results and theoretical analysis. The parameters were estimated using the maximum likelihood method. To align with the assumption of normality, the data were min–max normalized and fourth-root transformed before the establishment of SEM [34]. Normed chi-square (NC), comparative fit index (CFI), and standardized root mean square residual (SRMR) were calculated to evaluate the SEM result. Acceptable fit indices were defined as NC < 3, CFI > 0.90, and SRMR < 0.05 [35].
All statistical analyses were performed using R 4.0.3. Post hoc tests were performed with the R package agricolae 1.3-7. The establishment and permutation test of the random forest model were conducted with the R package rfPermute 2.5.2. SEM was established and evaluated with the R package lavaan 0.6-17.
3. Results
3.1. Weed Density and Biomass
The density of each of the three weed species in the study area is shown in Figure 2. Based on whether the study area was covered or not, the performances of weed density for the treatments could be divided into two groups. The density of each weed species showed no significant difference among the three uncovered treatments (0U, 2LPU, and 2SPU) and, similarly, among the four covered treatments (2LPC, 4LPC, 2SPC, and 4SPC). However, significant differences in weed density between the uncovered treatments and the covered treatments were generally observed. All covered treatments showed lower densities of Cyperus difformis, Eclipta prostrata, and Ammannia auriculata compared to the uncovered treatments. In comparison with the control (0U), the density of Cyperus difformis, Eclipta prostrata, and Ammannia auriculata in covered treatments significantly decreased by 80.83–100%, 43.54–72.11%, and 85.71–100%, respectively. The total density of the three weed species followed the same pattern. Compared to the control, the total density in 2LPC, 4LPC, 2SPC, and 4SPC significantly decreased by 68.43%, 77.01%, 60.54%, and 52.41%, respectively.
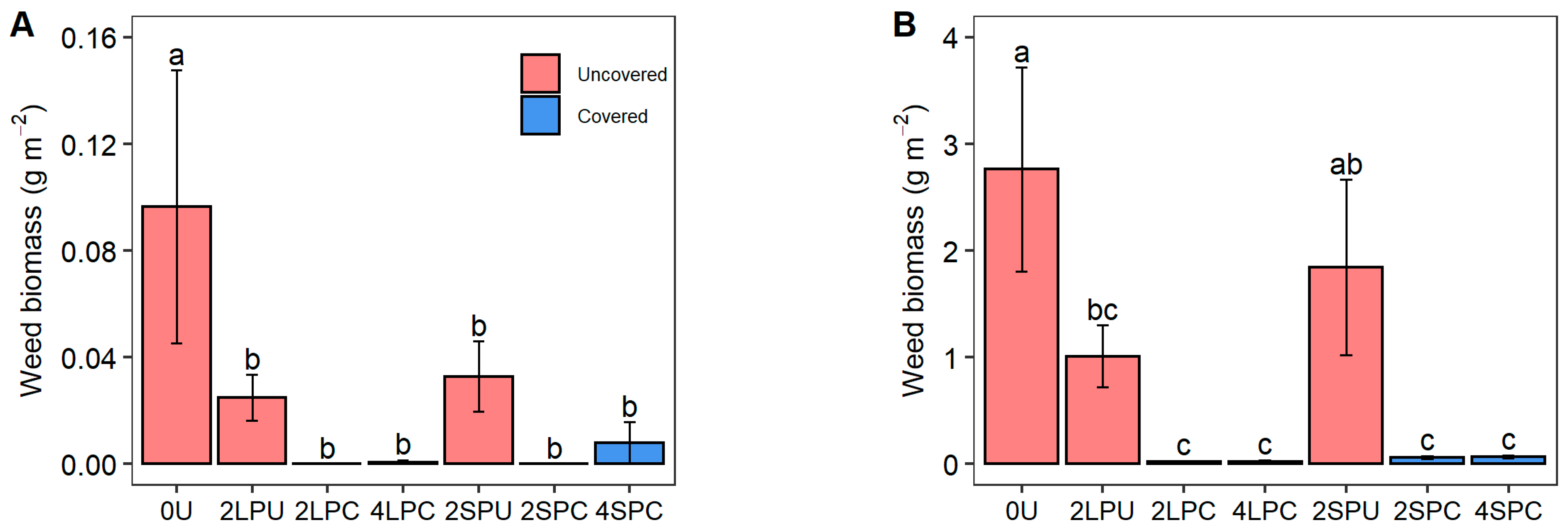
As shown in Figure 3, the biomass of each of the three weed species in the study area was abysmally low in the four covered treatments. Compared to the control, the biomass of Cyperus difformis significantly decreased by 74.16% in 2LPU, 65.98% in 2SPU, and 91.71–100% in the covered treatments. Similarly, the biomass of Eclipta prostrata significantly decreased by 63.42% in 2LPU and 97.53–99.21% in the covered treatments. The biomass of Ammannia auriculata declined to almost zero in the covered treatments. Due to the high proportion of biomass contributed by Eclipta prostrata to the total weed content, compared to the control, the total biomass of the three weed species significantly decreased by 64.11% in 2LPU and 97.36–99.23% in the covered treatments, approaching the biomass of Eclipta prostrata.
3.2. Duckweed and Rice Biomass
The biomass growth trends of the two duckweed species in the duckweed-covered portions are shown in Figure 4. The initial biomass values of Landoltia punctata and Spirodela polyrhiza under corresponding treatments ranged from 13.66 to 15.37 g m−2 and from 15.11 to 20.52 g m−2, respectively. During the first week of the experiment, Landoltia punctata’s biomass increased by 77.47%, 79.15%, and 25.28% in 2LPU, 2LPC, and 4LPC, respectively while Spirodela polyrhiza’s biomass increased by 37.11%, 58.88%, and 37.29% in 2SPU, 2SPC, and 4SPC, respectively. In the subsequent three weeks, Landoltia punctata’s biomass exhibited minimal changes in 2LPU and 2LPC and increased by 59.86% in 4LPC. In the last week, Spirodela polyrhiza’s biomass decreased by 33.45% and 22.90% in 2SPU and 2SPC. The 30-day mean of duckweed biomass was calculated, and the results indicated that there was no significant difference in the overall growth of duckweed in the experimental period among the treatments with duckweed introduction (Table 2).
At the end of the experiment, no significant differences in rice biomass were observed among the different treatments in the study area (Table 2). Rice grew uniformly across all treatments.
3.3. Environmental Factors
We calculated the mean of each environment factor in the study area over the entire experimental period (Figure 5). Naturally, light transmittance was directly affected by whether the study area was covered. Less than 8.64% of light passed through the duckweed mat in the covered treatments while most of the light penetrated the water surface in the uncovered treatments. The uncovered portions (2LPU and 2SPU) in the covered treatments exhibited about 19% lower light transmittance than the control (0U). Soil and water temperature showed minor differences among the different treatments. Compared to the control, soil temperature significantly decreased by 0.24 °C in 4LPC and 4SPC and water temperature significantly decreased by 0.16 °C in 2LPU. Water pH had a conspicuous low point in 4LPC, being 0.49–0.69 lower than in the other treatments. Dissolved oxygen (DO) concentration in the water displayed obvious step-like differences among the different treatments, with the uncovered treatments showing higher DO levels. Compared to the control, DO concentration in the covered treatments significantly decreased by 6.85–9.81%. Additionally, DO concentrations in 2LPU and 2SPU were 3.14% and 4.98% lower than in the control when considering the uncovered treatments. No consistent pattern was observed in the nutrient (NH4+-N, NO3−-N, total N, and total P) concentrations of the water. NH4+-N concentration in the water showed a reversed pattern with pH. The NH4+-N concentration in 4LPC was significantly higher than in the control, 2LPU, and 4SPU. The NO3−-N concentration in the water was lowest in 2LPU, which was significantly lower than in the control (by 10.69%), 4LPC, 2SPU, and 4SPC. Total N concentration in the water in 2LPU and 2LPC was significantly lower than in 4LPC. Total P concentration in the water in 2LPC was significantly lower than in 2SPU.
Based on the random forest algorithm, environment factors were ranked in the order of relative importance in Figure 6. Light transmittance and DO concentration in the water were demonstrated as the predominant predictors of weed density and biomass (p < 0.01). When the response variable was weed density, the increments in the mean squared errors for light transmittance and DO concentration were 15.13% and 11.94%, respectively. When the response variable was weed biomass, the increments in the mean squared errors for light transmittance and DO concentration were 12.73% and 8.71%, respectively.
As assumed, whether a study area would be shaded depended on the duckweed biomass in the study area while changes in other environmental factors in the study area depended on the duckweed biomass in the entire pot: both the study area and the adjacent area. The survey results of the predominant factors partly supported this assumption. According to the survey results and assumption, impact paths among duckweed biomass in the study area (DBS) and adjacent area (DBA), light transmittance, DO concentration in the water, weed density, and biomass were preset (Figure 7A).
The SEM was executed according to the preset impact paths (Figure 7B). The evaluation parameters, especially the CFI, which is stable in the evaluation of small samples, indicated that the model was reliable. As shown in the SEM result, light transmittance was directly affected by duckweed biomass in the study area (p < 0.001), and DO concentration was influenced by duckweed biomass in both the study and adjacent areas (p < 0.001). Light transmittance exerted direct positive effects on weed density (p < 0.001) and biomass (p = 0.001) while the effects of DO concentration were slight. Standardized indirect effects of duckweed biomass on weed were calculated and are shown in Table 3. The total effect of duckweed biomass on weed density was −0.760, with 72.37% of that having been induced by the decrease in light transmittance. Similarly, the total effect of duckweed biomass on weed biomass was −0.775, with light transmittance still being the dominant intermediate.
4. Discussion
4.1. Duckweed Growth and Weed Control
Landoltia punctata and Spirodela polyrhiza are two common native species in local paddy ecosystems. In paddy fields, our previous study and other relevant studies found that the duckweed population increases rapidly during an early stage of the rice growing seasons as sufficient nutrients are released from applied fertilizer and ample light penetrates the sparse rice canopy [24,27]. Our previous study results showed that Landoltia punctata and Spirodela polyrhiza both grew and attained their biomass limits (about 90 g m−2) after approximately three weeks of duckweed introduction into paddy fields. In the present study, the pot experiment tried to simulate the conditions of a paddy field early in the rice growing season [24]. The biomass of both duckweed species increased during the first week, and the duckweeds kept growing or steady in the following two weeks (Figure 4).
However, the duckweed biomass in the pot experiment was limited to about 30 g m−2 or less, which was far less than that in the field experiment. We found two direct reasons. One was a lower initial biomass of duckweeds. Our observations suggested that the thallus of duckweeds cultivated with Hoagland solution in the laboratory was thinner than that of wild duckweeds, which can be contributed to non-seasonal growth and environmental microorganisms [36,37]. The other was a lower growth rate of duckweeds. A low level of NH4+-N concentration could possibly have been a limit factor for duckweed growth in the pot experiment. Ammonia is usually absorbed prior to nitrate by duckweed because NH4+-N is transformed directly to plant protein rather than being assimilated and subsequently reduced as in the case of NO3−-N [38]. According to the pot-experimental results, a considerable part of nitrogen was not in an ionic form, and the proportions of NH4+-N were much lower than those of NO3−-N (Figure 5). We guess that the organic nitrogen lacked a period for releasing from organic fertilizer and mineralizing into inorganic forms as loading soil and fertilizer was quickly followed by introducing duckweed. Light intensity was discounted in the pot experiment likewise. The duckweeds were floating at approximately a 5 cm depth in the pots and partial sunlight was bounced back by the greenhouse roof. In addition, the expansion of duckweed populations was constrained by the narrow portions and windless water layer. The growth of Spirodela polyrhiza seemed to be weaker than the growth of Landoltia punctata; a similar phenomenon was recorded in a previous study where Landoltia punctata plants grown in different media developed close to each other [39].
Both duckweeds showed significant weeding effects despite the relatively low duckweed biomasses (Figure 2 and Figure 3). This was consistent with previous studies showing that duckweed had a control effect on weed germination, growth, and community structure [40,41]. Weed density and biomass were significantly decreased under the duckweed covering, similar to the results of the previous study conducted in paddy fields [24]. Nevertheless, when duckweed appeared in the adjacent area instead of covering it, the weed density was not influenced while the weed biomass was inhibited, which indicated that the presence of duckweed in the nearby waters limited the growth of weed plants rather than affecting the germination of weed seeds. Given the above, it can be preliminarily speculated that the shading effect brought by duckweed covering had contributed the majority of weed growth suppression and determined the colonization of domain weed species while the indirect interaction of duckweed had made a contribution to weed growth suppression.
The biomasses of the 55-day-old rice plants did not show significant differences among all different treatments, which means that neither duckweed coverage nor appearance adversely affected the early growth of rice (Table 2). Some previous studies agreed that duckweed introduction promoted rice yield through nutrient maintenance and delayed release [24,27] while other studies showed that duckweed as a typical weed outcompeted rice seedings in the race for nutrition [42,43]. But in a recent study, duckweed seemed to have more of a facilitating effect on rice grain quality than on yield [44]. Since there was no inhibition effect of duckweed on rice growth in the most competitive growth period (the first month) in this study, it is reasonable to assume that duckweed may have no negative effect on rice yield or even have a potential motivating effect after the duckweed residues release nutrients in the later period.
4.2. Environmental Effects of Duckweed
The formation of dense duckweed layers on the floodwater surface induced a series of changes in the environments of paddy fields. Obviously, the dense duckweed layer obstructed a significant proportion of incident light, leading to a reduction in light transmittance with increasing duckweed biomass. This light shielding can reduce up to 90% of the entry of sunlight (Figure 5). In the early stage of the rice growing season, due to the reduced sunlight reaching the paddy field water and soil, the environmental temperature decreased [45].
The observed variations in floodwater pH and DO concentration in the present study (Figure 5) can be explained by the response of associated microalgae. It was reported in a study that the photosynthesis of microalgae in paddy fields could absorb CO2, thereby increasing the pH of the water and releasing O2 [46,47]. Based on observations during the experiment, microalgae were indeed present in the control and uncovering treatments. According to the literature, microalgae own similar habits with duckweed [46], considering that duckweed have a shading effect; thus, the introduction of duckweed may inhibit the biomass of microalgae and the intensity of photosynthesis, causing reductions in pH and DO concentration [25,48]. The root exudate of duckweed containing organic acids can also lead to water acidification [49]. Although duckweed plants have the ability to secrete O2 from roots [31], the duckweed cover layer restricting gas exchange between the atmosphere and water body can be an inhibitor of DO concentration as well [50].
Substantial studies have indicated that duckweed could significantly reduce nitrogen and phosphorus concentrations in water and thus prevent the loss of these important nutrients [25,27,51]. However, the relevant results showed no clear pattern in the present study (Figure 5). One possible explanation is the relatively small biomass growth of duckweed, ranging from 5 to 10 g m−2, since our previous work showed that the nitrogen and phosphorus contents of duckweed cultivated in slurry were lower than 5% and 1.5%; the disturbance from duckweed assimilation to the environmental element can be weak. In spite of this, a slight fall of nitrate nitrogen and total nitrogen existed when duckweed existed, and the drop was bigger in duckweed covering treatments, which suggested that duckweed had a certain role in these processes. This can be due to the fact that some species of duckweed rhizosphere bacteria specialize in nitrogen removal [52,53]. Landoltia punctata and Spirodela polyrhiza introduction resulted in opposite changes in ammonium nitrogen (Figure 5). To explain, the environmental ammonium nitrogen dynamic is controlled by not only the adsorption of duckweed but also the changes in pH and DO concentration. Generally, as the pH increases, ammonium nitrogen is prone to be converted into ammonia gas escaping from the water while adequate levels of DO concentration contribute to promoting the process of nitrification, converting ammonium nitrogen to nitrate and nitrite nitrogen [54]. It can be found in our results that Landoltia punctata had a greater inhibition effect on pH and DO concentration than another species (Figure 5), and thus, more ammonium nitrogen was able to be retained.
4.3. Weeding Mechanism of Duckweed
The germination and growth of weeds are influenced by various environmental factors, including light intensity and duration, temperature, moisture content, pH, oxygen availability, nutrient content, and others [55,56]. To date, ecological methods of weed control in paddy fields are gradually popularized in the replacement of herbicide application, but the underlying mechanisms of biological interaction and ecosystem response in most methods still remain unknown. In this study investigating rice–duckweed coculturing, we separated the light condition from other environmental chemical factors through an interface-barrier experiment design. Factor screening with random forest analysis (Figure 6) and model validation with a structural equation model (Figure 7) both illustrated that the shading effect of duckweed mainly inhibited weed occurrence by influencing light transmittance and floodwater DO concentration, with sharply reduced light transmittance being the primary mechanism through which duckweed suppressed weed germination and growth (Table 3).
Light stimulation is crucial for breaking dormancy in weeds [57]. Plants possess various light receptors and related light-sensing systems, including photoreceptors such as phytochrome (PHY) and cryptochrome [58]. Studies have indicated that the germination rates of most plant seeds are much lower in darkness compared to well-lit conditions. Rice field weeds such as Lolium perenne, Echinochloa crus-galli, and Portulaca oleracea experience inhibited seed germination in the dark, and germination rates increase with higher intensity [59,60,61]. In addition, light as the energy source for photosynthesis is a critical limiting factor for plant growth. Some paddy weeds rely more on light capture than nutrient uptake [62]. Chlorophyll content, stomatal conductance, biomass accumulation, and flowering in typical weeds have all been found to be strictly restrained by light levels [63]. Recent achievements in weed control including intercropping, rotation, and increasing crop density follow the idea of light regulation, which has been proven to be effective [64,65].
Weeds, like all plants, rely on oxygen for processes such as respiration, nutrient uptake, and energy production. Under the anaerobic environment in a flooded rice-field system, oxygen availability plays an extremely crucial role in determining the growth and development of weeds. Some weeds choose the strategy of cutting down the pathway to obtain oxygen in water to crowd out other weed species [66]. Bioherbicides were used to prevent the gas exchange of weeds for sustainable weed management in a new study [67]. Our results showed that duckweed covering resulted in decreased floodwater DO concentrations and that a change in oxygen availability had a slight inhibitory effect on weed density but almost no influence on weed biomass (Table 3). The elucidation for this can be summarized as the waterlogging resistance of paddy weeds and their higher sensitivity to oxygen while establishing their presence. Weeds that live in flooded paddy fields have evolved effective ways to combat oxygen starvation, with genes responsible for metabolism under anoxia being up-regulated during the germination stage and adaptive strategies being initiated during the vegetative growth stage [68,69].
Although other environmental factors did not perform as the domain-driving factors mediating the inhibitory effect of duckweed covering on weeding in the present study, things may change in a real paddy field. Regardless of the lack of distinct fluctuation of nutrient content in this study, a broader natural paddy field can offer a suitable growing environment for duckweeds, which may operate to the disadvantage of weed growth through nutrient competition. It is also worth considering that a heavy duckweed layer in a paddy field can exert gravity pressure so that weeds are physically suppressed. Moreover, in view of the common phenomenon wherein some plants may produce allelopathic substances that inhibit the growth of other plants, duckweed may also be one of these plants. Contemporary researchers have discovered that duckweeds contain underlying allelopathic compounds such as phenolic acids and ionone that act against both aquatic and terrestrial plants [70,71,72], but there is no clear evidence to support their contribution in weeding.
While the specific physicochemical mechanisms underlying weed control by duckweed in paddy fields remain uncertain, further investigation is essential. Future studies should explore alternative pathways that may contribute, examine the individual contributions of different mechanisms, and assess whether the inhibitory effect of duckweeds on weeds persists throughout the remaining rice growth period and across different paddy systems.
5. Conclusions
This study validated the effectiveness of using two duckweed species, Lemna punctata and Spirodela polyrhiza, as natural coverings for weed control without negatively impacting rice growth during the early stages of the rice growing season. The presence of duckweed covering induced significant attenuation in light transmittance, temperature, pH, and dissolved oxygen concentration in the paddy environment. Duckweed played a crucial role in inhibiting weed development, primarily through two main factors: reducing light transmittance and altering dissolved oxygen concentration in the floodwater. The shading effect of duckweed emerged as a pivotal mechanism, substantially limiting light availability and inhibiting the germination and growth of weeds. These findings highlight the importance of light regulation by duckweed, providing an effective and sustainable approach to weed management.
Author Contributions
Conceptualization, F.W.; Methodology, Y.D.; Software, W.L.; Formal analysis, Y.L.; Investigation, Y.D., W.L. and Y.L.; Resources, Q.J.; Data curation, Q.J.; Writing—original draft, S.X.; Writing—review & editing, F.W. and P.S.; Visualization, S.X.; Supervision, P.S. and Z.S.; Project administration, Z.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Agriculture Research System of Shanghai, China (202203).
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mahajan, G.; Chauhan, B.S.; Kumar, V. Integrated weed management in rice. In Recent Advances in Weed Management; Springer: New York, NY, USA, 2014; pp. 125–153. [Google Scholar]
- Dong, F.; Xu, J.; Zhang, X.; Wang, S.; Xing, Y.; Mokoena, M.P.; Olaniran, A.O.; Shi, J. Gramineous weeds near paddy fields are alternative hosts for the Fusarium graminearum species complex that causes fusarium head blight in rice. Plant Pathol. 2020, 69, 433–441. [Google Scholar] [CrossRef]
- Shimada, N.; Sugiura, S. Indirect effects of weeds on rice plants via shared heteropteran herbivores. J. Appl. Èntomol. 2021, 145, 117–124. [Google Scholar] [CrossRef]
- Zhang, Z.; Gu, T.; Zhao, B.; Yang, X.; Peng, Q.; Li, Y.; Bai, L. Effects of common Echinochloa varieties on grain yield and grain quality of rice. Field Crop. Res. 2017, 203, 163–172. [Google Scholar] [CrossRef]
- Matloob, A.; Khaliq, A.; Chauhan, B.S. Weeds of direct-seeded rice in Asia: Problems and opportunities. Adv. Agron. 2015, 130, 291–336. [Google Scholar]
- Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Benbrook, C.M. Trends in glyphosate herbicide use in the United States and globally. Environ. Sci. Eur. 2016, 28, 3. [Google Scholar] [CrossRef] [PubMed]
- Owen, M.D.; Zelaya, I.A. Herbicide-resistant crops and weed resistance to herbicides. Pest Manag. Sci. Former. Pestic. Sci. 2005, 61, 301–311. [Google Scholar] [CrossRef]
- Peterson, M.A.; Collavo, A.; Ovejero, R.; Shivrain, V.; Walsh, M.J. The challenge of herbicide resistance around the world: A current summary. Pest Manag. Sci. 2018, 74, 2246–2259. [Google Scholar] [CrossRef]
- Sparks, T.C.; Hunter, J.E.; Lorsbach, B.A.; Hanger, G.; Gast, R.E.; Kemmitt, G.; Bryant, R.J. Crop protection discovery: Is being the first best? J. Agric. Food Chem. 2018, 66, 10337–10346. [Google Scholar] [CrossRef]
- Guzzella, L.; Pozzoni, F.; Giuliano, G. Herbicide contamination of surficial groundwater in Northern Italy. Environ. Pollut. 2006, 142, 344–353. [Google Scholar] [CrossRef]
- Kniss, A.R. Long-term trends in the intensity and relative toxicity of herbicide use. Nat. Commun. 2017, 8, 14865. [Google Scholar] [CrossRef]
- Zimdahl, R.L. Fundamentals of Weed Science; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Dass, A.; Shekhawat, K.; Choudhary, A.K.; Sepat, S.; Rathore, S.S.; Mahajan, G.; Chauhan, B.S. Weed management in rice using crop competition-a review. Crop Prot. 2017, 95, 45–52. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, J.; DiTommaso, A.; Zhang, C.; Zheng, G.; Liang, W.; Islam, F.; Yang, C.; Chen, X.; Zhou, W. Weed research status, challenges, and opportunities in China. Crop Prot. 2020, 134, 104449. [Google Scholar] [CrossRef]
- Mohammed, U.; Aimrun, W.; Amin, M.S.M.; Khalina, A.; Zubairu, U.B. Influence of soil cover on moisture content and weed suppression under system of rice intensification (SRI). Paddy Water Environ. 2016, 14, 159–167. [Google Scholar] [CrossRef]
- Zhu, J.; Liang, W.; Yang, S.; Wang, H.; Shi, C.; Wang, S.; Zhou, W.; Lu, Q.; Islam, F.; Xu, W.; et al. Safety of Oilseed Rape Straw Mulch of Different Lengths to Rice and Its Suppressive Effects on Weeds. Agronomy 2020, 10, 201. [Google Scholar] [CrossRef]
- Jabran, K.; Mahajan, G.; Sardana, V.; Chauhan, B.S. Allelopathy for weed control in agricultural systems. Crop Prot. 2015, 72, 57–65. [Google Scholar] [CrossRef]
- MacLaren, C.; Storkey, J.; Menegat, A.; Metcalfe, H.; Dehnen-Schmutz, K. An ecological future for weed science to sustain crop production and the environment. A review. Agron. Sustain. Dev. 2020, 40, 24. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, L.; Wei, J.; Xu, H.; Tang, Q.; Tang, J. Integrating cover crops with chicken grazing to improve soil nitrogen in rice fields and increase economic output. Sci. Total Environ. 2020, 713, 135218. [Google Scholar] [CrossRef]
- Hejný, S. Ellias Landolt The family of Lemnaceae—A monographic study—Vol.1 biosystematic investigations in the family of Duckweeds (Lemnaceae) (vol. 2). Folia Geobot. Phytotax. 1993, 28, 50. [Google Scholar] [CrossRef]
- Biswas, M.; Parveen, S.; Shimozawa, H.; Nakagoshi, N. Effects of Azolla species on weed emergence in a rice paddy ecosystem. Weed Biol. Manag. 2005, 5, 176–183. [Google Scholar] [CrossRef]
- Xu, J.; Shen, Y.; Zheng, Y.; Smith, G.; Sun, X.S.; Wang, D.; Zhao, Y.; Zhang, W.; Li, Y. Duckweed (Lemnaceae) for potentially nutritious human food: A review. Food Rev. Int. 2023, 39, 3620–3634. [Google Scholar] [CrossRef]
- Wang, F.; Wang, S.; Xu, S.; Shen, J.; Cao, L.; Sha, Z.; Chu, Q. A non-chemical weed control strategy, introducing duckweed into the paddy field. Pest Manag. Sci. 2022, 78, 3654–3663. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liang, X.; Lian, Y.; Xu, L.; Chen, Y. Reduction of Ammonia Volatilization from Urea by a Floating Duckweed in Flooded Rice Fields. Soil Sci. Soc. Am. J. 2009, 73, 1890–1895. [Google Scholar] [CrossRef]
- Sun, H.; Dan, A.; Feng, Y.; Vithanage, M.; Mandal, S.; Shaheen, S.M.; Rinklebe, J.; Shi, W.; Wang, H. Floating duckweed mitigated ammonia volatilization and increased grain yield and nitrogen use efficiency of rice in biochar amended paddy soils. Chemosphere 2019, 237, 124532. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Zhang, M.; Tian, Y.; Zhao, M.; Zhang, B.; Zhao, M.; Zeng, K.; Yin, B. Duckweed (Spirodela polyrhiza) as green manure for increasing yield and reducing nitrogen loss in rice production. Field Crop. Res. 2017, 214, 273–282. [Google Scholar] [CrossRef]
- Ekperusi, A.O.; Sikoki, F.D.; Nwachukwu, E.O. Application of common duckweed (Lemna minor) in phytoremediation of chemicals in the environment: State and future perspective. Chemosphere 2019, 223, 285–309. [Google Scholar] [CrossRef] [PubMed]
- Tabou, T.T.; Baya, D.T.; Eyul’anki, D.M.; Vasel, J.L. Monitoring the influence of light intensity on the growth and mortality of duckweed (Lemna minor) through digital images processing. Biotechnol. Agron. Soc. Environ. 2014, 18, 37–48. [Google Scholar]
- McLay, C.L. The Distribution of Duckweed Lemna perpusilla in a Small Southern California Lake: An Experimental Approach. Ecology 1974, 55, 262–276. [Google Scholar] [CrossRef]
- Körner, S.; Vermaat, J.E.; Veenstra, S. The capacity of duckweed to treat wastewater: Ecological considerations for a sound design. J. Environ. Qual. 2003, 32, 1583–1590. [Google Scholar] [CrossRef]
- Sha, Z.; Chen, Z.; Feng, Y.; Xue, L.; Yang, L.; Cao, L.; Chu, Q. Minerals loaded with oxygen nanobubbles mitigate arsenic translocation from paddy soils to rice. J. Hazard. Mater. 2020, 398, 122818. [Google Scholar] [CrossRef]
- Hastie, T.; Tibshirani, R.; Friedman, J.H.; Friedman, J.H. The Elements of Statistical Learning: Data Mining, Inference, and Prediction; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Bowen, N.K.; Guo, S. Structural Equation Modeling; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Ishizawa, H.; Kuroda, M.; Morikawa, M.; Ike, M. Evaluation of environmental bacterial communities as a factor affecting the growth of duckweed Lemna minor. Biotechnol. Biofuels 2017, 10, 62. [Google Scholar] [CrossRef] [PubMed]
- Van Dyck, I.; Vanhoudt, N.; i Batlle, J.V.; Horemans, N.; Nauts, R.; Van Gompel, A.; Claesen, J.; Vangronsveld, J. Effects of environmental parameters on Lemna minor growth: An integrated experimental and modelling approach. J. Environ. Manag. 2021, 300, 113705. [Google Scholar] [CrossRef] [PubMed]
- Elshafai, S.; Elgohary, F.; Nasr, F.A.; Petervandersteen, N.; Gijzen, H.J. Nutrient recovery from domestic wastewater using a UASB-duckweed ponds system. Bioresour. Technol. 2007, 98, 798–807. [Google Scholar] [CrossRef]
- Stadtlander, T.; Förster, S.; Rosskothen, D.; Leiber, F. Slurry-grown duckweed (Spirodela polyrhiza) as a means to recycle nitrogen into feed for rainbow trout fry. J. Clean. Prod. 2019, 228, 86–93. [Google Scholar] [CrossRef]
- Dai, W.F.; Zhai, C.H. Effect of duckweeds and rice chaff on weeds control in paddy field. Mod. Agric. Sci. Technol. 2020, 1, 97–98. [Google Scholar]
- Wang, F.; Lai, Y.; Tang, Z.; Zheng, M.; Shi, J.; Gu, M.; Shen, J.; Cao, L.; Sha, Z. Effects of duckweed mulching on composition and diversity of weed communities in paddy fields. Chin. J. Eco-Agric. 2021, 29, 672–682. [Google Scholar]
- Jing, L.; Li, F.; Zhao, Y.; Wang, X.; Zhao, F.; Lai, S.; Sun, X.; Wang, Y.; Yang, L. Research progress on the carbon and nitrogen sink of duckweed growing in paddy and its effects on rice yield. Sci. Agric. Sin. 2023, 56, 4717–4728. [Google Scholar]
- Wersal, R.M.; Turnage, G. Using contact herbicides for control of duckweed and watermeal with implications for management. J. Aquat. Plant Manag. 2021, 59, 40–45. [Google Scholar]
- Luo, J.; Hu, S.; Li, T.; He, F.; Tian, C.; Han, Y.; Mao, Y.; Jing, L.; Yang, L.; Wang, Y. A Preliminary Study of the Impacts of Duckweed Coverage during Rice Growth on Grain Yield and Quality. Plants 2023, 13, 57. [Google Scholar] [CrossRef] [PubMed]
- Chamara, B.; Marambe, B.; Chauhan, B.S. Management of Cleome rutidosperma DC. using high crop density in dry-seeded rice. Crop Prot. 2017, 95, 120–128. [Google Scholar] [CrossRef]
- Li, X.; Wu, S.; Yang, C.; Zeng, G. Microalgal and duckweed based constructed wetlands for swine wastewater treatment: A review. Bioresour. Technol. 2020, 318, 123858. [Google Scholar] [CrossRef] [PubMed]
- Thind, H.; Rowell, D. Effect of green manure and floodwater algae on diurnal fluctuations of floodwater pH and depth of aerobic soil layer under lowland rice conditions. Plant Soil 1997, 192, 161–165. [Google Scholar] [CrossRef]
- Parr, L.B.; Perkins, R.G.; Mason, C.F. Reduction in photosynthetic efficiency of Cladophora glomerata, induced by overlying canopies of Lemna spp. Water Res. 2002, 36, 1735–1742. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhou, Y.; Nakai, S.; Hosomi, M.; Zhang, H.; Kronzucker, H.J.; Shi, W. Stimulation of nitrogen removal in the rhizosphere of aquatic duckweed by root exudate components. Planta 2014, 239, 591–603. [Google Scholar] [CrossRef] [PubMed]
- Wollenberg, J.L.; Peters, S.C. Diminished mercury emission from waters with duckweed cover. J. Geophys. Res. Biogeosci. 2009, 114, G2. [Google Scholar] [CrossRef]
- Liu, C.; Dai, Z.; Sun, H. Potential of duckweed (Lemna minor) for removal of nitrogen and phosphorus from water under salt stress. J. Environ. Manag. 2017, 187, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Kronzucker, H.J.; Shi, W. Stigmasterol root exudation arising from Pseudomonas inoculation of the duckweed rhizosphere enhances nitrogen removal from polluted waters. Environ. Pollut. 2021, 287, 117587. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Yin, Z.; Xia, D.; Zhao, Q.; Kang, Y. Combination of heterotrophic nitrifying bacterium and duckweed (Lemna gibba L.) enhances ammonium nitrogen removal efficiency in aquaculture water via mutual growth promotion. J. Gen. Appl. Microbiol. 2019, 65, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Ying, J.; Li, X.; Wang, N.; Lan, Z.; He, J.; Bai, Y. Contrasting effects of nitrogen forms and soil pH on ammonia oxidizing microorganisms and their responses to long-term nitrogen fertilization in a typical steppe ecosystem. Soil Biol. Biochem. 2017, 107, 10–18. [Google Scholar] [CrossRef]
- Nozoe, T.; Tazawa, J.; Uchino, A.; Miura, S. Promotive effect of soil solution on germination of Monochoria vaginalis under paddy conditions. Soil Sci. Plant Nutr. 2018, 64, 396–405. [Google Scholar] [CrossRef]
- Raj, S.K.; Syriac, E.K. Weed management in direct seeded rice: A review. Agric. Rev. 2017, 38, 41–50. [Google Scholar] [CrossRef]
- Benech-Arnold, R.L.; Sánchez, R.A.; Forcella, F.; Kruk, B.C.; Ghersa, C.M. Environmental control of dormancy in weed seed banks in soil. Field Crop. Res. 2000, 67, 105–122. [Google Scholar] [CrossRef]
- Kami, C.; Lorrain, S.; Hornitschek, P.; Fankhauser, C. Light-Regulated Plant Growth and Development. Curr. Top. Dev. Biol. 2010, 91, 29–66. [Google Scholar] [PubMed]
- Chauhan, B.S.; Johnson, D.E. Germination Ecology of Chinese Sprangletop (Leptochloa chinensis) in the Philippines. Weed Sci. 2008, 56, 820–825. [Google Scholar] [CrossRef]
- Derakhshan, A.; Gherekhloo, J. Factors affecting Cyperus difformis seed germination and seedling emergence. Planta Daninha 2013, 31, 823–832. [Google Scholar] [CrossRef]
- Sumudunie, H.; Jayasuriya, K. Seed dormancy and germination of seven rice field weeds from Sri Lanka. Taiwania 2019, 64, 221–230. [Google Scholar] [CrossRef]
- Caton, B.; Foin, T.; Hill, J. Phenotypic plasticity of Ammannia spp. in competition with rice. Weed Res. 1997, 37, 33–38. [Google Scholar] [CrossRef]
- Yasin, M.; Rosenqvist, E.; Jensen, S.M.; Andreasen, C. The importance of reduced light intensity on the growth and development of six weed species. Weed Res. 2019, 59, 130–144. [Google Scholar] [CrossRef]
- Batlla, D.; Benech-Arnold, R.L. Weed seed germination and the light environment: Implications for weed management. Weed Biol. Manag. 2014, 14, 77–87. [Google Scholar] [CrossRef]
- Sharma, G.; Shrestha, S.; Kunwar, S.; Tseng, T.-M. Crop Diversification for Improved Weed Management: A Review. Agriculture 2021, 11, 461. [Google Scholar] [CrossRef]
- Jawed, A.; Kar, P.; Verma, R.; Shukla, K.; Hemanth, P.; Thakur, V.K.; Pandey, L.M.; Gupta, R.K. Integration of biological control with engineered heterojunction nano-photocatalysts for sustainable and effective management of water hyacinth weed. J. Environ. Chem. Eng. 2022, 10, 106976. [Google Scholar] [CrossRef]
- Hasan, M.; Ahmad-Hamdani, M.S.; Rosli, A.M.; Hamdan, H. Bioherbicides: An Eco-Friendly Tool for Sustainable Weed Management. Plants 2021, 10, 1212. [Google Scholar] [CrossRef] [PubMed]
- Kaspary, T.E.; Roma-Burgos, N.; Merotto, A. Snorkeling Strategy: Tolerance to Flooding in Rice and Potential Application for Weed Management. Genes 2020, 11, 975. [Google Scholar] [CrossRef] [PubMed]
- Turra, G.M.; Cutti, L.; Angonese, P.S.; Sulzbach, E.; Mariot, C.H.P.; Markus, C.; Merotto, A., Jr. Variability to flooding tolerance in barnyardgrass and early flooding benefits on weed management and rice grain yield. Field Crop. Res. 2023, 300, 108999. [Google Scholar] [CrossRef]
- Bich, T.T.N.; Kato-Noguchi, H. Allelopathic potential of two aquatic plants, duckweed (Lemna minor L.) and water lettuce (Pistia stratiotes L.), on terrestrial plant species. Aquat. Bot. 2012, 103, 30–36. [Google Scholar] [CrossRef]
- Bich, T.T.; Ohno, O.; Suenaga, K.; Kato-Noguchi, H. Isolation and identification of an allelopathic substance from duckweed (Lemna minor L.). Allelopath. J. 2013, 32, 213. [Google Scholar]
- Gostyńska, J.; Pankiewicz, R.; Romanowska-Duda, Z.; Messyasz, B. Overview of allelopathic potential of Lemna minor L. obtained from a shallow eutrophic lake. Molecules 2022, 27, 3428. [Google Scholar] [CrossRef]
Figure 1.
Schematic diagram of pot experiment. Rice plants and weeds have been omitted in the diagram.
Figure 1.
Schematic diagram of pot experiment. Rice plants and weeds have been omitted in the diagram.

Figure 2.
Density of (A) Cyperus difformis, (B) Eclipta prostrata, (C) Ammannia auriculata, and (D) total weed species in study area under different treatments. Different lowercase letters indicate significant differences among treatments (p < 0.05).
Figure 2.
Density of (A) Cyperus difformis, (B) Eclipta prostrata, (C) Ammannia auriculata, and (D) total weed species in study area under different treatments. Different lowercase letters indicate significant differences among treatments (p < 0.05).


Figure 3.
Biomass of (A) Cyperus difformis, (B) Eclipta prostrata, (C) Ammannia auriculata, and (D) total weed species in study area under different treatments. Different lowercase letters indicate significant differences among treatments (p < 0.05).
Figure 3.
Biomass of (A) Cyperus difformis, (B) Eclipta prostrata, (C) Ammannia auriculata, and (D) total weed species in study area under different treatments. Different lowercase letters indicate significant differences among treatments (p < 0.05).


Figure 4.
Growth trends of (A) Landoltia punctata and (B) Spirodela polyrhiza biomass in duckweed-covered portions under different treatments.
Figure 4.
Growth trends of (A) Landoltia punctata and (B) Spirodela polyrhiza biomass in duckweed-covered portions under different treatments.

Figure 5.
Thirty-day mean of environment factors, including (A) light transmittance, (B) soil temperature, (C) water temperature, (D) pH, (E) DO concentration, (F) NH4+-N concentration, (G) NO3−-N concentration, (H) total N concentration, and (I) total P concentration, in study area under different treatments. Different letters indicate that there were significant differences among treatments (p < 0.05).
Figure 5.
Thirty-day mean of environment factors, including (A) light transmittance, (B) soil temperature, (C) water temperature, (D) pH, (E) DO concentration, (F) NH4+-N concentration, (G) NO3−-N concentration, (H) total N concentration, and (I) total P concentration, in study area under different treatments. Different letters indicate that there were significant differences among treatments (p < 0.05).


Figure 6.
Importance ranking of environment factors affecting (A) weed density and (B) biomass. LT: light transmittance; DO: dissolved oxygen; pH: water pH; TP: water total phosphorus content; NN: water NO3−-N content; ST: soil temperature; TN: water total nitrogen content; AN: water NH4+-N content; WT: water temperature. Symbol ‘**’: significant at p < 0.01.
Figure 6.
Importance ranking of environment factors affecting (A) weed density and (B) biomass. LT: light transmittance; DO: dissolved oxygen; pH: water pH; TP: water total phosphorus content; NN: water NO3−-N content; ST: soil temperature; TN: water total nitrogen content; AN: water NH4+-N content; WT: water temperature. Symbol ‘**’: significant at p < 0.01.

Figure 7.
(A) Preset impact paths and (B) SEM result. DBS: duckweed biomass in study area; DBA: duckweed biomass in adjacent area; LT: light transmittance; DO: dissolved oxygen.
Figure 7.
(A) Preset impact paths and (B) SEM result. DBS: duckweed biomass in study area; DBA: duckweed biomass in adjacent area; LT: light transmittance; DO: dissolved oxygen.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Treatment settings.
| Treatments | Duckweed Species | Number of Quarters Duckweed Covered | Whether Duckweed Covered the Study Area |
|---|---|---|---|
| 0U | - | - | - |
| 2LPU | Landoltia punctata | 2 | No |
| 2LPC | Landoltia punctata | 2 | Yes |
| 4LPC | Landoltia punctata | 4 | Yes |
| 2SPU | Spirodela polyrhiza | 2 | No |
| 2SPC | Spirodela polyrhiza | 2 | Yes |
| 4SPC | Spirodela polyrhiza | 4 | Yes |
Table 2.
Rice biomass in study area and 30-day mean of duckweed biomass in duckweed-covered portions under different treatments. Same lowercase letters indicate no significant differences among treatments (p > 0.05).
Table 2.
Rice biomass in study area and 30-day mean of duckweed biomass in duckweed-covered portions under different treatments. Same lowercase letters indicate no significant differences among treatments (p > 0.05).
| Treatments | 30-Day Mean of Duckweed Biomass in Duckweed-Covered Portions (g m−2) | Rice Biomass in Study Area (g m−2) |
|---|---|---|
| 0U | - | 48.97 ± 6.97 a |
| 2LPU | 21.81 ± 0.69 a | 54.56 ± 2.10 a |
| 2LPC | 22.28 ± 0.74 a | 47.02 ± 8.62 a |
| 4LPC | 22.05 ± 1.05 a | 53.84 ± 6.32 a |
| 2SPU | 23.89 ± 1.18 a | 55.13 ± 9.57 a |
| 2SPC | 21.17 ± 1.03 a | 42.09 ± 3.10 a |
| 4SPC | 22.15 ± 1.72 a | 47.63 ± 7.76 a |
Table 3.
Standardized indirect and total effects of duckweed biomass on weed density and biomass.
| Predictors | Mediators | Responses | Indirect Effects | Total Effects |
|---|---|---|---|---|
| DBS † | LT ‡ | Weed density | −0.523 | −0.669 |
| DO ‡ | −0.146 | |||
| DBA † | LT ‡ | −0.027 | −0.091 | |
| DO ‡ | −0.064 | |||
| DBS † | LT ‡ | Weed biomass | −0.746 | −0.739 |
| DO ‡ | 0.007 | |||
| DBA † | LT ‡ | −0.039 | −0.036 | |
| DO ‡ | 0.003 |
† DBS: duckweed biomass in study area; DBA: duckweed biomass in adjacent area. ‡ LT: light transmittance; DO: dissolved oxygen.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xu, S.; Wang, F.; Ding, Y.; Liu, W.; Lan, Y.; Jia, Q.; Sun, P.; Sha, Z. An Ecological Weed Control Strategy in Paddy Fields: Light Interception from Duckweed Mulching. Agronomy 2024, 14, 670. https://doi.org/10.3390/agronomy14040670
AMA Style
Xu S, Wang F, Ding Y, Liu W, Lan Y, Jia Q, Sun P, Sha Z. An Ecological Weed Control Strategy in Paddy Fields: Light Interception from Duckweed Mulching. Agronomy. 2024; 14(4):670. https://doi.org/10.3390/agronomy14040670
Chicago/Turabian StyleXu, Shuhan, Feng Wang, Yuling Ding, Wenchao Liu, Yiyu Lan, Qingqing Jia, Peng Sun, and Zhimin Sha. 2024. "An Ecological Weed Control Strategy in Paddy Fields: Light Interception from Duckweed Mulching" Agronomy 14, no. 4: 670. https://doi.org/10.3390/agronomy14040670
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.