Genetic Structure and Molecular Identities of 46 Apple Landraces (Malus Mill.) in China

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Extraction and SSR Amplification
2.3. Data Analysis
3. Results
3.1. Characterization of SSR Loci
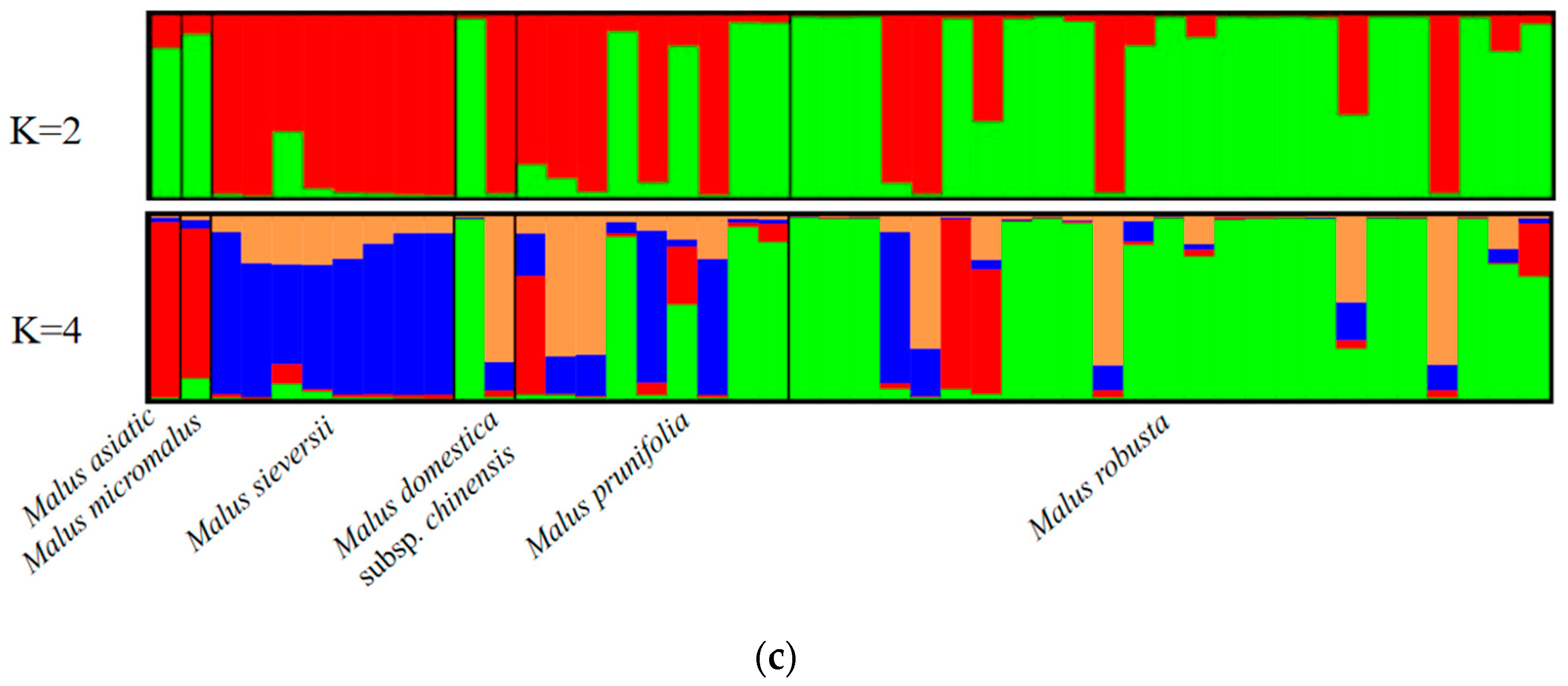
3.2. Genetic Structures of Six Populations
3.3. Selection of the Core Group of SSR Primers and Construction of TP-M13-SSR Fingerprints
3.4. Encoding Molecular Identities
4. Discussion
4.1. The System of Constructing Molecular Identities Based on Fingerprints
4.2. Genetic Diversity of Malus
4.3. Genetic Structure of Malus
4.4. The Construction of Molecular Identities Using Tp-M13-SSR
4.5. The Encoding of Molecular Identities of Malus
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Y.N. Resarches of Malus Mill. Germplasm Resources; China Agriculture Press: Beijing, China, 2001; pp. 125–141. [Google Scholar]
- Dong, Y.C.; Liu, X. Crops and Their Wild Relatives in China; China Agriculture Press: Beijing, China, 2006; pp. 55–84. [Google Scholar]
- Li, Y.N.; Li, X.L. Research on peroxidase isozyme in Malus Mill. J. Southwest Agric. Univ. 1995, 5, 371–377. [Google Scholar]
- Shi, Y.S.; Luo, J.X. Comparison of peroxidese zymogrames of ‘Delicious’ apple cultivers. J. Tianjin Agric. Coll. 1995, 1, 11–14. [Google Scholar]
- Wang, A.D.; Li, T.Z.; Xu, X.F.; Han, Z.H. Analysis for apple cultivars. Acta Hortic. Sin. 2005, 5, 875–877. [Google Scholar]
- Zhu, J.; Wang, T.; Zhao, Y.J.; Zhang, W.; Li, G.C. Identification of apple varieties with AFLP molecular markers. Acta Hortic. Sin. 2007, 2, 102–106. [Google Scholar]
- Gao, Y.; Liu, F.Z.; Cao, Y.F.; Wang, K. SSR analysis of genetic relationship for apple germplasm resources. J. Fruit Sci. 2007, 2, 129–134. [Google Scholar]
- Gao, Y.; Wang, K.; Tian, L.M.; Cao, Y.; Liu, F.Z. Identification of apple cultivars by TP-M13-SSR technique. J. Fruit Sci. 2010, 5, 833–837. [Google Scholar]
- Gao, Y.; Wang, K.; Tian, L.M.; Cao, Y.F.; Liu, F.Z. TP-M13-SSR technique and its applications in analysis of genetic diversity for apple germplasm resources. J. Plant Genet. Resour. 2011, 2, 228–233. [Google Scholar]
- Cao, Y.F.; Tian, L.M.; Gao, Y.; Liu, F.Z. Genetic diversity of cultivated and wild Ussurian Pear (Pyrus ussuriensis Maxim.) in China evaluated with M13-tailed SSR markers. Genet. Resour. Crop Evol. 2012, 59, 9–17. [Google Scholar] [CrossRef]
- Chen, C.W.; Cao, K.; Wang, L.R.; Zhu, G.R.; Fang, W.C. Molecular ID establishment of main China peach varieties and peach related species. Sci. Agric. Sin. 2011, 10, 2081–2093. [Google Scholar]
- Xu, L.F.; Ge, L.; Yuan, S.X.; Ren, J.F.; Yuan, Y.Y.; Li, Y.Y.; Liu, C.; Ming, J. Using the fluorescent labeled SSR markers to establish molecular identity of Lily germplasms. Acta Hortic. Sin. 2014, 10, 2055–2064. [Google Scholar]
- Ai, C.X.; Zhang, L.S.; Wei, H.R.; Yuan, K.J.; Jin, S.N.; Liu, Q.Z. Construction of molecular fingerprinting database for sweet cherry using SSR markers. Chin. Agric. Sci. Bull. 2007, 5, 55–58. [Google Scholar]
- Ohtsubok, K.; Nakamuras, S. Cultivar identification of rice (Oryza sativa L.) by polymerase chain reaction method and its application to processed rice products. J. Agr. Food Chem. 2007, 55, 1501–1509. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.W.; Tian, D.G.; Xu, Y.; Wang, F. Constructing SSR molecular database for identity system of hybrid rice parents. Fujian J. Agric. Sci. 2011, 2, 148–152. [Google Scholar]
- Pan, Y.B. Databasing molecular identities of sugarcane (Saccharum spp.) clones constructed with microsatellite (SSR) DNA markers. Am. J. Plant Sci. 2010, 1, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.L.; Ma, L.; Chen, X.K.; Ying, X.M.; Cai, Q.; Liu, J.Y.; Wu, C.W. Establishment of DNA fingerprint ID in sugarcane cultivars in Yunnan. Acta Agron. Sin. 2010, 2, 202–210. [Google Scholar]
- Yang, Y.; Liu, Z.; Zhao, Y.; Liang, G.Q. Construction of DNA fingerprints for tea cultivars originated from Hunan province. J. Tea Sci. 2010, 5, 367–373. [Google Scholar]
- Liu, Q.; Dai, Z.G.; Chen, J.Q.; Wen, L.; Gong, Y.C.; Su, J.G. Establishment of molecular identity for kenaf germplasm using SRAP marker. Sci. Agric. Sin. 2013, 10, 1974–1983. [Google Scholar]
- Qiu, Y.; Li, X.X.; Li, Q.X.; Chen, Y.C.; Shen, D.; Wang, H.P.; Song, J.P. Establishment of the molecular identification for radish germplasm using SSR Markers. J. Plant Genet. Resour. 2014, 3, 648–654. [Google Scholar]
- Gao, Y.; Liu, F.Z.; Wang, K.; Wang, D.J.; Gong, X.; Liu, L.J. Establishment of molecular identity card for Malus Mill. originated from China based on the fingerprints of TP-M13-SSR. J. Plant Genet. Resour. 2015, 6, 1290–1297. [Google Scholar]
- Gao, Y.; Liu, F.Z.; Wang, K.; Wang, D.J.; Gong, X.; Liu, L.J. Establishment of molecular ID for some apple germplasm resources. Sci. Agric. Sin. 2015, 19, 3887–3898. [Google Scholar]
- Liebhard, R.; Gianfrancechi, L.; Koller, B.; Ryder, C.D.; Tarchini, R.; Van, D.W.E.; Gessler, C. Development and characterization of 140 new microsatellites in apple (Malus × domestica Borkh.). Mol. Breed. 2002, 10, 217–241. [Google Scholar] [CrossRef]
- Yang, R.C.; Yeh, F.C. Multilocus structure in Pinus contorta Dougl. Theor. Appl. Genet. 1993, 87, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Sau, S.; Pastore, C.; D’hallewin, G.; Dondini, L.; Bacchetta, G. Characterisation of microsatellite loci in Sardinian pears (Pyrus communis L. and P. spinosa Forssk.). Sci. Hortic. 2020, 270, 109443. [Google Scholar] [CrossRef]
- Arnal, A.; Gogorcena, Y.; Tardío, J.; Roldán-Ruiz, I.; Lázaro, A. Simple Sequence Repeat Characterisation of Traditional Apple Cultivars (Malus domestica Borkh.) Grown in the Region of Madrid (Central Spain). Plant Mol. Biol. Rep. 2020, 38, 676–690. [Google Scholar] [CrossRef]
- Ferreira, V.; Ramos-Cabrer, A.M.; Carnide, V.; Pinto-Carnide, O.; Assunção, A.; Marreiros, A.; Rodrigues, R.; Pereira-Lorenzo, S.; Castro, I. Genetic pool structure of local apple cultivars from Portugal assessed by microsatellites. Tree Genet. Genomes 2016, 12, 36. [Google Scholar] [CrossRef]
- Khachtib, Y.; Zinelabidine, L.H.; Bouda, S.; Hamdali, H.; Hammada, S.; Haddioui, A. Genetic characterization of cultivated apple (Malus x domestica Borkh.) in Morocco using microsatellite (SSR) markers. Ecol. Genet. Genom. 2022, 23, 100122. [Google Scholar] [CrossRef]
- Galli, Z.; Halasz, G.; Kiss, E.; Heszky, L.; Dobranszki, J. Molecular identification of commercial apple cultivars with microsatellite markers. Hortscience 2005, 40, 1974–1977. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Wang, K.; Wang, D.J.; Gong, X.; Liu, L.J.; Liu, F.Z. Molecular ID Establishment of Apple Cultivars by TP-M13-SSR. Acta Hortic. Sin. 2016, 43, 25–37. [Google Scholar]
- Gao, Y.; Wang, D.J.; Wang, K.; Cong, P.H.; Li, L.W.; Piao, J.C. Genetic relationship and structure analysis of 15 species of Malus Mill. based on SNP markers. Sci. Agric. Sin. 2020, 53, 3333–3343. [Google Scholar]
- Gao, Y.; Wang, D.J.; Wang, K.; Cong, P.H.; Li, L.W.; Piao, J.C. Genetic diversity and population structure of 17 species of Malus Mill. native to China based on fluorescent SSR analysis. J. Fruit Sci. 2020, 37, 1611–1622. [Google Scholar]
- Xiao, Z.A.; Cheng, M.H.; Li, X.L. Reltedness and evolution among populations of Chinese apple species. J. Southwest Agric. Univ. 1986, 2, 108–112. [Google Scholar]
- Powell, W.; Morgante, M.; Andre, C.; Hanafey, M.; Vogel, J.; Tingey, S.; Rafalski, A. The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol. Breed. 1996, 2, 225–238. [Google Scholar] [CrossRef]
- Ming, J.; Zhang, Q.X.; Ru, C.G.; Yan, X.L.; Mao, Q.S. The Evolution of identification of gardening plants with DNA molecular Markers. World For. Res. 2003, 5, 12–16. [Google Scholar]
- Li, L.; Yang, J.B.; Mackill, D.J.; Colowit, P.M. Comparison of different detection methods for rice SSR analysis. Chin. J. Rice Sci. 2000, 3, 185–188. [Google Scholar]
- Hao, C.Y.; Wang, L.F.; Jia, J.Z.; Dong, Y.C.; Zhang, X.Y. Comparison of fluorescence and silver-staining detection systems of microsatellite markers. Acta Agron. Sin. 2005, 2, 144–149. [Google Scholar]
- Gong, Y.M.; Hu, Q.Z.; Mao, W.H.; Li, Y.D.; Zhang, G.W.; Ding, J. Application and evaluation of fluorescent EST-SSR markers detection technique with capillary electrophoresis in pea. Acta Agric. Zhejiangensis 2009, 6, 540–543. [Google Scholar]
- Cheng, B.Y.; Xia, J.H.; Gong, J.Y.; Yang, S.H. Application of capillary electrophoresis detection with fluorescent SSR markers in rice DNA fingerprint identification. Chin. J. Rice Sci. 2011, 6, 672–676. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Accession Name | Species | No. | Accession Name | Species |
|---|---|---|---|---|---|
| 1 | Huahong | M. asiatica | 24 | Sankuaishihaitang1 | M. robusta |
| 2 | Xifuhaitang | M. micromalus | 25 | Xiaofanshanbaleng | M. robusta |
| 3 | Dongbaiguo | M. sieversii | 26 | Sankuaishihaitang1shisheng | M. robusta |
| 4 | Hongguozi | M. sieversii | 27 | Xiaofanshanhaitangshisheng | M. robusta |
| 5 | Huochengbaizuozi | M. sieversii | 28 | Mudanjianghaitangshisheng | M. prunifolia |
| 6 | Keziaerma | M. sieversii | 29 | Wanbaihaitangshisheng | M. robusta |
| 7 | Ningmenghaitang | M. sieversii | 30 | Regunzi | M. robusta |
| 8 | Aliusitan | M. sieversii | 31 | Lenghaitang | M. robusta |
| 9 | Baihaitang | M. sieversii | 32 | Xiaofanshanhaitang4 | M. robusta |
| 10 | Donghongguo | M. sieversii | 33 | Zaobaihaitang | M. robusta |
| 11 | Mianpingguo | domestica subsp. chinensis | 34 | Regunzishisheng | M. robusta |
| 12 | Xiaofanshanbinzishisheng | M. domestica subsp. chinensis | 35 | Xiaofanshanbinzishisheng | M. robusta |
| 13 | Yuanyehaitang | M. prunifolia | 36 | Daguchengbalenghaitangshisheng | M. robusta |
| 14 | Pingyaoguchenghaitang | M. prunifolia | 37 | Luanzhuangshaguoshisheng | M. robusta |
| 15 | Daqiu | M. prunifolia | 38 | Xiaofanshanbalenghaitangshisheng | M. robusta |
| 16 | Daqiuzi | M. prunifolia | 39 | Pingdinghaitang | M. robusta |
| 17 | Jilinxiaohuanghaitang | M. prunifolia | 40 | Sankuaishihaitang2 | M. robusta |
| 18 | Qiuzi | M. prunifolia | 41 | Xiaofanshanbaleng1 | M. robusta |
| 19 | Niumamahaitang | M. prunifolia | 42 | Honghaitang | M. robusta |
| 20 | Dongbeihuanghaitang | M. prunifolia | 43 | Zisexiaofanshanbaleng | M. robusta |
| 21 | Laiwunanyan | prunifolia | 44 | Duanzhigunzi | M. robusta |
| 22 | Hebeibaleng | M. robusta | 45 | Wanbaihaitang | M. robusta |
| 23 | Hebeipingdinghaitangshisheng | M. robusta | 46 | Huailaibalenghaitang | M. robusta |
| Primer Name | Forward Primer Sequence | Reverse Primer Sequence | Annealing Temperature (°C) |
| CH01f03b | F:CACGACGTTGTAAAACGACGAGAAGCAAATGCAAAACCC | R:CTCCCCGGCTCCTATTCTAC | 58 |
| CH02b12 | F:CACGACGTTGTAAAACGACGGCAGGCTTTACGATTATGC | R:CCCACTAAAAGTTCACAGGC | 59 |
| CH03d07 | F:CACGACGTTGTAAAACGACCAAATCAATGCAAAACTGTCA | R:GGCTTCTGGCCATGATTTTA | 51 |
| CH04e03 | F:CACGACGTTGTAAAACGACTTGAAGATGTTTGGCTGTGC | R:TGCATGTCTGTCTCCTCCAT | 60 |
| CH04h02 | F:CACGACGTTGTAAAACGACGGAAGCTGCATGATGAGACC | R:CTCAAGGATTTCATGCCCAC | 55.5 |
| CH05c06 | F:CACGACGTTGTAAAACGACATTGGAACTCTCCGTATTGTGC | R:ATCAACAGTAGTGGTAGCCGGT | 58 |
| CH05d08 | F:CACGACGTTGTAAAACGACTCATGGATGGGAAAAAGAGG | R:TGATTGCCACATGTCAGTGTT | 55.5 |
| CH02a04 | F:CACGACGTTGTAAAACGACGAAACAGGCGCCATTATTTG | R:AAAGGAGACGTTGCAAGTGG | 58 |
| CH05b06 | F:CACGACGTTGTAAAACGACACAAGCAAACCTAATACCACCG | R:GAGACTGGAAGAGTTGCAGAGG | 55 |
| CH05d04 | F:CACGACGTTGTAAAACGACACTTGTGAGCCGTGAGAGGT | R:TCCGAAGGTATGCTTCGATT | 60 |
| CH01f07a | F:CACGACGTTGTAAAACGACCCCTACACAGTTTCTCAACCC | R:CGTTTTTGGAGCGTAGGAAC | 59 |
| CH04g07 | F:CACGACGTTGTAAAACGACCCCTAACCTCAATCCCCAAT | R:ATGAGGCAGGTGAAGAAGGA | 57 |
| CH05e04 | F:CACGACGTTGTAAAACGACAAGGAGAAGACCGTGTGAAATC | R:CATGGATAAGGCATAGTCAGGA | 58 |
| CH05h12 | F:CACGACGTTGTAAAACGACTTGCGGAGTAGGTTTGCTTT | R:TCAATCCTCATCTGTGCCAA | 60 |
| Marker | Number of Observed Alleles (Na) | Number of Effective Alleles (Ne) | Expected Heterozygosity (He) | Observed Heterozygosity (Ho) | Shannon’s Information Index (I) | Accessions Distinguished | Identification Ratio |
|---|---|---|---|---|---|---|---|
| CH01f03b | 10 | 5.2834 | 0.8092 | 0.0851 | 1.9134 | 12 | 26.09% |
| CH02b12 | 15 | 8.0304 | 0.8787 | 0.7234 | 2.3474 | 27 | 58.70% |
| CH03d07 | 19 | 5.9606 | 0.8368 | 0.8723 | 2.2809 | 28 | 60.87% |
| CH04e03 | 22 | 10.9072 | 0.9099 | 0.7872 | 2.6601 | 35 | 76.09% |
| CH04h02 | 23 | 12.0914 | 0.9185 | 0.8936 | 2.7592 | 33 | 71.74% |
| CH05c06 | 17 | 7.2782 | 0.8746 | 0.7872 | 2.4060 | 24 | 52.17% |
| CH05d08 | 18 | 5.7203 | 0.8298 | 0.7660 | 2.2189 | 26 | 56.32% |
| CH02a04 | 16 | 6.6856 | 0.8497 | 0.8298 | 2.2970 | 24 | 52.17% |
| CH05b06 | 27 | 10.9072 | 0.9054 | 0.7660 | 2.7453 | 31 | 67.39% |
| CH05d04 | 19 | 6.5854 | 0.8581 | 0.7021 | 2.3501 | 30 | 65.22% |
| CH01f07a | 22 | 11.4175 | 0.9131 | 0.7234 | 2.6865 | 30 | 65.22% |
| CH04g07 | 22 | 8.1385 | 0.8796 | 0.7660 | 2.5478 | 31 | 67.39% |
| CH05e04 | 17 | 8.0304 | 0.8794 | 0.6596 | 2.3980 | 29 | 63.04% |
| CH05h12 | 17 | 7.3345 | 0.8642 | 0.9362 | 2.3174 | 33 | 71.74% |
| Mean | 18.9 | 8.1693 | 0.8719 | 0.7356 | 2.4234 | 28.1 | 61.01% |
| Number | Primer Combination | Accessions Not Distinguished | Identification Rate |
|---|---|---|---|
| 1 | CH04e03 and CH04h02 | 23, 24, 25, 42 | 91.30% |
| 2 | CH04e03 and CH05b06 | 23, 24, 25, 31, 37, 39, 42, 43 | 82.61% |
| 3 | CH04e03 and CH01f07a | 24, 25, 31, 45 | 91.30% |
| 4 | CH04e03 and CH04g07 | 4, 6, 23, 24, 25, 37 | 86.96% |
| 5 | CH04h02 and CH05b06 | 23, 25, 30, 38, 39 | 89.13% |
| 6 | CH04h02 and CH01f07a | 38, 39 | 95.65% |
| 7 | CH04h02 and CH04g07 | 23, 25 | 95.65% |
| 8 | CH05b06 and CH01f07a | 18, 24, 38, 39 | 91.30% |
| 9 | CH05b06 and CH04g07 | 23, 24, 25, 33, 37, 44 | 86.96% |
| 10 | CH01f07a and CH04g07 | 37, 40 | 95.65% |
| Code | Primer | Code | Primer | ||||
|---|---|---|---|---|---|---|---|
| CH04h02 | CH01f07a | CH04g07 | CH04h02 | CH01f07a | CH04g07 | ||
| 01 | 141 | 104 | 152 | 13 | 201 | 208 | 186 |
| 02 | 163 | 108 | 156 | 14 | 203 | 210 | 188 |
| 03 | 177 | 112 | 160 | 15 | 205 | 212 | 190 |
| 04 | 183 | 116 | 164 | 16 | 211 | 214 | 192 |
| 05 | 185 | 184 | 166 | 17 | 213 | 216 | 194 |
| 06 | 187 | 186 | 168 | 18 | 215 | 218 | 200 |
| 07 | 189 | 190 | 174 | 19 | 217 | 220 | 202 |
| 08 | 191 | 192 | 176 | 20 | 219 | 224 | 206 |
| 09 | 193 | 194 | 178 | 21 | 221 | 244 | 210 |
| 10 | 195 | 196 | 180 | 22 | 247 | −9 | −9 |
| 11 | 197 | 202 | 182 | 23 | −9 | ||
| 12 | 199 | 206 | 184 | 24 | |||
| No. of Accession | Molecular Identity | No. of Accession | Molecular Identity | No. of Accession | Molecular Identity |
|---|---|---|---|---|---|
| 1 | 061012150811 | 17 | 061112130818 | 33 | 051914160608 |
| 2 | 031213130607 | 18 | 061522220712 | 34 | 061615150212 |
| 3 | 051414180214 | 19 | 102114160615 | 35 | 232310140606 |
| 4 | 070717190610 | 20 | 010212170612 | 36 | 061313150512 |
| 5 | 060817171213 | 21 | 051905140518 | 37 | 061613170606 |
| 6 | 071609150610 | 22 | 060815170105 | 38 | 061613170506 |
| 7 | 060705150311 | 23 | 232322220512 | 39 | 131613150512 |
| 8 | 070912171014 | 24 | 060822220105 | 40 | 101322220612 |
| 9 | 050914180219 | 25 | 061012211717 | 41 | 232317200505 |
| 10 | 101812170615 | 26 | 111603080516 | 42 | 062001040606 |
| 11 | 161922220506 | 27 | 080809110412 | 43 | 060802020609 |
| 12 | 040917190609 | 28 | 080908121220 | 44 | 051913130113 |
| 13 | 051207140808 | 29 | 060809172222 | 45 | 161909150505 |
| 14 | 091722220606 | 30 | 051313130206 | 46 | 030817170721 |
| 15 | 040809120606 | 31 | 162205140512 | ||
| 16 | 061915160507 | 32 | 040806160609 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Wang, D.; Wang, K.; Sun, S.; Tian, W.; Li, Z.; Wang, G.; Lu, X.; Liu, Z.; Li, Q.; et al. Genetic Structure and Molecular Identities of 46 Apple Landraces (Malus Mill.) in China. Agronomy 2023, 13, 1262. https://doi.org/10.3390/agronomy13051262
Wang L, Wang D, Wang K, Sun S, Tian W, Li Z, Wang G, Lu X, Liu Z, Li Q, et al. Genetic Structure and Molecular Identities of 46 Apple Landraces (Malus Mill.) in China. Agronomy. 2023; 13(5):1262. https://doi.org/10.3390/agronomy13051262
Chicago/Turabian StyleWang, Lin, Dajiang Wang, Kun Wang, Simiao Sun, Wen Tian, Zichen Li, Guangyi Wang, Xiang Lu, Zhao Liu, Qingshan Li, and et al. 2023. "Genetic Structure and Molecular Identities of 46 Apple Landraces (Malus Mill.) in China" Agronomy 13, no. 5: 1262. https://doi.org/10.3390/agronomy13051262