
Effects of Drought Stress on Annual Herbaceous Plants under Different Mixed Growth Conditions in Desert Oasis Transition Zone of the Hexi Corridor
Abstract
:1. Introduction
2. Materials and Methods
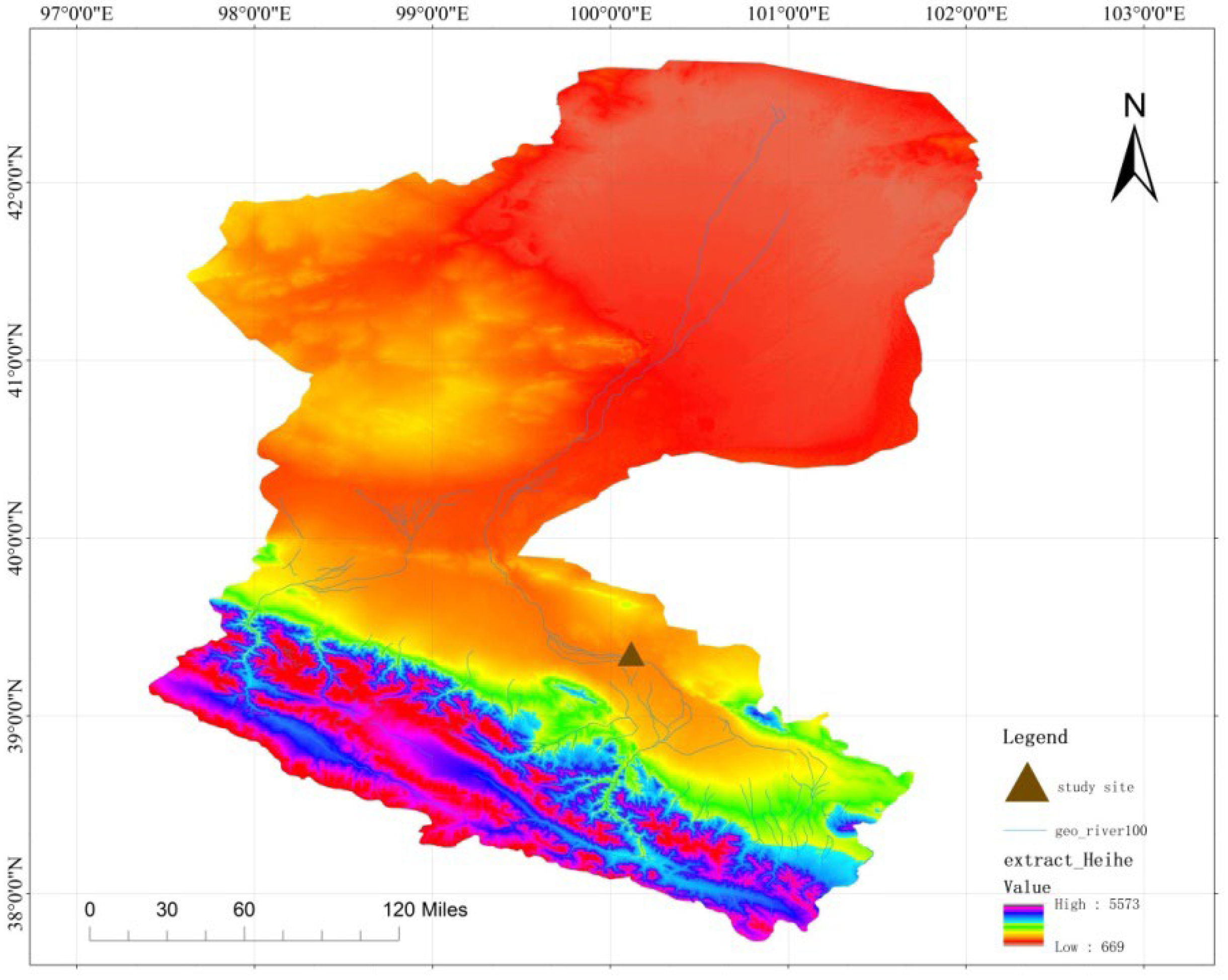
2.1. Overview of the Study Area
2.2. Experimental Design
2.3. Measurement Methods for Indicators
2.4. Data Processing
3. Results and Analysis
3.1. Characteristics of Plant Germination under Different Mixed Conditions
3.2. Effects of Drought Stress on the Growth of Annual Plants under Different Mixed Conditions
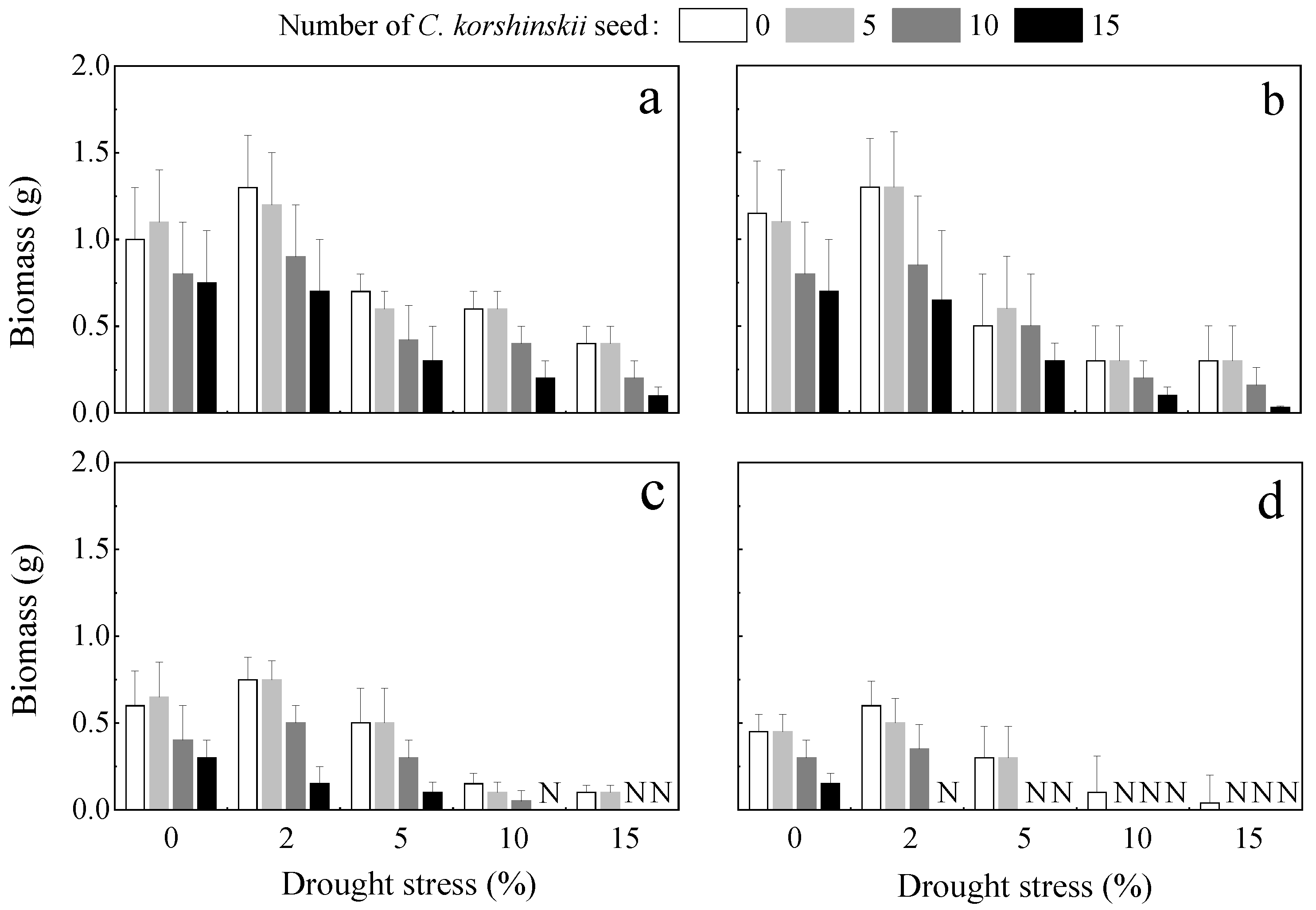
3.2.1. Amount of Biomass
3.2.2. Survival Rate
3.3. Effects of Drought Stress on the Root-Shoot Ratio of Annual Plants under Different Mixed Conditions
3.4. Plant Physiology under Different Drought Stress Intensities
3.4.1. Root Vitality
3.4.2. Chlorophyll Content
4. Discussion
4.1. Effect of Different Mixing Conditions on the Germination of Annual Herbaceous Plants
4.2. Effects of Different Mixed Planting and Drought Stress on the Growth and Survival of Annual Herbaceous Plants
4.3. Effects of Different Mixes and Drought Stress on the Physiology of Annual Herbaceous Plants
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oba, G.; Weladji, R.B.; Msangameno, D.J.; Kaitira, L.M.; Stave, J. Scaling effects of proximate desertification drivers on soil nutrients in northeastern Tanzania. J. Arid Environ. 2008, 72, 1820–1829. [Google Scholar] [CrossRef]
- Xiao, H.; Li, X.; Song, Y.; Li, S. Impact of Evolvement of Soil-plant System on Engineering of Mobile Sand Dunes Control. Sci. Silvae Sin. 2004, 40, 24–30. (In Chinese) [Google Scholar]
- Friedman, J. The effect of competition by adult zygophy1lum dumosum Bioss. on seeding of Artemisia herba-abta Asso in the Negev Desert of Israel. J. Ecol. 1971, 59, 775–782. [Google Scholar] [CrossRef]
- Zhang, D.Y. Researches on Theoretical Ecological; Higher Education Press: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Went, F.W. The ecology of desert plants. Sci. Am. 1955, 192, 68–75. [Google Scholar] [CrossRef]
- Zhou, H.; Zhao, W.; He, Z. Water sources of Nitraria sibirica and response to precipitation in two desert habitats. Chin. J. Appl. Ecol. 2017, 28, 2083–2092. (In Chinese) [Google Scholar]
- Han, G.; Zhao, Z. Light response characteristics of photosynthesis of four xerophilous shrubs under different soil moistures. Acta Ecol. Sin. 2010, 30, 4019–4026. (In Chinese) [Google Scholar]
- Yan, X.F.; Zhou, L.B.; Si, B.B.; Sun, Y.; Gao, Y.F. Stress effects of simulated drought by polyethylene glycol on the germination of Caragana korshinskii Kom. Seeds under different temperature conditions. Acta Ecol. Sin. 2016, 36, 1989–1996. (In Chinese) [Google Scholar]
- Wang, G.; Zhao, W. Effect of sand burying on seed germination and seeding growth of Haloxylon ammodendron. J. Des. Res 2015, 35, 338–344. (In Chinese) [Google Scholar]
- Liang, C.; Liu, Z.; Zhu, Z.; Wang, W. Specific diversity and distribution characteristics of annual synusia in Alashan desert. Chin. J. Appl. Ecol. 2003, 14, 897–903. (In Chinese) [Google Scholar]
- Wang, X.P.; Berndtsson, R.; Li, X.R.; Kang, E.S. Water balance change for a re-vegetated xerophyte shrub area/Changement du bilan hydrique d’une zone replantée d’arbustes xérophiles. Hydrol. Sci. J. 2004, 49, 283–295. [Google Scholar] [CrossRef]
- Liu, B. Responses of Shrubs to Precipitation Pulses in Desert Region; Bulletin of Chinese Academy of Science: Beijing, China, 2009. (In Chinese) [Google Scholar]
- Shi, W.; Xu, H.; Zhao, X.; Ling, H.; Li, Y. Physiological and biochemical responses to drought stress during seed germination of Glycyrrhiza inflata. Acta Ecol. Sin. 2010, 30, 2112–2117. (In Chinese) [Google Scholar]
- Li, Z.; Zhang, W.; Cui, Y. Effects of PEG Simulated Drought Stress On Seed Germination and Growth Physiology of Quercus variabilis. Acta Bot. Boreali-Occident. Sin. 2013, 33, 2043–2049. (In Chinese) [Google Scholar]
- Guo, Y.; MI, F.; Yan, L.; Ren, Y.; Lv, S.; Fu, B. Physiological response to drought stresses and drought resistance evaluation of different Kentucky bluegrass varieties. Acta Pratacult. Sin. 2014, 23, 220–228. (In Chinese) [Google Scholar]
- Michel, B.E.; Kaufmann, M.R. The osmotic potential of polyethylene glycol 6000. Plant Physiol. 1973, 51, 914–916. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Liu, J.; Bei, Z.; Liu, C.; Gao, Y. Characteristics of seed germination and seedling growth of Caragana korshinskii under different light intensities. Chin. J. Appl. Ecol. 2015, 34, 912–918. (In Chinese) [Google Scholar]
- Zhang, Z.; Qu, W. Instruction about Plant Physiology Experiment; Higher Education Press: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Fennner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Li, B.; Chen, J.; Andrew, R.W. A Literature Review on Plant Competition. Chin. Bull. Bot. 1998, 7, 20–31. (In Chinese) [Google Scholar]
- Li, C. Research on Species Diversity and Community Stability of Minqin Liangucheng Nature Reserve; Northwest Normal University: Lanzhou, China, 2005. (In Chinese) [Google Scholar]
- Yan, Q.; Liu, Z.; Li, X.; Ma, J. Effects of burial on seed germination characteristics of 65 plant species on Horqin semi-arid. Chin. J. Appl. Ecol. 2007, 18, 777–782. (In Chinese) [Google Scholar]
- Liu, Z. Plant Regenerative Strategies in the Horqin Sand Land; China Meteorological Press: Beijing, China, 2010; pp. 113–121. (In Chinese) [Google Scholar]
- Luo, G.; Wang, J.; Yan, X.; Chen, N.; Zhang, Y. Effects of drought on seed germination and seedling growth of Nitraria tangutorum. J. Des. Res. 2014, 34, 1537–1543. (In Chinese) [Google Scholar]
- Liu, H.-L.; Shi, X.; Wang, J.-C.; Yin, J.-K.; Huang, Z.-Y.; Zhang, D.-Y. Effects of sand burial, soil water content and distribution pattern of seeds in sand on seed germination and seedling survival of Eremosparton songoricum (Fabaceae), a rare species inhabiting the moving sand dunes of the Gurbantunggut Desert of China. Plant Soil 2011, 345, 69–87. [Google Scholar] [CrossRef]
- He, S.; Liang, Z.; Wei, L.; Zhou, Z. Growth and Physiological Characteristics of Wild Sour Jujube Seedlings from Two Provenances under Soil Water Stress. Acta Bot. Boreali-Occident. Sin. 2009, 29, 1387–1393. (In Chinese) [Google Scholar]
- Sang, Z.; Ma, L.; Chen, F. Growth and Physiological Characteristics of Magnoliawuf engensis Seedlings under Drought Stress. Acta Bot. Boreali-Occident. Sin. 2011, 31, 109–115. (In Chinese) [Google Scholar]
- Watkinson, A.R.; Harper, J.L. The demography of a sand dune annual: Vulpia fasciculata. Ecology 1978, 66, 15–33. [Google Scholar] [CrossRef]
- Harper, J.L.; Williams, J.J.; Sagar, G.R. The behaviour of seeds in soil. I. The heterogeneity of the soil surface and its role in determining the establishment of plants from seed. J. Ecol. 1965, 53, 273–286. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Li, X.; Zhang, Z.; Wang, G. Advances and Prospect of Researches on Desert Plant Life History Strategies. J. Des. Res. 2005, 3, 306–314. (In Chinese) [Google Scholar]
- Su, M. Effects of Precipitation on Functional Characters and Interspecific Relationship between Reaumuria soongaric and Salsola passerina; Gansu Agricultural University: Lanzhou, China, 2019. (In Chinese) [Google Scholar]
- Lin, Y.; Berger, U.; Grimm, V.; Ji, Q.R. Differences between symmetric and asymmetric facilitation matter: Exploring the interplay between modes of positive and negative plant interactions. J. Ecol. 2012, 100, 1482–1491. [Google Scholar] [CrossRef] [Green Version]
- Maestre, F.T.; Callaway, R.M.; Valladares, F.; Lortie, C.J. Refining the stress-gradient hypothesis for competition and facilitation in plant communities. J. Ecol. 2009, 97, 199–205. [Google Scholar] [CrossRef]
- Yang, J.; Wang, Y. Effects of drought stress simulated by PEG on seed germination of four desert plant species. Acta Pratacult. Sin. 2012, 21, 23–29. (In Chinese) [Google Scholar]
- Liu, C.; Wang, W.; Cui, J.; Li, S. Effects of Drought Stress on Photosynthesis Characteristics and Biomass Allocation of Glycyrrhiza uralensis. J. Des. Res. 2006, 1, 142–145. (In Chinese) [Google Scholar]
- Yang, F.; Miao, L.; Xu, X.; Li, C. Progress in Research of Plant Responses to Drought Stress. Chin. J. Appl. Environ. Biol. 2007, 4, 586–591. (In Chinese) [Google Scholar]
- Brouwer, R. Some Aspects of the Equilibrium between Overground and Underground Plant Parts; Jaarboek IBS: Wageningen, The Netherlands, 1963; pp. 31–39. [Google Scholar]
- Zeng, Y.; Wang, Y.; Sa, R.; Tian, X. Response of seed germination of three xeromorphic shrubs to drought stress. Chin. J. Appl. Ecol. 2002, 13, 953–956. (In Chinese) [Google Scholar]
- Han, J.; Wang, X.; Zhang, Z. Effects of Surface Soil Drought on Root Spatial Distribution and Activity of White Clovers. Chin. Agric. Sci. Bull. 2007, 23, 458–461. (In Chinese) [Google Scholar]
- Li, F.L.; Bao, W.K.; Wu, N. Morphological and physiological responses of current sophora davidii seedlings to drought stress. Acta Ecol. Sin. 2009, 29, 5406–5416. (In Chinese) [Google Scholar]
- Ren, Y.; Yin, F.; Liu, S.; Qi, W. Effects of Drought Stress on Mulberry Varieties of Chlorophyll, Water Saturation Deficit and Malondialdehyde. Southwest China J. Agric. Sci. 2016, 29, 2583–2587. (In Chinese) [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indexes | Plant Species | C. korshinskii Number | Drought Stress | C. korshinskii Number × Drought Stress | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p | df | F | p | df | F | p | |||
| Morphology | Biomass | S. viridis | 3 | 13.236 | <0.01 | 4 | 27.254 | <0.01 | 12 | 0.688 | 0.753 |
| C. virgata | 7.299 | <0.01 | 27.693 | <0.01 | 0.578 | 0.847 | |||||
| H. arachnoideus | 29.644 | <0.01 | 49.406 | <0.01 | 2.414 | 0.018 | |||||
| B. dasyphylla | 24.561 | <0.01 | 32.890 | <0.01 | 3.774 | <0.01 | |||||
| Survival rate | S. viridis | 3 | 37.877 | <0.01 | 4 | 39.560 | <0.01 | 12 | 2.280 | 0.025 | |
| C. virgate | 123.698 | <0.01 | 65.628 | <0.01 | 8.660 | <0.01 | |||||
| H. arachnoideus | 133.251 | <0.01 | 223.270 | <0.01 | 10.282 | <0.01 | |||||
| B. dasyphylla | 133.251 | <0.01 | 223.270 | <0.01 | 10.282 | <0.01 | |||||
| Root shoot ratio | S. viridis | 3 | 0.407 | 0.749 | 4 | 15.470 | <0.01 | 12 | 0.395 | 0.957 | |
| C. virgate | 0.509 | 0.678 | 12.853 | <0.01 | 0.263 | 0.992 | |||||
| H. arachnoideus | 13.812 | <0.01 | 8.412 | <0.01 | 12.496 | <0.01 | |||||
| B. dasyphylla | 62.136 | <0.01 | 42.562 | <0.01 | 25.637 | <0.01 | |||||
| physiology | Root activity | S. viridis | 3 | 6.817 | <0.01 | 4 | 12.223 | <0.01 | 12 | 0.205 | 0.997 |
| C. virgate | 4.881 | <0.01 | 10.311 | <0.01 | 0.171 | 0.999 | |||||
| H. arachnoideus | 4.889 | <0.01 | 9.432 | <0.01 | 0.95 | 0.504 | |||||
| B. dasyphylla | 29.707 | <0.01 | 21.619 | <0.01 | 1.850 | 0.072 | |||||
| Chlorophyll | S. viridis | 3 | 16.844 | <0.01 | 4 | 15.991 | <0.01 | 12 | 0.411 | 0.950 | |
| C. virgata | 6.153 | <0.01 | 8.607 | <0.01 | 0.152 | 0.999 | |||||
| H. arachnoideus | 57.240 | <0.01 | 15.386 | <0.01 | 0.621 | 0.811 | |||||
| B. dasyphylla | 27.544 | <0.01 | 13.279 | <0.01 | 2.561 | 0.013 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gou, Q.; Song, B.; Li, Y.; Xi, L.; Wang, G. Effects of Drought Stress on Annual Herbaceous Plants under Different Mixed Growth Conditions in Desert Oasis Transition Zone of the Hexi Corridor. Sustainability 2022, 14, 14956. https://doi.org/10.3390/su142214956
Gou Q, Song B, Li Y, Xi L, Wang G. Effects of Drought Stress on Annual Herbaceous Plants under Different Mixed Growth Conditions in Desert Oasis Transition Zone of the Hexi Corridor. Sustainability. 2022; 14(22):14956. https://doi.org/10.3390/su142214956
Chicago/Turabian StyleGou, Qianqian, Bing Song, Yuda Li, Lulu Xi, and Guohua Wang. 2022. "Effects of Drought Stress on Annual Herbaceous Plants under Different Mixed Growth Conditions in Desert Oasis Transition Zone of the Hexi Corridor" Sustainability 14, no. 22: 14956. https://doi.org/10.3390/su142214956