 Márcio de Carvalho Moretzsohn1*
Márcio de Carvalho Moretzsohn1* João Francisco dos Santos2
João Francisco dos Santos2 Andrea Rocha Almeida Moraes2
Andrea Rocha Almeida Moraes2 Adriana Regina Custódio1
Adriana Regina Custódio1 Marcos Doniseti Michelotto3
Marcos Doniseti Michelotto3 Namrata Mahrajan4
Namrata Mahrajan4 Soraya Cristina de Macedo Leal-Bertioli4,5Ignácio José Godoy2David John Bertioli4,6
Soraya Cristina de Macedo Leal-Bertioli4,5Ignácio José Godoy2David John Bertioli4,6- 1Plant Genetics Laboratory, Embrapa Genetic Resources and Biotechnology, Brasília, DF, Brazil
- 2Grain and Fiber Center, Agronomic Institute of Campinas (IAC), Campinas, SP, Brazil
- 3Grain and Fiber Center, Agronomic Institute of Campinas (IAC), Pindorama, SP, Brazil
- 4Institute of Plant Breeding, Genetics and Genomics, University of Georgia, Athens, GA, United States
- 5Department of Plant Pathology, University of Georgia, Athens, GA, United States
- 6Department of Crop and Soil Science, University of Georgia, Athens, GA, United States
Introduction: Fungal foliar diseases can severely affect the productivity of the peanut crop worldwide. Late leaf spot is the most frequent disease and a major problem of the crop in Brazil and many other tropical countries. Only partial resistance to fungal diseases has been found in cultivated peanut, but high resistances have been described on the secondary gene pool.
Methods: To overcome the known compatibility barriers for the use of wild species in peanut breeding programs, we used an induced allotetraploid (Arachis stenosperma × A. magna)4x, as a donor parent, in a successive backcrossing scheme with the high-yielding Brazilian cultivar IAC OL 4. We used microsatellite markers associated with late leaf spot and rust resistance for foreground selection and high-throughput SNP genotyping for background selection.
Results: With these tools, we developed agronomically adapted lines with high cultivated genome recovery, high-yield potential, and wild chromosome segments from both A. stenosperma and A. magna conferring high resistance to late leaf spot and rust. These segments include the four previously identified as having QTLs (quantitative trait loci) for resistance to both diseases, which could be confirmed here, and at least four additional QTLs identified by using mapping populations on four generations.
Discussion: The introgression germplasm developed here will extend the useful genetic diversity of the primary gene pool by providing novel wild resistance genes against these two destructive peanut diseases.
Introduction
Cultivated peanut (Arachis hypogaea L.) is an important oilseed crop, grown throughout the tropics and sub-tropics. It is also widely used for human and animal consumption being a valuable source of dietary protein. Globally, 53.9 million tons of unshelled peanut were produced in 2021 in 32.7 million ha (FAOSTAT – Food and Agriculture Organization of the United Nations, 2022). Brazil only produced approximately 1.4% of this total; however, the crop is expanding in the country. In the last decade, the national production and the area planted increased 164% and 96%, respectively, reaching 746,700 tons of unshelled peanut harvested in 200,100 ha in the 2021/2022 season (CONAB - Companhia Nacional de Abastecimento, 2023). Peanut production in Brazil is highly concentrated in the state of São Paulo. In 2021/2022, São Paulo produced 692,700 tons of unshelled peanuts (approximately 90% of the Brazilian peanut production), of which more than 50% were shelled and processed for export (CONAB - Companhia Nacional de Abastecimento, 2023). Breeding programs are active in the country and releasing new cultivars to attend to this market. However, it is necessary to prepare this chain of production to be more competitive, especially through reducing costs of production, such as reducing dependence on fungicides.
The genus Arachis is native to South America. It contains 83 described species, assembled into nine taxonomic sections according to their morphology, geographical distribution, and cross-compatibility relationships (Krapovickas and Gregory, 1994; Valls and Simpson, 2005; Valls et al., 2013; Valls and Simpson, 2017; Seijo et al., 2021). Cultivated peanut belongs to section Arachis, which also includes 32 closely related wild species. Of these, 28 are diploid with x = 10 (2n=20), three species are diploid with x = 9 (2n=18), and A. hypogaea and A. monticola Krapov. & Rigoni are allotetraploids (2n=4x=40) with a genome formula AABB (Lavia et al., 2009; Stalker, 2017 and references therein; Seijo et al., 2021). Six genome types, A, B, D, F, K, and G, have been described for the diploid species in section Arachis, differing on the chromosome morphology, distribution patterns of heterochromatic bands and rDNA loci, and cross-compatibility (Smartt et al., 1978; Stalker, 1991; Fernandez and Krapovickas, 1994; Robledo and Seijo, 2010; Silvestri et al., 2015).
Arachis hypogaea has a narrow genetic base and is susceptible to various biotic stresses. Among them, fungal foliar diseases, especially late leaf spot (LLS) [Nothopassalora personata syn. Cercosporidium personatum (Berk. & M.A. Curtis) S.A. Khan & M. Kamal], early leaf spot (Passalora arachidicola syn. Cercospora arachidicola S. Hori), and rust (Puccinia arachidis Speg.), are widespread in most of the producing countries and severely affect productivity. Reductions of up to 70% in peanut yield caused by leaf spots have been described (Backman and Crawford, 1984; Singh et al., 2011; Coutinho and Suassuna, 2014). LLS is the most frequent disease and a major problem of the crop in Brazil. The disease starts at 45–50 days after germination and tends to progress until the end of the crop cycle. Fungus sporulation occurs on the abaxial surface of the leaves and disease symptoms appear approximately 10 days after infection (Shokes and Culbreath, 1997). The main symptom is the presence of round-shaped black lesions on the leaves, which reduces the foliar area and induces defoliation. This causes a reduction in photosynthesis and consequently decreases peanut productivity. Rust is a sporadic disease, but can be highly destructive. Initial symptoms are small yellow round lesions on both surfaces of the leaves, and subsequently, these lesions turn to reddish brown. Unlike LLS, rust-infected leaves tend to remain attached to the plant, which favors the pathogen multiplication and rapid spread of the disease (Moraes and Godoy, 1997). The main damage of rust is the reduction of foliar area available for photosynthesis. To control both diseases, multiple fungicide sprays are needed throughout the growing season, usually at intervals of up to 15 days, depending on weather conditions. Although several fungicides are available, their application significantly increases crop management costs and the risks of soil and environment contamination. Resistant varieties are considered the most efficient way to control these diseases. Few, and only moderate, resistance sources to fungal diseases have been found in A. hypogaea, but high resistance has been described for many Arachis wild species (reviewed by Stalker, 2017). Therefore, there is a growing need to improve peanut resistance to fungal foliar diseases by diversifying its genetic variability utilizing crop wild relatives.
Since most Arachis wild species are diploid and A. hypogaea is an allotetraploid, the introgression of useful wild genes into peanut is not a trivial task. To overcome this, interspecific induced allotetraploid plants have been developed (Burow et al., 2001; Fávero et al., 2006; Kumari et al., 2014; Leal-Bertioli et al., 2015b; Leal-Bertioli et al., 2017; Gao et al., 2021; Bertioli et al., 2021b). In this so-called tetraploid route (Simpson, 1991), species with A and B genomes are intercrossed and the resulting sterile hybrid (AB) is treated with colchicine to duplicate the chromosomes and restore fertility (AABB). The allotetraploid thus obtained can be crossed with A. hypogaea to produce fertile hybrids.
Specially for introgression of wild genes, marker-assisted backcrossing (MABC) is essential, as it enables a rapid recovery of the recurrent parent genome, the pyramiding of multiple useful genes into the same genotype, and considerable time saving (Frisch and Melchinger, 2005; Bharadwaj et al., 2022; Kim et al., 2022). Nevertheless, very few genomic segments of Arachis wild species containing genes of interest have been well-defined to date. Regarding resistance to biotic stresses, a few segments from the species A. cardenasii Krapov. & W.C.Greg. (A genome) with genes for resistance to root-knot nematode (RKN) (Chu et al., 2007), leaf spots, rust, and web blotch (Kolekar et al., 2016; Pandey et al., 2017b; Ahmad et al., 2020; Lamon et al., 2020; Bertioli et al., 2021a) were identified. By using marker-assisted selection, segments conferring RKN resistance were introgressed into released peanut cultivars (Chu et al., 2011; Simpson et al., 2013; Clevenger et al., 2017b). In addition, both by anonymous phenotypic and by marker selection, A. cardenasii-containing segments that confer resistance to leaf spots and rust have been introgressed into peanut in breeding programs worldwide (Varshney et al., 2014; Shasidhar et al., 2020; Godoy et al., 2022; Holbrook et al., 2022). The very large-scale anonymous contribution of A. cardenasii to the peanut crop was recently shown with high-throughput genotyping; 251 peanut lines and cultivars in 30 countries were found to have genetics from this wild species; in almost all cases, the breeders involved were unaware of the wild genetics (Bertioli et al., 2021a).
In previous studies, we identified wild genomic segments conferring resistance to LLS and to RKN in A. stenosperma Krapov. & W.C.Greg. (Leal-Bertioli et al., 2009, Leal-Bertioli et al., 2016; Ballén-Taborda et al., 2019), and a robust QTL (quantitative trait locus) for resistance to rust in A. magna Krapov., W.C.Greg. & C.E.Simpson (Leal-Bertioli et al., 2015a). Arachis stenosperma (A genome 2n=2x=20) is an annual plant, belongs to section Arachis, and is endemic to Brazil (Krapovickas and Gregory, 1994), where it was cultivated for food by native people (Simpson et al., 2001). Arachis magna (B genome, 2n=2x=20) is also annual, belongs to section Arachis, and has been collected in Brazil and Bolivia (Krapovickas and Gregory, 1994; Custódio et al., 2013). An induced allotetraploid (2n=4x=40) was previously developed using these species and found to be resistant to both rust and LLS (Fávero et al., 2015a; Michelotto et al., 2016; and unpublished data). In the present study, this induced allotetraploid was used as the donor parent in successive backcrossings with an elite Brazilian cultivar for the incorporation of wild resistance QTLs into cultivated peanut. We used molecular markers for the selection of plants containing the resistance QTLs with a lower proportion of wild chromosome segments for a faster recovery of the recurrent A. hypogaea genome. The plants obtained were assayed in the field each year, supporting the hypothesis that the selected chromosome regions of A. stenosperma and A. magna conferred resistance to otherwise susceptible plants. Furthermore, we identified additional QTLs controlling LLS and rust.
Material and methods
Plant material
The recurrent parent ‘IAC OL 4’ (A. hypogaea subsp. hypogaea var. hypogaea) is a commercial peanut cultivar developed by the Instituto Agronômico (IAC), Campinas, Brazil (Godoy et al., 2014). It is a high oleic runner cultivar widely grown in São Paulo state, highly yielding, but very susceptible to foliar diseases. The wild donor parent was the induced allotetraploid (A. magna K 30097 × A. stenosperma V 15076)4x (Fávero et al., 2015b), hereafter called MagSten. This induced allotetraploid and its diploid parents have high resistance to foliar fungal diseases (Michelotto et al., 2015, Michelotto et al., 2016). The diploid species were obtained from the Arachis Germplasm Collection, maintained at Embrapa Genetic Resources and Biotechnology (Brasília-DF, Brazil).
Marker-assisted backcrossing scheme
The first round of crosses was made in 2012 using ‘IAC OL 4’ as the female and MagSten as the male parent (Figure 1). True F1 hybrids were identified by microsatellite markers, grown in a greenhouse, and used as male parents on the first backcross with ‘IAC OL 4’. Using ‘IAC OL 4’ as a recurrent parent, generations were advanced to BC4 from 2013 to 2017/2018. On each backcross cycle, progenies were selected using microsatellite markers linked to known desirable QTLs, essentially as described by Moretzsohn et al. (2013). These included a QTL for rust resistance from A. magna K 30097 (on chromosome B08; Leal-Bertioli et al., 2015a) and four QTLs for LLS resistance, located on three chromosomes (A02, A06, and two segments on the middle of A04; Leal-Bertioli et al., 2009) (Supplementary file 1). These QTLs were identified in a different A. stenosperma accession (V 10309), but tentatively used on the accession V 15076 also highly resistant to fungal foliar diseases (Michelotto et al., 2015), and the one incorporated into MagSten. Most of the backcrossings and selfings were also accompanied by different SNP genotyping methods for monitoring the introgressed wild segments and for faster recovery of A. hypogaea genetic background. BC1F1 plants were genotyped with SNP markers dispersed through the 10 A-genome chromosomes using a 384 Illumina BeadXpress array (Supplementary file 1). The BC2 generation was not submitted to background selection. BC3F1 plants were genotyped with the first version of the Thermofisher SNP array (Axiom_Arachis 58k array) that assayed 58,233 SNP markers (Clevenger et al., 2017a; Pandey et al., 2017a), while BC3F2 to BC3F5, BC4F1, and BC4F2 plants were genotyped with the improved array version (Axiom_Arachis v.02) that assays 48,000 SNP markers (Korani et al., 2019). These populations were evaluated for resistance to fungal foliar diseases, pod weight, number of seeds per pod, and pod shape (cultivated, mixed or wild type) in field assays. Data on genotypic and field evaluation were used for the selection of progenies to be advanced to the next generation.
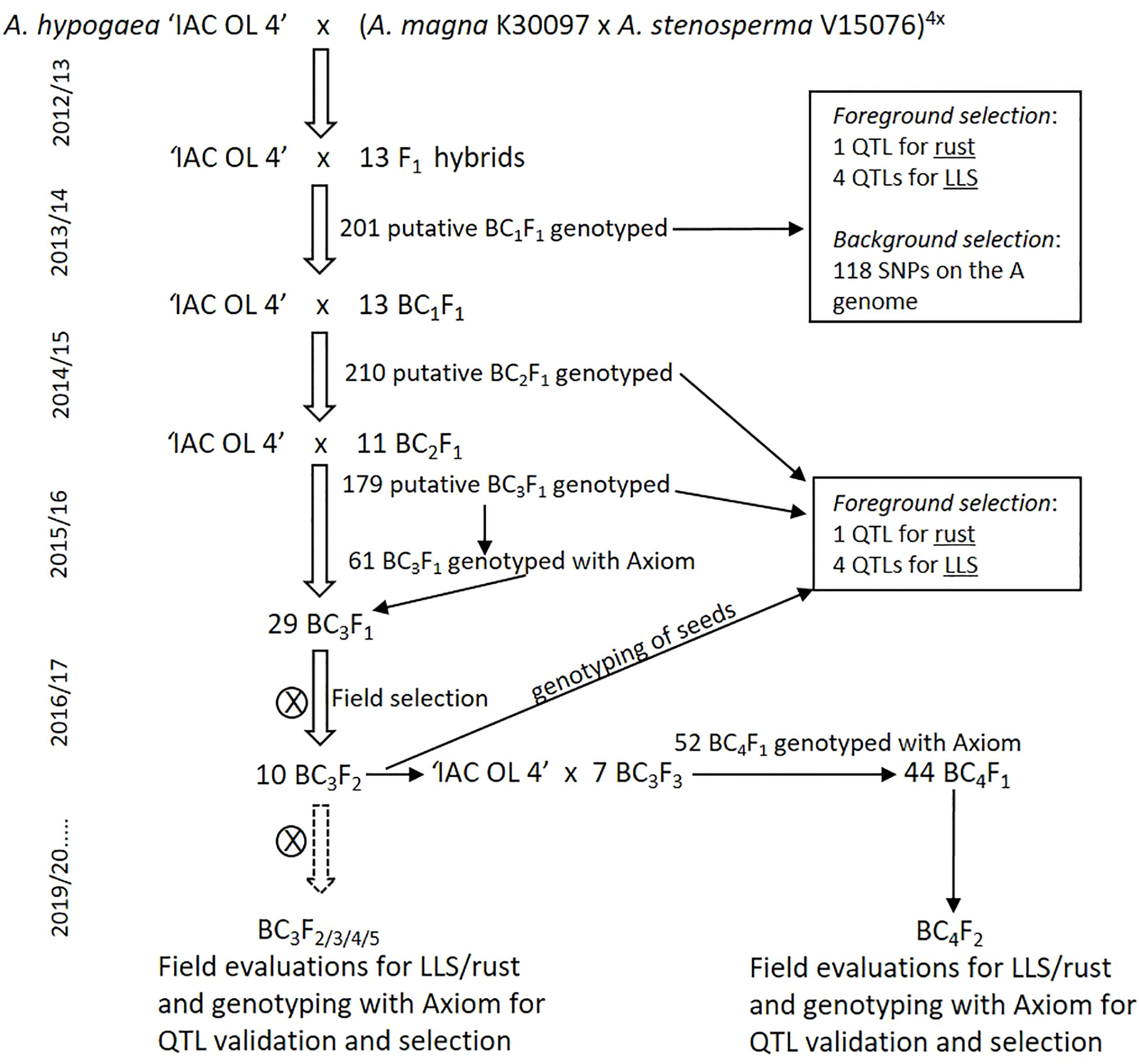
Figure 1 Marker-assisted backcrossing schedule for introgressing chromosome segments from A. magna K 30097 and A. stenosperma V 15076 into the peanut cultivar IAC OL 4.
Field evaluations
Five field trials were conducted in the APTA (Agência Paulista de Tecnologia dos Agronegócios) experimental station in Pindorama, São Paulo State, located in the most important peanut-producing state of Brazil, using generations BC3F2, BC3F3, BC3F4, BC3F5, and BC4F2. Thirty seeds per selected family were planted in 15-m-long single-randomized row plots, using ‘IAC OL 3’ as susceptible control, and ‘IAC Caiapó’ and ‘IAC Sempre Verde’ as resistant checks. MagSten was also included in some of the trials. Field management followed standard technical recommendations for the crop, except for disease control as no fungicide was sprayed. At approximately 120 days after sowing, each plant was evaluated for LLS incidence using the grade scale ranging from “1” (no spots or defoliation) to “9” (many spots on the remaining leaves and a severe defoliation), according to Subrahmanyam et al. (1995), but including the 0.5 scores for intermediate aspects between adjacent integer scores. Rust is a sporadic disease and the incidence was low on four of the five assays. Therefore, it was evaluated using “1” for the presence and “0” for absence of rust lesions in leaves. On the BC4F2 field assay, rust incidence was moderate and the plants were evaluated using the 1–9 grade scale.
Mapping QTLs for rust and LLS resistance
Phenotypic and genotypic data obtained here were used not only for MABC, but also for QTL mapping. From BC1 to BC4, the progenies were submitted to a stringent foreground selection. Therefore, most selected plants had different combinations of the four wild chromosome segments of interest, in A02, A06, B08, and A04 (with two QTLs located closely to each other), some wild genomic segments not marker-selected (“linkage drag”) and a cultivated background on most of the chromosomes. Those populations are not ideal for QTL mapping, as the plants had been marker-selected and some wild chromosome segments had been lost on all the analyzed plants. However, they were used with this aim, as an additional test to validate the association of previous identified QTLs with disease resistances, since they were present in plants of the four populations. Loci homozygous for the cultivated alleles were discarded and the remaining loci were submitted to QTL analysis using the nonparametric interval mapping based on the Kruskal–Wallis test statistic, performed in R/qtl (Broman et al., 2003). The scanone function and the argument model=“np” were used for LLS resistance, and the argument model=“binary” was used for rust resistance. We also used the model=“np” for rust resistance in BC4F2, since the plants were evaluated using the 1–9 grade scale. To increase the reliability, GWAS (genome-wide association study) was conducted using the GAPIT and FarmCPU model in R. Kinship matrix was generated using whole-genome SNPs from Affymetrix data in TASSEL. Significance threshold was calculated based on FDR value at 0.05. The BC3F3 field assay was not included on QTL analyses and GWAS due to the low number of plants assayed.
Results
Marker-assisted development of advanced lines
The induced allotetraploid MagSten was used as a donor parent in crosses with ‘IAC OL 4’ (Figure 1). Thirteen true F1 hybrids identified by microsatellite markers were crossed with ‘IAC OL 4’. A total of 201 putative BC1F1 seeds were obtained, planted in greenhouses, and genotyped with microsatellite markers linked to the five QTLs for resistance to LLS and rust. For monitoring the percentages of genome donor in each progeny, we also genotyped the 201 plants with 384 SNPs dispersed through the 10 A-genome chromosomes developed for an A. duranensis × A. stenosperma cross based on EST (Expressed Sequence Tags) data (Supplementary file 1). This was the highest coverage genotyping method available for Arachis at that time (2014). A total of 118 SNPs were informative. The proportion of wild genome ranged from 9.1% to 40.8%, with an average of 22.7% on the 201 analyzed plants. Seventeen self-pollinated plants were discarded. Thirteen BC1F1 plants were selected based on the presence of at least four of the five resistance QTLs and smaller percentage of donor (wild) A genome, ranging from 12.7% to 25.9% on the selected 13 plants. The 13 BC1F1 plants were used as male parents in backcrosses with ‘IAC OL 4’ to produce the BC2 generation. In total, 210 putative BC2F1 plants were genotyped with the microsatellite markers linked to the five QTLs. Twenty-six plants were discarded as self-pollinations and 11 were selected based on the presence of at least three of the five desired QTLs and used as male parents in backcrosses with ‘IAC OL 4’. One hundred seventy-nine putative BC3F1 plants were obtained and genotyped with the same microsatellite markers. Sixty-one true BC3F1 plants were genotyped using the Thermofisher Axiom Arachis v.01 (Clevenger et al., 2017a; Pandey et al., 2017a). Twenty-nine plants, with low genome donor (ranging from 2.3% to 12.2%), were chosen and selfed to produce BC3F2 seeds. BC3F2 plants were tested in the field, as described below, and 10 plants with LLS scores below “3”, good pod production (over 100 g/plant, not shown), and cultivated-like pod shape were selected. Slices of 30 seeds from each of the 10 plants were genotyped with microsatellite markers linked to the resistance QTLs, and seven plants (BC3F4) were backcrossed with ‘IAC OL 4’ to’ produce the BC4 generation (Figure 1). Fifty-two seeds were obtained, planted in greenhouses, and genotyped with the Axiom chip. The 44 true BC4F1 plants were selfed to produce BC4F2 seeds. Besides BC3F2, field assays were conducted on BC3F3, BC3F4, BC3F5, and BC4F2 plants. Foliar disease resistance, pod traits, and SNP genotyping data were used for the selection of plants to be advanced by selfing to the next generation.
Field assays
Plants of five generations (BC3F2, BC3F3, BC3F4, BC3F5, and BC4F2) were assayed on the field for resistance to LLS and rust, pod shape, and pod production under disease pressure. ‘IAC Sempre Verde Caiapó’, and ‘IAC OL 3’, which are resistant, partly resistant, and susceptible to both diseases, respectively, were used as controls. All populations showed genetic variation for LLS resistance and pod weight (Tables 1 and 2).
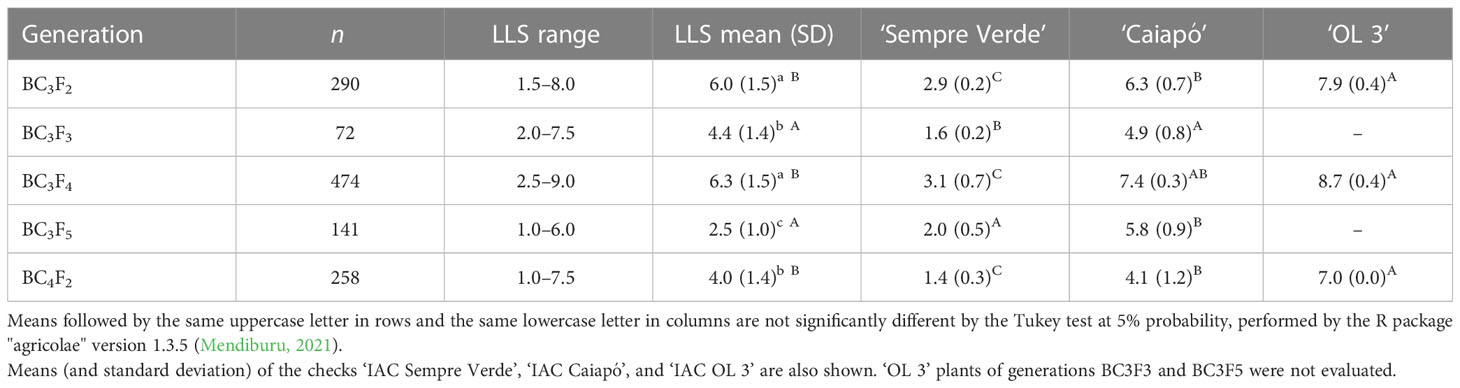
Table 1 Number of plants evaluated (n), LLS score range, and means of plants with introgressions from Arachis stenosperma and A. magna of generations BC3F2 to BC3F5, and BC4F2.

Table 2 Number of plants evaluated (n), pod weight range per plant, and mean (g) of generations BC3F2, BC3F3, BC3F5, and BC4F2 of selected plants with introgressions from Arachis stenosperma and A. magna.
A total of 290 individuals of 10 of the 12 selected BC3F2 families (two of them produced few seeds) were evaluated in 2017/2018. For comparison, 141 individuals of seven BC3F2 families that did not carry any, or only a few, of the four wild genomic segments of interest were also included. LLS scores ranged from 1.5 to 8.0, averaging 6.0 within the 10 selected families (Table 1) and ranged from 3.0 to 8.5 with an average of 7.2 within the seven check families. MagSten, ‘IAC Sempre Verde’, ‘IAC Caiapó’, and ‘IAC OL 3’ plants averaged 1.1, 2.9, 6.3, and 7.9, respectively. Rust lesions were detected in 102 out of the 290 plants from the selected families (35.2%) and on 97 out of the 141 plants of the check families (68.9%). For the controls, rust lesions were found on 24 of the 42 ‘IAC Caiapó’, 15 of the 16 ‘IAC Sempre Verde’, and all the 34 ‘OL 3’ plants. No symptoms were observed on the six MagSten plants. Population pod weight ranged from 24.0 g to 521.0 g, with an average of 157.0 g (Table 2), higher than the recurrent parent ‘OL 3’ n-significantly different from ‘IAC Sempre Verde’ and ‘IAC Caiapó’. Twenty-five plants were selected based on LLS scores (below 4.5), no symptoms of rust, the presence of most of the desired wild genomic segments, individual pod production (above 100 g per plant), and pods as similar as possible to the Runner shape. These same traits were used for the selection of plants on subsequent generations, except the LLS resistance score that varied on each trial.
Seeds of the 25 BC3F2 plants were planted in 2018/2019. A severe drought impaired plant growth and only 72 BC3F3 plants were evaluated for LLS resistance. Average LLS scores of the 72 plants ranged from 2.0 to 7.5, with an average of 4.4 (Table 1). ‘IAC Sempre Verde’ and ‘IAC Caiapó’ averaged 1.6 and 4.9, respectively. Incidence of rust was very scarce and observed on only four plants (5.6%). Despite the severe drought, pod production was relatively high, numerically comparable to the other trials: pod weight ranged from 23.2 g to 456.7 g, with an average of 167.5 g (Table 2).
Ten plants with LLS scores lower than 3.0 and the other desirable traits were selected and their seeds were planted in the field. LLS scores of the resulting 474 BC3F4 plants ranged from 2.5 to 9.0, averaging 6.3 (Table 1). MagSten, ‘IAC Sempre Verde’, ‘IAC Caiapó’, and ‘IAC OL 3’ plants averaged 1.0, 3.1, 7.4, and 8.7, respectively. Rust lesions were detected in 54.2% of the BC3F4 plants, in 17% of the ‘IAC Sempre Verde’ plants, and in all ‘IAC Caiapó’ and ‘IAC OL 3’ plants. No rust lesions were observed on the MagSten plants.
Forty BC3F4 plants were selected for having LLS scores lower than 4.5 and the other desirable traits (in this case, individual production was not weighed, but assessed visually). Seeds were planted, and 141 BC3F5 plants reached maturity and were evaluated. LLS scores ranged from 1.0 to 6.0, with an average of 2.5 and only six plants (4.3%) showed some few rust lesions (Table 1). LLS scores for ‘IAC Sempre Verde’ and ‘IAC Caiapó’ plants averaged 2.0 and 5.8, respectively. Pod weight per plant ranged from 30.1 g to 417.2 g, with an average of 185.4 g (Table 2). A total of 66 plants with LLS scores lower than 2.0, no rust lesions, with the desired wild segments, and good yield and pod/kernel traits were selected to be advanced. One of these plants was scored as 2.0 for LLS resistance, the same as ‘IAC Sempre Verde’, the highly resistant check, but it was more productive (417.2 g against 377.1 g). In addition, two plants were more resistant to LLS (scored as 1.0), with production comparable to ‘Sempre Verde’ (Supplementary file 2). None of these three plants had rust lesions, while two of the six ‘Sempre Verde’ showed some lesions.
The BC4F2 generation was planted in the field in the 2019/2020 growing season for another round of genotypic and phenotypic selection for disease resistance, in a total of 258 plants. LLS scores ranged from 1.0 to 7.5, averaging 4.0. ‘IAC Sempre Verde’, ‘IAC Caiapó’, and ‘IAC OL 3’ plants averaged 1.4, 4.1, and 7.0, respectively. This season, rust incidence enabled us to score disease severity: Rust scores averaged 1.5, and varied from 1.0 to 5.0. ‘IAC Sempre Verde’, ‘IAC Caiapó’, and ‘IAC OL 3’ plants averaged 1.4, 1.5, and 2.0, respectively. Pod weight ranged from 11.6 g to 410.2 g, with an average of 142.1 g. With this evaluation and based on the presence of the wild segments of interest, 67 plants rating 1 to 3 and with the other desired traits were selected for future evaluation and selection. Two of these plants were as resistant to LLS as ‘IAC Sempre Verde’, and one plant yielded the same while the other produced considerably more than ‘Sempre Verde’ (311.8 g against 215.6 g) (Supplementary file 2). A summary of the results of these five field assays is shown in Figure 2.

Figure 2 Minimum (blue), average (orange), and maximum (gray) LLS scores of 1,235 plants with introgressions from Arachis stenosperma and A. magna of generations BC3F2, BC3F3, BC3F4, BC3F5, and BC4F2, and average scores of the checks ‘IAC Sempre Verde’, ‘IAC Caiapó’, and ‘IAC OL’.
Mapping QTLs for LLS and rust resistance
A total of 1,204 plants of BC3F2, BC3F3, BC3F4, BC3F5, and BC4F2 generations were both field assayed and genotyped with the Thermofisher Axiom Array (Supplementary file 2). These data helped us on the selection of plants on each generation, but was also used for QTL mapping and to validate the association of previously identified QTLs with resistance against both diseases.
The skewness, kurtosis, and normality test by χ2, estimated with WinQTL Cartographer 2.5 (Wang et al., 2006), showed that the scores of LLS incidence on the four generations, as well as the rust resistance on BC4F2, were non-normally distributed (data not shown). We were unable to find any transformation of data that approximated the distributions to normality, and the nonparametric interval mapping was performed for QTL detection. The frequency distribution of plants according to their scores for LLS incidence showed bias toward susceptibility in the generations BC3F2 and BC3F4, and, in contrast, toward resistance on generations BC3F5 and BC4F2 (Figure 3). For rust severity (on BC4F2 only), the frequency distribution was strongly biased toward resistance, and 148 plants were evaluated as 1.0, and 87 as 2.0. Only 22 plants had scores ranging from 3.0 to 5.0. Pearson’s correlation between LLS scores and rust incidence was 0.624 for the 1,204 plants (not significant at 5% probability by the t-test). However, there was a clear tendency for plants more resistant to LLS to have fewer rust lesions (Supplementary file 2). The correlations between pod weight per plant and LLS scores and rust incidence were −0.177 and −0.106, respectively, both not significant at 5% probability by the t-test.

Figure 3 Frequency distribution of LLS resistance in four populations derived from 'IAC OL 4' × MagSten crosses. ‘IAC Sempre Verde’ (resistant), ‘IAC Caiapó’ (partly resistant), and ‘IAC OL 3’ (susceptible) were used as controls, and their average scores were indicated by arrows. MSD is the minimum significant difference estimated by the Tukey test at 5% probability.
On the BC3F2 assay, with 406 genotyped plants, 1,133 loci were informative, since they were heterozygous or homozygous for the wild allele. These loci were scattered throughout the 20 chromosomes. Markers associated with LLS resistance were significantly (p < 0.01) mapped in all but four chromosomes (A05, A08, B04, and B10) using the nonparametric interval mapping (Supplementary file 3). The major QTLs, detected with higher LOD scores, were located on A02, B06, A10, A06, B01, and A04 in descending order of LOD scores (Table 3). Due to the small number of plants obtained, data from the BC3F3 population was not used for QTL mapping. For the BC3F4 plants, with 380 genotyped plants, 1,002 loci scattered through 18 chromosomes had wild alleles (all the 85 loci of chromosome A08 and the 66 loci of B02 informative in BC3F2 were homozygous for the cultivated allele). Loci linked to LLS resistance were significantly (p < 0.01) identified in 14 chromosomes (A01, A02, A03, A04, A06, A10, B01, B03, B05, B06, B07, B08, B09, and B10). The QTLs with higher LOD scores were located on B06, A04, A02, A06, A03, A10, and B01. For the 134 genotyped BC3F5 plants, 602 loci scattered through 15 chromosomes were informative. In addition to A08 and B02, which lost the wild segments in BC3F4, all the loci on the chromosomes A05, A07, and B04 were homozygous for the cultivated alleles. Loci associated with LLS resistance were significantly (p < 0.01) identified in all the remaining chromosomes, except B05 (Supplementary file 3). The major QTLs, detected with higher LOD scores were located on A02, B06, A03, B01, A04, A10, and A06 (Table 3). A total of 257 BC4F2 plants, belonging to four families, were evaluated in the field for LLS and rust resistance, in 2019/2020, using a 1–9 grade scale. On the BC4F2 generation, only nine chromosomes (A02, A04, A06, A10, B01, B03, B06, B08, and B10) still had some wild alleles on their 449 informative loci. Loci linked to LLS resistance were significantly (p < 0.01) identified in six chromosomes, B06, A10, A04, B01, A06 and B10, in descending order of LOD scores. Therefore, the significant and major QTLs detected on all the four generations were located on chromosomes A04, A06, A10, B01, and B06. In addition, QTLs were identified on A02 with high LOD scores in all but the BC4F2 generation (Table 3). These six chromosome segments were considered as having the major QTLs for LLS resistance. The effects of cultivated x wild alleles of the nearest SNP of each identified QTL for LLS resistance on these six segments are shown in Figure 4. Differences were significant for all loci, according to a Kruskal–Wallis test (p < 0.01 and *p < 0.05), and show that the wild alleles significantly decrease LLS scores for all loci. Some BC3F5 plants containing the resistance chromosome segments from the wild species are shown in Figure 5, as compared to ‘IAC OL 3’. Complete QTL mapping information for the four populations is shown in Supplementary file 3.
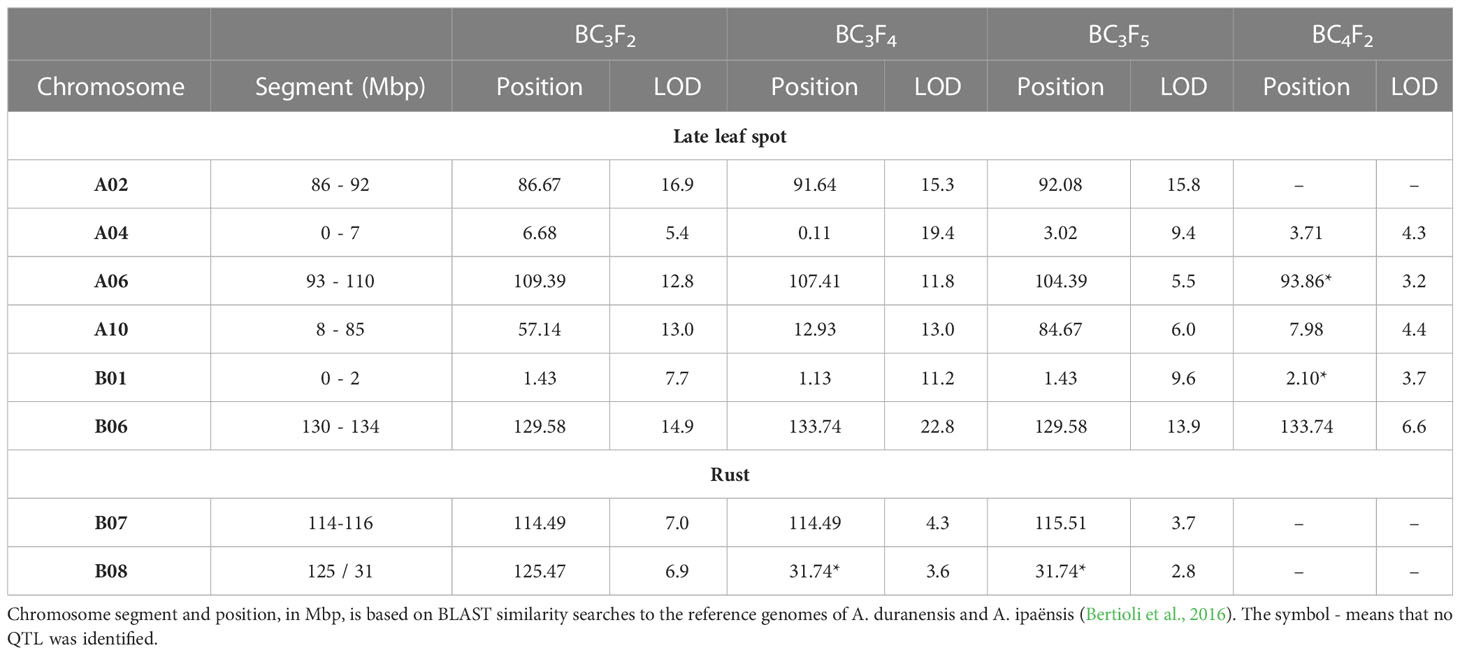
Table 3 Quantitative trait loci identified for late leaf spot and rust resistance on populations of four generations (BC3F2, BC3F4, BC3F5 and BC4F2) derived from the cross ‘IAC OL 4’ × (A. magna K 30097 × A. stenosperma V 15076)4x using the non-parametric interval mapping in R/qtl and a significance level of 1%, except the four QTLs with an asterisk (*) that were identified at 5%.
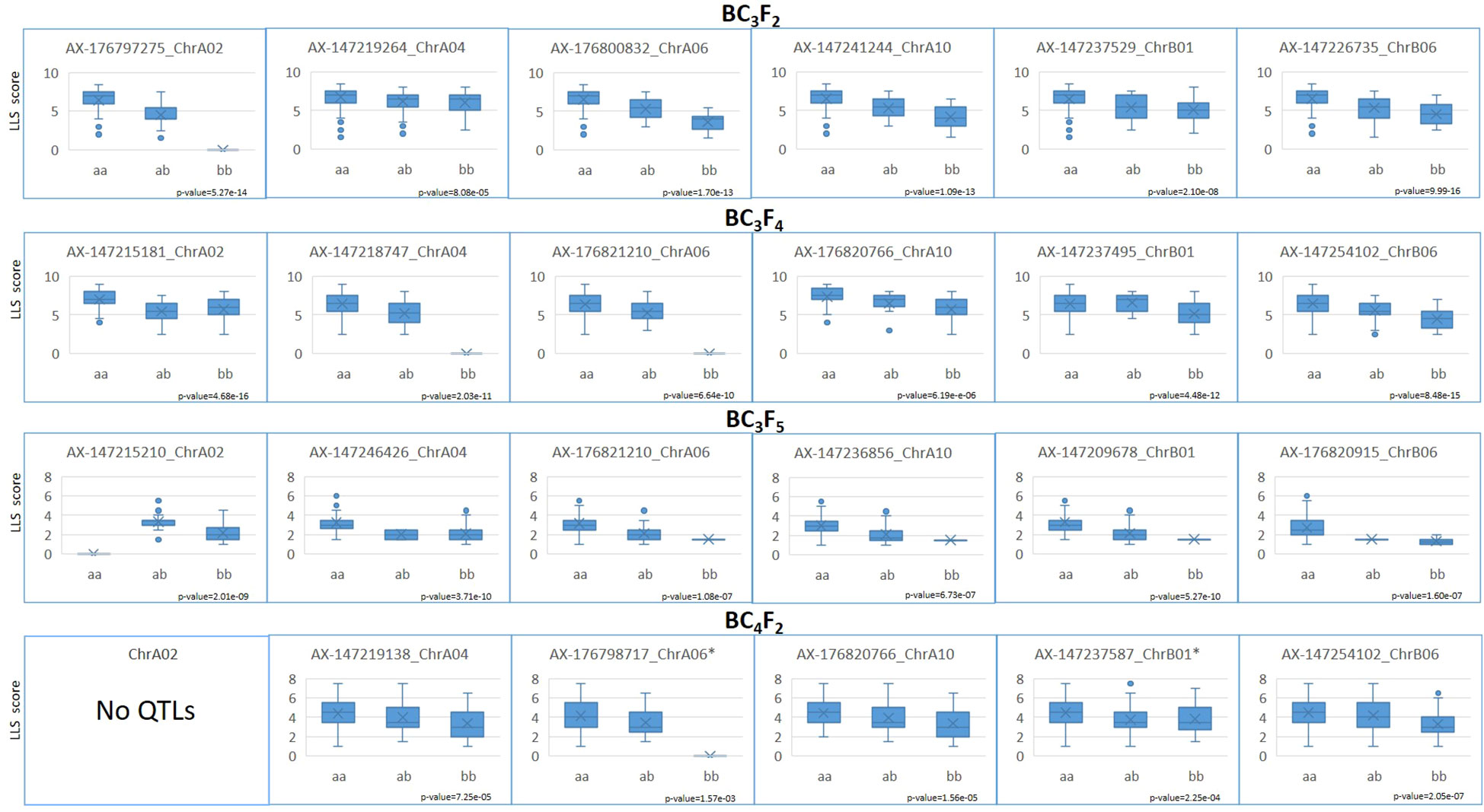
Figure 4 Boxplots showing the significant differences of LLS resistance scores between individuals homozygous for the cultivated (aa) and wild (bb) alleles, and heterozygous (ab) for the nearest loci of QTLs identified on chromosomes A02, A04, A06, A10, B01, and B06, on populations BC3F2, BC3F4, BC3F5, and BC4F2. Genotypes not observed are shown as x with a score of 0. No QTL was detected on the chromosome A02 on BC4F2.

Figure 5 Field trial of BC3F5 plants descending from ‘IAC OL 4’ × (A. magna K30097 × A. stenosperma V15076)4x. Disease pressure was high and no fungicide sprays were applied. Field at the middle of the season. Mid row is the susceptible control ‘IAC OL 3’, highly affected by LLS, and left and right rows are BC3F5 plants with chromosome segments conferring disease resistance.
GWAS analysis did not identify any significant marker on populations BC3F5 and BC4F2, but showed some significant SNPs on BC3F2 and BC3F4, especially on chromosome B06 (Figure 6). The significant markers identified by QTL mapping and GWAS analysis on the chromosome B06 are also shown in Figure 6.
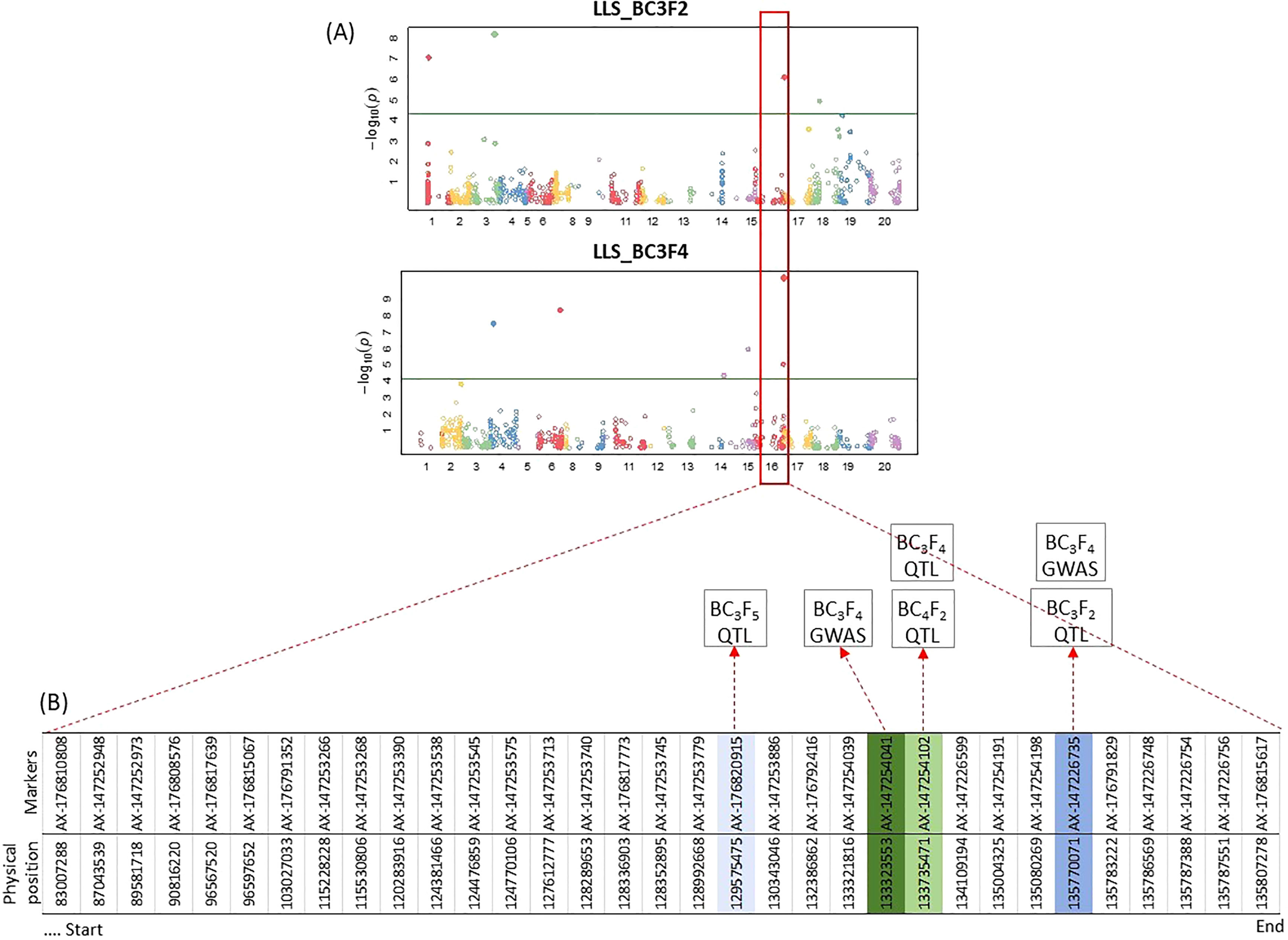
Figure 6 (A) Manhattan plot representing the results of GWAS of LLS using SNPs obtained from Axiom data for BC3F2 and BC3F4 generations. Each dot represents an SNP. The green line represents significance threshold (FDR = 0.05). No significant SNPs were identified for BC3F5 and BC4F2. (B) Distribution of significant markers identified from QTL and GWAS analyses in chromosome B06 on populations BC3F2, BC3F4, BC3F5, and BC4F2. Colored boxes of markers represent significant SNPs/QTLs from the GWAS/QTL analyses for the different generations.
For rust, QTLs were identified on 16 chromosomes in BC3F2 (except A03, A05, B01, and B04), 8 chromosomes in BC3F4 (A01, A04, A10, B04, B07, B08, B09, and B10), and 6 chromosomes in BC3F5 (A02, A03, A06, B03, B07, and B08) at a 5% significance level (Supplementary file 3). Only two common QTLs were detected, located on chromosomes B07 and B08 (Table 3). No QTLs were detected on BC4F2.
Discussion
Arachis hypogaea, the cultivated peanut, has a very narrow genetic base and few sources of resistance to the major biotic and abiotic stresses that impair the crop productivity. This is especially true for the fungal foliar diseases, since no highly resistant genotype of pure cultivated peanut has been found to date. Crop wild relatives are the reservoir of many useful genes and highly resistant Arachis wild species and accessions have been identified (reviewed by Stalker, 2017). Despite this, the use of peanut relatives in crop improvement is still incipient and has been mainly hampered by the barriers of fertility due to the ploidy differences, the transfer of undesirable genes associated with the wild genes of interest (linkage drag), and the difficulties of monitoring the wild chromosomic segments being introgressed into the cultivars.
To overcome the ploidy difference, we developed an induced allotetraploid by crossing the diploid B genome species A. magna accession K 30097 with the diploid A genome A. stenosperma accession V 15076, both highly resistant to LLS, early leaf spot, and rust (Fávero et al., 2015b; Leal-Bertioli et al., 2015a; Michelotto et al., 2015). The resulting allotetraploid also showed high resistance to LLS, early leaf spot, and rust (Michelotto et al., 2016). We used a backcross approach followed by selfings. For the efficient introgression of the genes of interest, we also identified and used microsatellite markers associated with resistance to rust in A. magna K 30097 (Leal-Bertioli et al., 2015a) and LLS in A. stenosperma V 10309 (Leal-Bertioli et al., 2009). Although this is not the same accession used in MagSten, we tentatively used and validated the markers on V 15076, which was found to be more resistant to LLS than V 10309 by Michelotto et al. (2015). Finally, for tracking the introgressed wild chromosome segments and speeding the recovery of the cultivated genome, we took advantage of the recently developed Thermofisher Axiom Arachis Arrays v.01 and v.02 (Clevenger et al., 2017a; Pandey et al., 2017a; Korani et al., 2019). These tools greatly reduced the barriers for the use of Arachis wild relatives in the peanut breeding program. We developed advanced lines containing wild chromosome segments conferring high resistance to two fungal foliar diseases, rust, and LLS. In addition, preliminary results suggest that some of them are also high-yielding lines (Figure 7).
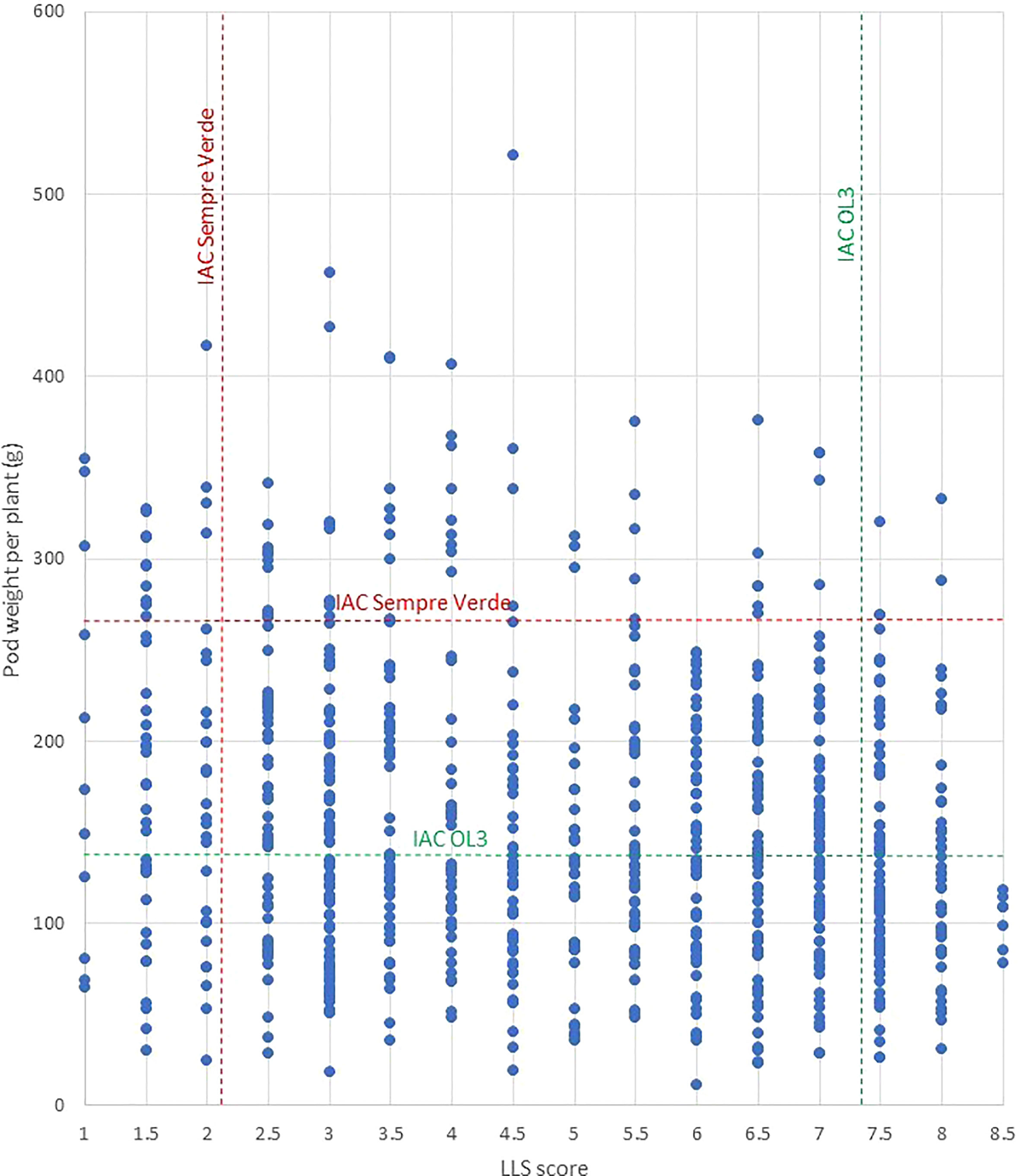
Figure 7 Pod weight produced and LLS score of each of the 808 plants of generations BC3F2, BC3F3, BC3F5, and BC4F2 and the controls ‘IAC Sempre Verde’ (red) and ‘IAC OL 3’ (green). No fungicide was sprayed on the four field assays.
Among these lines, 31 were highly resistant to LLS and productive under unsprayed fungicide conditions, and 5 of them showed resistance and production (considering the production of individual plants) similar to or higher than ‘IAC Sempre Verde’. This cultivar was released in Brazil in 2019 and incorporates very strong LLS and rust resistance (Godoy et al., 2022). Under cultivation without fungicide control of foliar diseases, it produces 5,000 kg ha−1. ‘IAC Sempre Verde’ provides the first viable option for ‘Organic’ production in the high disease pressure peanut growing environments in Brazil. During the development of ‘IAC Sempre Verde’, we discovered that its disease-resistant parent IAC 69007 has A. cardenasii chromosome segments that confer resistance to fungal foliar diseases located in chromosomes A02 and A03. Further investigations revealed similar segments in disease-resistant cultivars used in 30 countries in the Americas, Africa, Asia, and Oceania (Bertioli et al., 2021a). We consider it likely that A. cardenasii has been the main, and perhaps the only source of high resistance to fungal foliar diseases used in peanut breeding programs worldwide. Additional sources of resistance to these diseases are thus essential for the development of new peanut cultivars to reduce the risk of resistance breakdown. In this work, we identified resistances to LLS and rust provided by A. stenosperma and A. magna located mostly on different chromosomal locations from the A. cardenasii segments, providing new sources of resistance. Although QTLs were also detected on chromosome A02 of A. stenosperma, on the bottom end of the chromosome (from 86 to 92 Mbp, Table 3), they seem to be located in different segments from those of A. cardenasii that were located on the top of the chromosome (122,410 pb to 4.4 Mbp), using both the same physical map and SNP array (Lamon et al., 2020). In addition, some lines had a higher number of pods per plant than elite cultivars, indicating putative yield superiority. This will be tested in further field trials in large areas, cultivated under the recommended control of diseases for the crop. Some lines would also be tested as additional options for ‘Organic’ production, since this is a growing demand from the market in Brazil, and, more importantly, for conventional farming with reduced sprays. Utilization of these promising lines derived from wild Arachis species in peanut breeding programs will assist in developing new disease-resistant cultivars with a broader genetic base. Selected lines are also being crossed to highly resistant A. cardenasii-containing chromosome segment lines for pyramiding resistant genes, aiming at even greater resistance and durability.
QTL mapping, using the nonparametric interval mapping and 1,177 plants of BC3F2, BC3F4, BC3F5, and BC4F2, showed that six common segments had loci significantly associated with LLS resistance, located on chromosomes A02, A04, A06, A10, B01, and B06 (Table 3, Figure 4). These chromosome segments include the three segments containing the QTLs for LLS resistance previously identified in A. stenosperma V 10309, in chromosomes A02, A04, and A06 (Leal-Bertioli et al., 2009). These results corroborated and provided additional lines of evidence for the presence of major QTLs on these three chromosomes. Therefore, both accessions of A. stenosperma (V 10309 and V 15076) seem to share these LLS resistance genes. Arachis stenosperma has a peculiar disjunct distribution in Central Brazil and along the Atlantic coast, separated by more than 1,000 km. It is well known that A. stenosperma was cultivated by local people in the past, who probably promoted its migration to the coast (Krapovickas and Gregory, 1994; Custódio et al., 2005). The accession V 10309 was collected in the Mato Grosso state (Central Brazil), while V 15076 was collected in Paraná state, on the Atlantic coast. Studies have shown that all accessions of A. stenosperma studied to date are very closely related genetically (Singh et al., 2004; Koppolu et al., 2010; Moretzsohn et al., 2013) and morphologically similar (Krapovickas and Gregory, 1994). Based on this, we were confident that they should share some of the resistance segments, as corroborated by the present study.
Our QTL analyses also identified three additional chromosome segments associated with LLS resistance, on chromosomes A10, B01, and especially on B06, which showed QTLs with the highest or one of the highest LOD scores on all the four populations (Table 3). GWAS also identified significant markers on B06, very close to the mapped QTLs (Figure 6), which corroborated that the end of chromosome B06 (from 130 Mbp to 134 Mbp) on the induced allotetraploid MagSten has QTLs for resistance to LLS. Loci on A03, B07, and B09 were significantly associated on the initial generations with high LOD scores, but the wild alleles were lost on the following generations, since we were not marker-selecting segments on these chromosomes. BC3F4 or BC3F5 plants containing these segments will be rescued and advanced on our breeding schedule, for pyramiding of resistance QTLs for a more durable resistance. In addition, and as expected, several minor QTLs were detected in different chromosomes and populations (Supplementary file 3). The presence of some minor QTLs will also be monitored on the next cycles of selection.
Out of the 31 high-yielding lines, highly resistant to LLS and probably to rust, 23 have wild alleles on all the main six segments with QTLs conferring resistance to LLS, while the remaining eight lines have four or five of them (Supplementary file 2). In contrast, the most susceptible plants to LLS, considering the 1,177 genotyped and phenotyped plants of the four generations, have very few, if any, of the resistance segments. These results strongly suggest that the six chromosome segments are associated with LLS resistance, although the exact location of the resistance genes within the segments is still unknown. Additional evidence for the identified wild resistance segments was the clear decreasing of LLS scores on the successive BC3 generations, which were advanced by using a stringent foreground selection (Figures 2, 3). The exception was BC3F4, which showed the higher minimum, average, and maximum disease values. In the 2018/2019 season, when the BC3F4 plants were evaluated, an early incidence of LLS occurred. Therefore, despite the plants being evaluated at approximately 120 days after sowing, as all the other generations, the disease damage was considerably higher than the other trials. Many ‘IAC OL 3’ plants were dead and LLS scores of 'IAC Sempre Verde' and ‘IAC Caiapó’ were higher than the values observed on the other seasons (Table 1).
Rust is a sporadic disease, and its incidence was very weak in the four BC3 field assays. Therefore, plants were evaluated by the presence (1) or absence (0) of rust lesions. In BC4F2, plants could be evaluated using the 1–9 grade scale. However, the incidence was only moderate and 228 out of 257 plants (88.7%) had scores lower than 2.0. Probably due to this low variation, no QTL was detected. On BC3, several QTLs were identified, but only two QTLs were significantly associated with all the three populations analyzed, using the nonparametric interval mapping and the argument model=“binary”, at p < 0.05 (Table 3). These QTLs were located on chromosomes B07 and B08. The segment on B07 was well defined, located within 114 and 116 Mbp, and the QTL with higher LOD scores on the three BC3 populations (Table 3; Supplementary file 3). The end of chromosome B08, where the QTL was identified on BC3F2 (at p < 0.01) coincides with the QTL previously identified for A. magna K 30097 on a diploid cross (Leal-Bertioli et al., 2015a). However, the QTLs were detected (at p < 0.05) close to 32 Mbp on BC3F4 and BC3F5. Therefore, the location of QTLs for rust resistance in A. magna still needs further investigation.
Very few QTLs for resistance to LLS or rust have been mapped in Arachis wild species. In contrast, a number of QTLs for resistance to both diseases have been mapped in cultivated peanut using A. hypogaea × A. hypogaea crosses, despite two out of the three sources of resistance used on these studies having wild species on their pedigrees. Using the A. cardenasii-derived GBPD4 as the source of resistance, the number of QTLs ranged from 4 to 28 for LLS resistance, and from 5 to 15 for rust resistance (Khedikar et al., 2010; Sujay et al., 2012; Kolekar et al., 2016). As expected, most of them are minor QTLs. However, two major and common QTL regions were detected, located on chromosomes A03 for LLS and rust resistance, and A02 for LLS resistance only. These two candidate genomic regions conferring resistance to LLS and rust have been validated and used on marker-assisted selection for developing foliar disease-resistant lines (Yeri and Bhat, 2016; Kolekar et al., 2017; Pandey et al., 2017b; Ahmad et al., 2020). It is now well established that both resistance regions came from A. cardenasii accession GKP 10017 (Bertioli et al., 2021a). QTL mapping studies using Tifrunner as the source of LLS resistance identified up to 22 QTLs, but three consistent QTLs, located on chromosomes A05, B03 and B05, were mapped and validated (Pandey et al., 2017c; Clevenger et al., 2018; Chu et al., 2019). Finally, a study using as the source of resistance the ICGV 86699 line, which is reported to be a derivative of A. batizocoi/A. duranensis × A. hypogaea “NC 2” crosses (Reddy et al., 1996), but which was found to have A. cardenasii segments from chromosomes A02 and A03 by Bertioli et al. (2021a), identified two major QTLs for LLS resistance on chromosomes B06 and A10 (Zhou et al., 2016). Therefore, from the six wild chromosome segments containing LLS resistance genes identified and introgressed into peanut here, only three were located on the same chromosomes of previously mapped QTLs: A02 from A. cardenasii, and A10 and B06, whose origin is unclear. At least the other three segments are new and have never been used by any breeding program. The co-localization or not of QTLs on chromosomes A02, A10, and B06 needs further investigation. For rust resistance, the present study provided additional lines of evidence for the presence of resistance genes on the chromosome B08. The two segments identified here (on chromosomes B07 and B08) do not coincide with the only other consistent QTL for rust resistance mapped to date, located on chromosome A03, from A. cardenasii.
Here, we showed the efficient introgression of wild segments with resistance genes against two important diseases into an elite high oleic peanut cultivar from the secondary gene pool. The approach used in this study provides a way to expand the genetic base of cultivated peanut by exploiting genetic variability present in wild species, as well as ensuring a continuous supply of new sources of resistance to different biotic and abiotic stresses. As an example, the accession V 15076 of A. stenosperma was collected in the pure sand of a beach, very close to the sea, where it was found growing vigorously and producing many flowers (Figure 8) and seeds. This accession may also have tolerance to salinity, drought, and heat, among other stresses. Therefore, the material produced in the present study might also be important for the development of climate-resilient plants. Finally, we showed that genetic limits of a complex trait such as yield were also overcome during the introgression of segments from crop wild relatives.

Figure 8 The accession A. stenosperma V 15076 growing in the place where it was collected, in Caiobá Beach, Paraná state. Note the pure sand where the plants were growing vigorously and the very close proximity to the sea. The photo was taken during its collection and kindly provided by the collector José Valls.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author contributions
MCM, SCML-B, IJG, and DJB conceived the study. JFS, ARAM, MDM, and IJG conducted the field experiments. MCM, ARC, SCML-B, and DJB performed the molecular data analyses. MCM and JFS performed the QTL and statistical analyses. NM carried out GWAS analysis. The first draft of the manuscript was written by MCM, and all authors commented on previous versions of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by Mars Wrigley Inc. and the host institutions. Author MCM has received a Research Productivity Fellowship (Proc. 310707/2019-6) from the National Council for Scientific and Technological Development (CNPq). The authors declare that no funds, grants, or other support were received during the preparation of this manuscript.
Acknowledgments
The authors thank Alessandra Fávero for allotetraploid material and for making initial crosses. We also thank the National Council for Scientific and Technological Development (CNPq) for MCM Research Productivity Fellowship (Proc. 310707/2019-6). This work was funded by Mars Wrigley Inc. and the host institutions.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This study received funding from Mars Wrigley Inc. The funder was not involved in the study design, collection, analysis, interpretation of data, the writing of this article or the decision to submit it for publication.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2023.1139361/full#supplementary-material
References
Ahmad, S., Nawade, B., Sangh, C., Mishra, G. P., Bosamia, T. C., Radhakrishnan, T., et al. (2020). Identification of novel QTLs for late leaf spot resistance and validation of a major rust QTL in peanut (Arachis hypogaea l.). 3 Biotech. 10, 1–13. doi: 10.1007/s13205-020-02446-4
Backman, P. A., Crawford, M. A. (1984). Relationship between yield loss and severity of early and late leaf spot diseases of peanut (Arachis hypogaea l.). Phytopathology 74, 1101–1103. doi: 10.1094/Phyto-74-1101
Ballén-Taborda, C., Chu, Y., Ozias-Akins, P., Timper, P., Holbrook, C. C., Jackson, S. A., et al. (2019). A new source of root-knot nematode resistance from Arachis stenosperma incorporated into allotetraploid peanut (Arachis hypogaea). Sci. Rep. 9, 1–13. doi: 10.1038/s41598-019-54183-1
Bertioli, D. J., Cannon, S. B., Froenicke, L., Huang, G., Farmer, A. D., Cannon, E. K., et al. (2016). The genome sequence of Arachis duranensis and Arachis ipaënsis, the diploid ancestors of cultivated peanut.). Nat. Genet. 48, 438–446. doi: 10.1038/ng.3517
Bertioli, D. J., Clevenger, J., Godoy, I. J., Stalker, H. T., Wood, S., Santos, J. F., et al. (2021a). Legacy genetics of Arachis cardenasii in the peanut crop shows the profound benefits of international seed exchange. Proc. Natl. Acad. Sci. U. S. A. 118, 1–9. doi: 10.1073/pnas.2104899118
Bertioli, D. J., Gao, D., Ballen-Taborda, C., Chu, Y., Ozias-Akins, P., Jackson, S. A., et al. (2021b). Registration of GA-BatSten1 and GA-MagSten1, two induced allotetraploids derived from peanut wild relatives with superior resistance to leaf spots, rust, and root-knot nematode. J. Plant Regist. 15, 1–7. doi: 10.1002/plr2.20133
Bharadwaj, C., Jorben, J., Rao, A., Roorkiwal, M., Patil, B. S., Jayalakshmi, et al. (2022). Development of high yielding fusarium wilt resistant cultivar by pyramiding of "Genes" through marker-assisted backcrossing in chickpea (Cicer arietinum l.). Front. Genet. 13. doi: 10.3389/fgene.2022.924287
Broman, K. W., Wu, H., Sen, S., Churchill., G. A. (2003). R/qtl: QTL mapping in experimental crosses. Bioinform 19 (7), 889–890. doi: 10.1093/bioinformatics/btg112
Burow, M. D., Simpson, C. E., Starr, J. L., Paterson, A. H. (2001). Transmission genetics of chromatin from a synthetic amphidiploid to cultivated peanut (Arachis hypogaea l.): Broadening the gene pool of a monophyletic polyploid species. Genetics 159, 823–837. doi: 10.1093/genetics/159.2.823
Chu, Y., Chee, P., Culbreath, A., Isleib, T. G., Holbrook, C. C., Ozias-Akins, P. (2019). Major QTLs for resistance to early and late leaf spot diseases are identified on chromosomes 3 and 5 in peanut (Arachis hypogaea). Front. Plant Sci. 10. doi: 10.3389/fpls.2019.00883
Chu, Y., Holbrook, C. C., Timper, P., Ozias-Akins, P. (2007). Development of a PCR-based molecular marker to select for nematode resistance in peanut. Crop Sci. 47, 841–847. doi: 10.2135/cropsci2006.07.0474
Chu, Y., Wu, C. L., Holbrook, C. C., Tillman, B. L., Person, G., Ozias-Akins, P. (2011). Marker-assisted selection to pyramid nematode resistance and the high oleic trait in peanut. Plant Genome J. 4, 110. doi: 10.3835/plantgenome2011.01.0001
Clevenger, J., Chu, Y., Arrais Guimaraes, L., Maia, T., Bertioli, D., Leal-Bertioli, S., et al. (2017b). Gene expression profiling describes the genetic regulation of Meloidogyne arenaria resistance in Arachis hypogaea and reveals a candidate gene for resistance. Sci. Rep. 7, 1317. doi: 10.1038/s41598-017-00971-6
Clevenger, J., Chu, Y., Chavarro, C., Agarwal, G., Bertioli, D. J., Leal-Bertioli, S. C. M., et al. (2017a). Genome-wide SNP genotyping resolves signatures of selection and tetrasomic recombination in peanut. Mol. Plant 10, 309–322. doi: 10.1016/j.molp.2016.11.015
Clevenger, J., Chu, Y., Chavarro, C., Botton, S., Culbreath, A., Isleib, T. G., et al. (2018). Mapping late leaf spot resistance in peanut (Arachis hypogaea) using QTL-seq reveals markers for marker-assisted selection. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.00083
CONAB - Companhia Nacional de Abastecimento (2023). Available at: https://www.conab.gov.br/info-agro/safras/serie-historica-das-safras/itemlist/category/899-amendoim (Accessed 10 January 2023).
Coutinho, W. M., Suassuna, N. D. (2014). “Manejo integrado de doenças,” in Sistema de produção de amendoim, 2. ed. Ed. Suassuna, T. M. F. (Campina Grande: Embrapa Algodão). Available at: https://www.spo.cnptia.embrapa.br/temas-publicados.
Custódio, A. R., Peñaloza, A. P. S., Valls, J. F. M. (2005). Further cytogenetic information on Arachis stenosperma (Leguminosae). Cytologia (Tokyo). 70, 331–335. doi: 10.1508/cytologia.70.331
Custódio, A. R., Seijo, G., Valls, J. F. M. (2013). Characterization of Brazilian accessions of wild Arachis species of section Arachis (Fabaceae) using heterochromatin detection and fluorescence in situ hybridization (FISH). Genet. Mol. Biol. 36, 364–370. doi: 10.1590/S1415-47572013000300011
FAOSTAT – Food and Agriculture Organization of the United Nations (2022). Available at: https://www.fao.org/faostat/en/#data/QCL (Accessed 10 January 2023).
Fávero, A. P., Pádua, J. G., Costa, T. S., Gimenes, M. A., Godoy, I. J., Moretzsohn, M. C., et al. (2015b). New hybrids from peanut (Arachis hypogaea l.) and synthetic amphidiploid crosses show promise in increasing pest and disease tolerance. Genet. Mol. Res. 14, 16694–16703. doi: 10.4238/2015.December.11.17
Fávero, A. P., Santos, R. F., Simpson, C. E., Valls, J. F. M., Vello, N. A. (2015a). Successful crosses between fungal-resistant wild species of Arachis (Section Arachis) and Arachis hypogaea. Genet. Mol. Biol. 38, 353–365. doi: 10.1590/S1415-475738320140376
Fávero, A. P., Simpson, C. E., Valls, J. F. M., Vello, N. A. (2006). Study of the evolution of cultivated peanut through crossability studies among Arachis ipaënsis, a. duranensis, and A. hypogaea. Crop Sci. 46, 1546–1552. doi: 10.2135/cropsci2005.09-0331
Fernandez, A., Krapovickas, A. (1994). Cromosomas y evolución en Arachis (Leguminosae). Bonplandia 8, 187–220. doi: 10.30972/bon.81-41499
Frisch, M., Melchinger, A. E. (2005). Selection theory for marker-assisted back-crossing. Genetics 170, 909–917. doi: 10.1534/genetics.104.035451
Gao, D., Araujo, A. C. G., Nascimento, E. F. M. B., Chavarro, M. C., Xia, H., Jackson, S. A., et al. (2021). ValSten: A new wild species derived allotetraploid for increasing genetic diversity of the peanut crop (Arachis hypogaea l.). Genet. Resour. Crop Evol. 68, 1471–1485. doi: 10.1007/s10722-020-01076-2
Godoy, I. J., Santos, J. F., Carvalho, C. R. L., Michelotto, M. D., Bolonhezi, D., Freitas, R. S., et al. (2014). IAC OL 3 and IAC OL 4: New Brazilian peanut cultivars with the high oleic trait. Crop Breed. Appl. Biotechnol. 14, 200–203. doi: 10.1590/1984-70332014v14n3a30
Godoy, I. J., Santos, J. F., Moretzsohn, M. C., Moraes, A. R. A., Michelotto, M. D., Bolonhezi, D., et al. (2022). ‘IAC SEMPRE VERDE’: A wild-derived peanut cultivar highly resistant to foliar diseases. Crop Breed. Appl. Biotechnol. 22, e41252232. doi: 10.1590/1984-70332022v22n3c25
Holbrook, C. C., Ozias-Akins, P., Chu, Y., Lamon, S., Bertioli, D. J., Leal-Bertioli, S. C., et al. (2022). Registration of TifGP-3 and TifGP-4 peanut germplasm lines. J. Plant Regist. 16, 120–123. doi: 10.1002/plr2.20179
Khedikar, Y. P., Gowda, M. V. C., Sarvamangala, C., Patgar, K. V., Upadhyaya, H. D., Varshney, R. A. (2010). QTL study on late leaf spot and rust revealed one major QTL for molecular breeding for rust resistance in groundnut (Arachis hypogaea l.). K Theor. Appl. Genet. 121, 971–984. doi: 10.1007/s00122-010-1366-x
Kim, M. S., Yu, J. K., Ko, S. R., Kim, K. J., Ji, H., Kang, K. K., et al. (2022). Marker-assisted backcrossing (MABc) to improve eating quality with thin seed coat and aleurone layer of non-glutinous japonica variety in rice. Genes 13, 210. doi: 10.3390/genes13020210
Kolekar, R. M., Sujay, V., Shirasawa, K., Sukruth, M., Khedikar, Y. P., Gowda, M. V. C., et al. (2016). QTL mapping for late leaf spot and rust resistance using an improved genetic map and extensive phenotypic data on a recombinant inbred line population in peanut (Arachis hypogaea l.). Euphytica 209, 147–156. doi: 10.1007/s10681-016-1651-0
Kolekar, R. M., Sukruth, M., Shirasawa, K., Nadaf, H. L., Motagi, B. N., Lingaraju, S., et al. (2017). Marker-assisted backcrossing to develop foliar disease-resistant genotypes in TMV 2 variety of peanut (Arachis hypogaea l.). Plant Breed. 136, 948–953. doi: 10.1111/pbr.12549
Koppolu, R., Upadhyaya, H. D., Dwivedi, S. L., Hoisington, D. A., Varshney, R. K. (2010). Genetic relationships among seven sections of genus Arachis studied by using SSR markers. BMC Plant Biol. 10, 15. doi: 10.1186/1471-2229-10-15
Korani, W., Clevenger, J. P., Chu, Y., Ozias-Akins, P. (2019). Machine learning as an effective method for identifying true single nucleotide polymorphisms in polyploid plants. Plant Genome 12, 1–10. doi: 10.3835/plantgenome2018.05.0023
Krapovickas, A., Gregory, W. C. (1994). Taxonomia del genero Arachis (Leguminosae). Bonplandia 8, 1–186.
Kumari, V., Gowda, M. V. C., Tasiwal, V., Pandey, M. K., Bhat, R. S., Mallikarjuna, N., et al. (2014). Diversification of primary gene pool through introgression of resistance to foliar diseases from synthetic amphidiploids to cultivated groundnut (Arachis hypogaea l.). Crop J. 2, 110–119. doi: 10.1016/j.cj.2014.03.002
Lamon, S., Chu, Y., Guimaraes, L. A., Bertioli, D. J., Leal-Bertioli, S. C. M., Santos, J. F., et al. (2020). Characterization of peanut lines with interspecific introgressions conferring late leaf spot resistance. Crop Sci. 61, 1724–1738. doi: 10.1002/csc2.20414
Lavia, G. I., Ortiz, A. M., Fernández, A. (2009). Karyotypic studies in wild germplasm of Arachis (Leguminosae). Genet. Resour. Crop Evol. 56, 755–764. doi: 10.1007/s10722-008-9399-6
Leal-Bertioli, S. C. M., Cavalcante, U., Gouvea, E. G., Ballén-Taborda, C., Shirasawa, K., Guimarães, P. M., et al. (2015a). Identification of QTLs for rust resistance in the peanut wild species Arachis magna and the development of KASP markers for marker-assisted selection. G3; Genes|Genomes|Genetics 5, 1403–1413. doi: 10.1534/g3.115.018796
Leal-Bertioli, S. C. M., José, A. C. V. F., Alves-Freitas, D. M. T., Moretzsohn, M. C., Guimarães, P. M., Nielen, S., et al. (2009). Identification of candidate genome regions controlling disease resistance in Arachis. BMC Plant Biol. 9, 112. doi: 10.1186/1471-2229-9-112
Leal-Bertioli, S. C. M., Moretzsohn, M. C., Roberts, P. A., Ballén-Taborda, C., Borba, T. C. O., Valdisser, P. A., et al. (2016). Genetic mapping of resistance to Meloidogyne arenaria in Arachis stenosperma: A new source of nematode resistance for peanut. G3; Genes|Genomes|Genetics 6, 377–390. doi: 10.1534/g3.115.023044
Leal-Bertioli, S. C. M., Moretzsohn, M. C., Santos, S. P., Brasileiro, A. C. M., Guimarães, P. M., Bertioli, D. J., et al. (2017). Phenotypic effects of allotetraploidization of wild Arachis and their implications for peanut domestication. Am. J. Bot. 104, 379–388. doi: 10.3732/ajb.1600402
Leal-Bertioli, S. C. M., Santos, S. P., Dantas, K. M., Inglis, P. W., Nielen, S., Araujo, A. C. G., et al. (2015b). Arachis batizocoi: A study of its relationship to cultivated peanut (A. hypogaea) and its potential for introgression of wild genes into the peanut crop using induced allotetraploids. Ann. Bot. 115, 237–249. doi: 10.1093/aob/mcu237
Mendiburu, F. G. Agricolae: Statistical Procedures for Agricultural Research. R package version 1.3.5. Available at: https://CRAN.R-project.org/package=agricolae.
Michelotto, M. D., Barioni, W., Resende, M. D. V., Godoy, I. J., Leonardecz, E., Fávero, A. P. (2015). Identification of fungus resistant wild accessions and interspecific hybrids of the genus Arachis. PloS One 10, 1–17. doi: 10.1371/journal.pone.0128811
Michelotto, M. D., Godoy, I. J., Santos, J. F., Martins, A. L. M., Leonardecz, E., Fávero, A. P. (2016). Identifying Arachis amphidiploids resistant to foliar fungal diseases. Crop Sci. 56, 1792–1798. doi: 10.2135/cropsci2015.06.0393
Moraes, A. S., Godoy, I. J. (1997). “Amendoim - controle de doenças,” in Controle de doenças de plantas: Grandes culturas, vol. 1 . Eds. Zambolim, L., Vale, F. X. R. (Viçosa, Universidade Federal de Viçosa; Brasília, Ministério da Agricultura e do Abastecimento), 1–49.
Moretzsohn, M. C., Gouvea, E. G., Inglis, P. W., Leal-Bertioli, S. C. M., Valls, J. F. M., Bertioli, D. J. (2013). A study of the relationships of cultivated peanut (Arachis hypogaea) and its most closely related wild species using intron sequences and microsatellite markers. Ann. Bot. 111, 113–126. doi: 10.1093/aob/mcs237
Pandey, M. K., Agarwal, G., Kale, S. M., Clevenger, J., Nayak, S. N., Sriswathi, M., et al. (2017a). Development and evaluation of a high density genotyping “Axiom-Arachis” array with 58 K SNPs for accelerating genetics and breeding in groundnut. Sci. Rep. 7, 1–10. doi: 10.1038/srep40577
Pandey, M. K., Khan, A. W., Singh, V. K., Vishwakarma, M. K., Shasidhar, Y., Kumar, V., et al. (2017b). QTL-seq approach identified genomic regions and diagnostic markers for rust and late leaf spot resistance in groundnut (Arachis hypogaea l.). Plant Biotechnol. J. 15, 927–941. doi: 10.1111/pbi.12686
Pandey, M. K., Wang, H., Khera, P., Vishwakarma, M. K., Kale, S. M., Culbreath, A. K., et al. (2017c). Genetic dissection of novel QTLs for resistance to leaf spots and tomato spotted wilt virus in peanut (Arachis hypogaea l.). Front. Plant Sci. 8. doi: 10.3389/fpls.2017.00025
Reddy, L. J., Nigam, S. N., Moss, J. P., Singh, A. K., Subrahmanyam, P., McDonald, D., et al. (1996). Registration of ICGV 86699 peanut germplasm line with multiple disease and insect resistance. Crop Sci. 36, 821. doi: 10.2135/cropsci1996.0011183X003600030072x
Robledo, G., Seijo, G. (2010). Species relationships among the wild b genome of Arachis species (section Arachis) based on FISH mapping of rDNA loci and heterochromatin detection: A new proposal for genome arrangement. Theor. Appl. Genet. 121, 1033–1046. doi: 10.1007/s00122-010-1369-7
Seijo, G. J., Atahuachi, M., Simpson, C. E., Krapovickas, A. (2021). Arachis inflata: A new b genome species of Arachis (Fabaceae). Bonplandia 30, 1–6. doi: 10.30972/bon.3024942
Shasidhar, Y., Variath, M. T., Vishwakarma, M. K., Manohar, S. S., Gangurde, S. S., Sriswathi, M., et al. (2020). Improvement of three popular Indian groundnut varieties for foliar disease resistance and high oleic acid using SSR markers and SNP array in marker-assisted backcrossing. Crop J. 8, 1–15. doi: 10.1016/J.CJ.2019.07.001
Shokes, F. M., Culbreath, A. K. (1997). “Early and late leaf spots,” in Compendium of peanut diseases, 2nd ed. Eds. Kokalis-Burelle, N., Porter, D. M., Rodríguez-Kábana, R., Smith, D. H., Subrahmanyam, P. (St Paul, MN, USA: The American Phytopathological Society). APS Press.
Silvestri, M. C., Ortiz, A. M., Lavia, G. I. (2015). rDNA loci and heterochromatin positions support a distinct genome type for “x = 9 species” of section Arachis (Arachis, leguminosae). Plant Syst. Evol. 301, 555–562. doi: 10.1007/s00606-014-1092-y
Simpson, C. E. (1991). Pathways for introgression of pest resistance into Arachis hypogaea l. Peanut Sci. 18, 22–26. doi: 10.3146/i0095-3679-18-1-8
Simpson, C. E., Krapovickas, A., Valls, J. F. M. (2001). History of Arachis including evidence of A. hypogaea l. progenitors. Peanut Sci. 28, 78–80. doi: 10.3146/i0095-3679-28-2-7
Simpson, C. E., Starr, J. L., Baring, M. R., Burow, M. D., Cason, J. M., Wilson, J. N. (2013). Registration of ‘Webb’ peanut. J. Plant Regist. 7, 265–268. doi: 10.3198/jpr2013.01.0005crc
Singh, M. P., Erickson, J. E., Boote, K. J., Tillman, B. J., Jones, J. W., van Bruggen, A. H. C. (2011). Late leaf spot effects on growth, photosynthesis, and yield in peanut cultivars of differing resistance. Agron. J. 103, 85–91. doi: 10.2134/agronj2010.0322
Singh, A. K., Smartt, J., Singh, R. (2004). Variation studies in a wild groundnut species, Arachis stenosperma krapov. & W.C. Gregory nov. sp. Plant Genet. Resour. 2, 99–106. doi: 10.1079/PGR200437
Smartt, J., Gregory, W. C., Gregory, M. P. (1978). The genomes of Arachis hypogaea. 1. cytogenetic studies of putative genome donors. Euphytica 27, 665–675. doi: 10.1007/BF00023701
Stalker, H. T. (1991). A new species in section Arachis of peanuts with a d genome. Am. J. Bot. 78, 630–637. doi: 10.1002/j.1537-2197.1991.tb12587.x
Stalker, H. T. (2017). Utilizing wild species for peanut improvement. Crop Sci. 57, 1102–1120. doi: 10.2135/cropsci2016.09.0824
Subrahmanyam, P., Mcdonald, D., Waliyar, F., Reddy, L. J., Nigam, S. N., Gibbons, R. W., et al. (1995). Screening methods and sources of resistance to rust and late leaf spot of groundnut. Inf. Bull. 47, 1–26. Patancheru: ICRISAT.
Sujay, V., Gowda, M. V. C., Pandey, M. K., Bhat, R. S., Khedikar, Y. P., Nadaf, H. L., et al. (2012). Quantitative trait locus analysis and construction of consensus genetic map for foliar disease resistance based on two recombinant inbred line populations in cultivated groundnut (Arachis hypogaea l.). Mol. Breed. 30, 773–788. doi: 10.1007/s11032-011-9661-z
Valls, J. F. M., Costa, L. C., Custódio, A. R. (2013). A novel trifoliolate species of Arachis (Fabaceae) and further comments on taxonomic section trierectoides. Bonplandia 22, 91–97. doi: 10.30972/bon.2211257
Valls, J. F. M., Simpson, C. E. (2005). New species of Arachis (Leguminosae) from Brazil, Paraguay and Bolivia. Bonplandia 14, 35–63. doi: 10.30972/bon.141-21387
Valls, J. F. M., Simpson, C. E. (2017). A new species of Arachis (Fabaceae) from mato grosso, Brazil, related to Arachis matiensis. Bonplandia 26, 143–149. doi: 10.30972/bon.2622575
Varshney, R. K., Pandey, M. K., Janila, P., Nigam, S. N., Sudini, H., Gowda, M. V. C., et al. (2014). Marker−assisted introgression of a QTL region to improve rust resistance in three elite and popular varieties of peanut (Arachis hypogaea l.). Theor. Appl. Genet. 127, 1771–1781. doi: 10.1007/s00122-014-2338-3
Wang, S., Basten, C. J., Zeng, Z. B. (2006). Windows QTL cartographer 2.5. department of statistics (Raleigh, NC: North Carolina State University).
Yeri, S. B., Bhat, R. S. (2016). Development of late leaf spot and rust resistant backcross lines in jl 24 variety of groundnut (Arachis hypogaea l.). Electron. J. Plant Breed. 7, 37–41. doi: 10.5958/0975-928X.2016.00005.3
Keywords: Arachis stenosperma, A. magna, induced allotetraploid, late leaf spot, rust, molecular breeding
Citation: Moretzsohn MdC, Santos JFd, Moraes ARA, Custódio AR, Michelotto MD, Mahrajan N, Leal-Bertioli SCdM, Godoy IJ and Bertioli DJ (2023) Marker-assisted introgression of wild chromosome segments conferring resistance to fungal foliar diseases into peanut (Arachis hypogaea L.). Front. Plant Sci. 14:1139361. doi: 10.3389/fpls.2023.1139361
Received: 06 January 2023; Accepted: 20 February 2023;
Published: 17 March 2023.
Edited by:
Rajib Roychowdhury, Volcani Center, IsraelReviewed by:
Sunil S. Gangurde, University of Georgia, United StatesNarayanan Manikanda Boopathi, Tamil Nadu Agricultural University, India
Soumya Prakash Das, Seacom Skills University, India
Copyright © 2023 Moretzsohn, Santos, Moraes, Custódio, Michelotto, Mahrajan, Leal-Bertioli, Godoy and Bertioli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Márcio de Carvalho Moretzsohn, marcio.moretzsohn@embrapa.br