 Haiyue Gong1
Haiyue Gong1 Xiaozhong Lan
Xiaozhong Lan Zhihua Liao
Zhihua Liao- 1School of Life Sciences, Integrative Science Center of Germplasm Creation in Western China (CHONGQING) Science City and Southwest University, The Provincial and Ministerial Co-founded Collaborative Innovation Center for R&D in Tibet Characteristic Agricultural and Animal Husbandry Resources, Southwest University, Chongqing, China
- 2Chongqing Academy of Science and Technology, Chongqing, China
- 3Xizang Agricultural and Husbandry College, The Provincial and Ministerial Co-founded Collaborative Innovation Center for R&D in Tibet Characteristic Agricultural and Animal Husbandry Resources, The Center for Xizang Chinese (Tibetan) Medicine Resource, TAAHC-SWU Medicinal Plant Joint R&D Centre, Nyingchi, China
Hyoscyamine and scopolamine, belonging to medicinal tropane alkaloids (MTAs), are potent anticholinergic drugs. Their industrial production relies on medicinal plants, but the levels of the two alkaloids are very low in planta. Engineering the MTA’s production is an everlasting hot topic for pharmaceutical industry. With understanding the MTA’s biosynthesis, biotechnological approaches are established to produce hyoscyamine and scopolamine in an efficient manner. Great advances have been obtained in engineering MTA’s production in planta. In this review, we summarize the advances on the biosynthesis of MTAs and engineering the MTA’s production in hairy root cultures, as well in plants. The problems and perspectives on engineering the MTA’s production are also discussed.
Introduction
Medicinal plants constitute a treasure for human beings to obtain diverse drugs, flavors and fine chemicals. Of them, medicinal plants of Solanaceae produce a kind of specialized metabolites named tropane alkaloids (TAs), some of which have potent anticholinergic activity. Hyoscyamine and scopolamine are well-known medicinal tropane alkaloids (MTAs) and anticholinergic drugs (Grynkiewicz and Gadzikowska, 2008). They are clinically used to treat asthma, pain, motion sickness, functional gastrointestinal disorders, Parkinson’s syndrome and etc. (Huang et al., 2021). Furthermore, hyoscyamine is the key material for industrially producing ipratropium bromide, and scopolamine for tiotropium bromide. Ipratropium bromide, as well as tiotropium bromide, is an essential drug in the treatment of chronic obstructive pulmonary disease (COPD; Griffin et al., 2008). Therefore, the demand for the two alkaloids is huge. However, their production is limited due to low levels in medicinal plants producing TAs. To develop efficient ways to manufacture hyoscyamine and scopolamine, the scientists have been trying their best. Chemists successfully synthesized them, but failed in market because chemical synthesis was not economically availed (Grynkiewicz and Gadzikowska, 2008). Then, the final solution relies on biotechnological approaches, such as metabolic engineering and synthetic biology.
Two amino acids, including ornithine and phenylalanine, are the starting precursors for the two MTAs, hyoscyamine, and scopolamine. The intact biosynthetic pathway of MTAs has been elucidated, which is composed of 13 biosynthesis enzymes (Figure 1). Such MTA’s biosynthesis enzymes and their corresponding genes provide valuable tools to engineering MTA’s production in plants and yeast cells. Although the production of hyoscyamine and scopolamine were obtained in engineered yeast cells, their liter yields are extremely low (Srinivasan and Smolke, 2020). For hyoscyamine and scopolamine, their highest production were, respectively, 80 and 30 μg L−1, very far away from the industry-requiring level, not less than 5 g L−1 (Srinivasan and Smolke, 2020). Therefore, the industrial production of MTAs is, and will be dependent on plant materials, not only at present but in a long future period. In this review, we summarize engineering MTA’s production in planta. The problems and perspectives on engineering MTA’s production are discussed, as well.
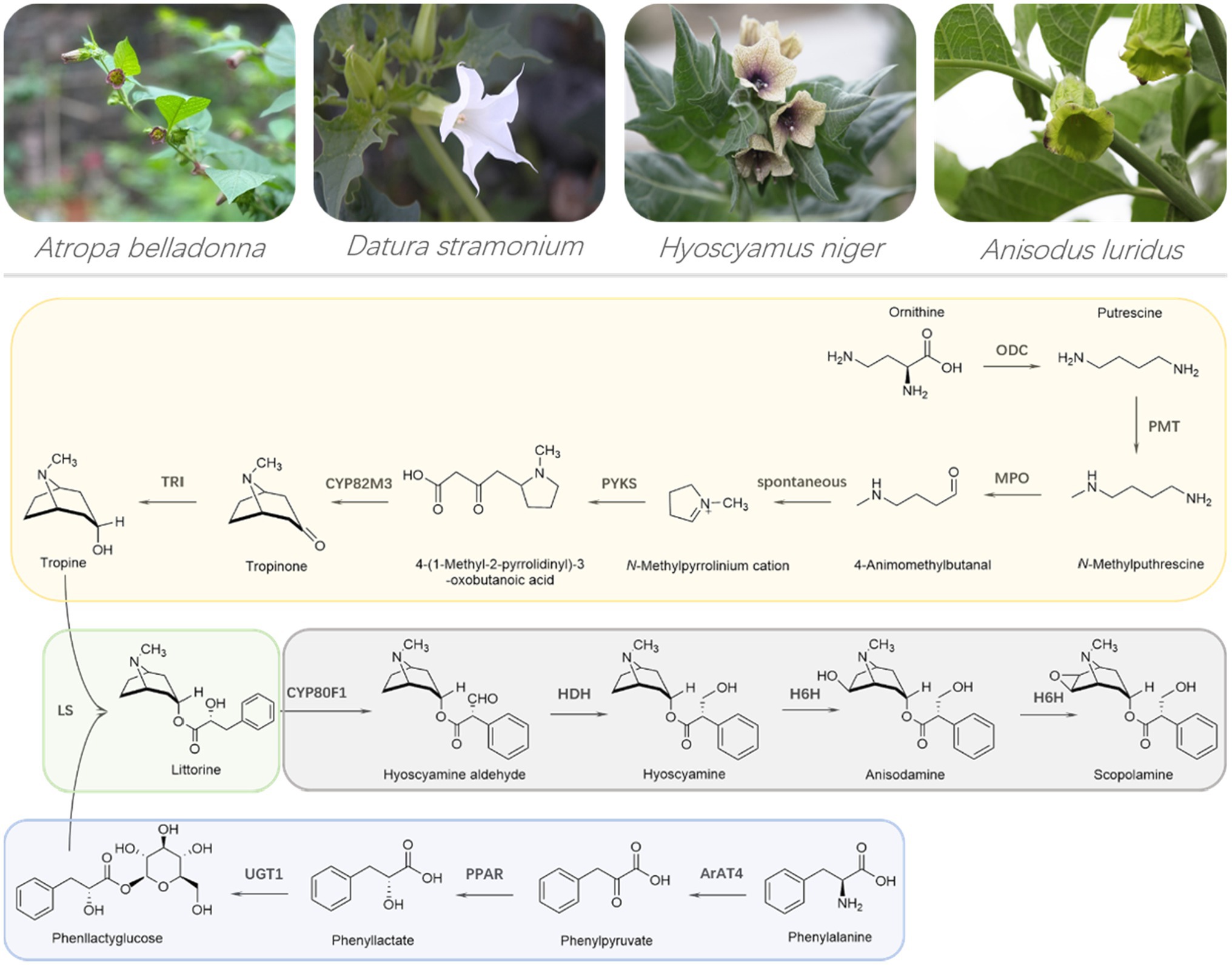
Figure 1. Representative plant species producing medicinal tropane alkaloids and the intact biosynthetic pathway of medicinal tropane alkaloids. ODC, ornithine decarboxylase; PMT, putrescine N-methyltransferase; MPO, N-methylputrescine oxidase; PYKS, polyketide type III synthase; CYP82M3, tropinone synthase; TRI, tropinone reductase I or tropine-forming reductase; ArAT4, phenylalanine aminotransferase; PPAR, phenylpyruvic acid reductase; UGT1, phenyllactate UDP-glycosyltransferase; LS, littorine synthase; CYP80F1, littorine mutase; HDH, hyoscyamine dehydrogenase or hyoscyamine aldehyde reductase; H6H, hyoscyamine 6β-hydroxylase.
Engineering Medicinal Tropane Alkaloid Production in Hairy Roots
Hairy root cultures are often used to engineering production of plant natural products. Hyoscyamine, as well as scopolamine, is specifically synthesized in secondary roots of plants (Li et al., 2006; Matthew et al., 2014; Qiu et al., 2021). A lot of publications report engineering the biosynthesis of MTAs in hairy root cultures of various MTA-producing medicinal plants. Overexpression of biosynthesis genes is a widely applied method to promote metabolite production. Seven biosynthesis genes required for MTA’s biosynthesis are studied in engineering the production of MTAs using hairy root cultures of different plants. Such biosynthesis genes include: ornithine decarboxylase (ODC), putrescine N-methyltransferase (PMT), tropinone reductase I (TRI), littorine mutase (CYP80F1), hyoscyamine dehydrogenase (HDH), and hyoscyamine β-hydroxylase (H6H).
Engineering MTA’s Production by Overexpressing a Single MTA Biosynthesis Gene
Ornithine decarboxylase is the rate-limiting enzyme catalyzing the conversion of ornithine to putrescine (Bacchi et al., 1980). In Atropa belladonna, the ODC enzyme (AbODC) rather than arginine decarboxylase (AbADC) contributes to the MTA’s biosynthesis, and when A. belladonna ODC gene was overexpressed in hairy root cultures and plants of A. belladonna, the production of hyoscyamine and anisodamine was significantly increased (Zhao et al., 2020). Although ODC is located at the beginning of the MTA’s biosynthetic pathway, its role in promoting MTA’s production indicates that it is a key regulator directing ornithine into alkaloid metabolism (Zhao et al., 2020). Overexpression of ODC increases the putrescine level and provides metabolic flux enough for facilitating the downstream biochemical reactions (Zhao et al., 2020). However, such increased metabolic flux is not enough to support that ODC overexpression promotes MTA’s production. Another explanation might be polyamine metabolism regulated by ODC (Nölke et al., 2008). Polyamines are plant growth regulators involved in diverse cell physiology. Since the ODC enzyme is a key primary-metabolism one regulating polyamine biosynthesis, this implies that it may have complicated synergic effects on MTA’s biosynthesis through regulating polyamine biosynthesis.
Putrescine N-methyltransferase is a committed enzyme in catalyzing putrescine to form N-methylputrescine, and this enzyme is a secondary-metabolism enzyme (Hashimoto et al., 1989) Especially, tobacco PMT gene (NtPMT) were overexpressed in hairy root cultures of several MTA-producing plants, and such plants include Duboisia × hybrid (Moyano et al., 2002), Hyoscyamus muticus (Moyano et al., 2003), Datura metal (Moyano et al., 2003), A. belladonna (Rothe et al., 2003), Hyoscyamus niger (Zhang et al., 2004) and Scopolia parviflora (Lee et al., 2005). Overexpression of NtPMT in root cultures of D. hybrid, A. belladonna, and H. niger did not alter the production of hyoscyamine and scopolamine, while overexpression the same PMT gene (NtPMT) in hairy root cultures of S. parviflora, the levels of both hyoscyamine and scopolamine were significantly elevated. Moyano et al analyzed hyoscyamine and scopolamine content in individual clones of H. muticus and D. metal hairy root cultures with NtPMT overexpression, and they concluded that NtPMT overexpression increased hyoscyamine and scopolamine contents in D. metal, but only increased hyoscyamine content in H. muticus. Subsequently, Rothe et al reported their work on overexpression of NtPMT in hairy root cultures of A. belladonna and found no significantly increased production of MTAs when NtPMT was overexpressed. In the publication of Rothe et al. (2003), they discussed that the individual clones indicated both increased and decreased hyoscyamine and scopolamine content when NtPMT was overexpressed in root cultures of H. muticus and Datura metel. Zhang et al also found that overexpression of NtPMT did not promote hyoscyamine and scopolamine content in hairy root cultures of H. niger (Zhang et al., 2004). It should be noted that only two NtPMT-overexpressing root lines of A. belladonna and S. parviflora were used for studying MTA’s production by Rothe et al. (2003) and Lee et al. (2005). Probably, such a few root lines might not be enough to solidify their conclusion. Such different results above suggest the role of PMT in engineering MTA’s production should be carefully studied through intensive experiments.
Tropinone reductase I, also named tropinone-forming reductase, catalyzes tropinone reduction to give tropinone entering formation of littorine, a key intermediate for hyoscyamine biosynthesis (Hashimoto et al., 1992; Dräger, 2006). Richter et al. (2005) reported that overexpression of Datura stramonium TRI gene in hairy root cultures of A. belladonna markedly increased the TRI activity, tropine production, and contents of hyoscyamine and scopolamine. Kai et al found that overexpression Anisodus acutangulus TRI significantly increase the contents of hyoscyamine and scopolamine in hairy root cultures of Ambrysus acutangulus. Zhao et al also found that overexpression of Scopolia lurida (also named Anisodus luridus) TRI gene significantly promoted the production of hyoscyamine and scopolamine in hairy root cultures of S. luridus (Zhao et al., 2017). These previous reports indicate that TRI is a useful target on engineering MTA’s production in hairy root cultures, but there are not reports on TRI overexpression in transgenic plants.
Littorine mutase (CYP80F1) catalyzes littorine to form hyoscyamine aldehyde (Li et al., 2006), which is able to be reduced to give hyoscyamine under catalysis by hyoscyamine dehydrogenase (Qiu et al., 2021). The level of hyoscyamine aldehyde is extremely low in plants, suggesting that CYP80F1 might be a rate-limiting enzyme or hyoscyamine aldehyde might be metabolized very quickly to form some other related alkaloids. However, overexpression of CYP80F1 showed no significant alteration in hyoscyamine and scopolamine production in hairy root cultures of H. niger (Li et al., 2006). Our group identified hyoscyamine dehydrogenase (Qiu et al., 2021), also named hyoscyamine aldehyde reductase (Liao et al., 2019), and its role in engineering MTA’s production was studied. When A. belladonna HDH gene was overexpressed in A. belladonna hairy root cultures, the production of hyoscyamine and scopolamine were markedly elevated, indicating that HDH is a valuable MTA biosynthesis gene in engineering MTA’s production (Qiu et al., 2021).
Hyoscyamine 6β-hydroxylase (H6H) catalyzes hyoscyamine to produce anisodamine through 6β-hydroxylation, and subsequently oxidizes anisodamine to generate scopolamine (Hashimoto and Yamada, 1986) The H6H genes have been identified from H. niger (Hashimoto and Yamada, 1986), A. belladonna (Li et al., 2012), A. acutangulus (Kai et al., 2011a), and S. luridus (Lan et al., 2018). Of these H6H enzymes, the H. niger one (HnH6H) showed the highest efficiency in converting hyoscyamine to scopolamine, and HnH6H is the best choice in engineering scopolamine production. Lan et al compared the effects of HnH6H and S. luridus H6H gene (SlH6H) on engineering scopolamine production, and found that HnH6H was much stronger than SlH6H in promoting scopolamine production (Lan et al., 2018). Without exception, overexpression of H6H definitively promoted the scopolamine content in hairy root cultures of different plants producing MTAs. However, the scopolamine production is quite a lot variable in engineered hairy root cultures generated from different plants, even when the same H6H gene (HnH6H) was overexpressed, indicating that the original capacity on MTA’s production depends on plant species themselves. Impressively, When HnH6H was overexpressed hairy root cultures of H. muticus, the best clone produce scopolamine at a level over 100 times more than the control clones (Jouhikainen et al., 1999). Of these HnH6H-overexpressing hairy root cultures, the highest scopolamine production was reported in H. niger, reaching 184 mg per liter (Zhang et al., 2004). However, it should be noted that hyoscyamine is not able to be completely converted to scopolamine in root cultures, even under HnH6H overexpression. To be concluded, overexpression of H6H, especially HnH6H, is absolutely necessary for engineering scopolamine production.
Engineering MTA’s Production by Overexpressing Combinations of MTA Biosynthesis Genes
From ornithine and phenylalanine to hyoscyamine or scopolamine, more than 10 biosynthesis genes involved. Such a long biosynthetic pathway implies that there are several rate-limiting steps governing by some enzymes. Furthermore, individual MTA’s biosynthesis genes may have synergic effects on regulating alkaloid metabolism. Therefore, overexpression of the combinations of MTA’s biosynthesis genes is very important in engineering MTA’s production. Zhang et al. (2004) reported for the first time that overexpression of both NtPMT and HnH6H led to much higher levels of scopolamine in hairy root cultures than those overexpressing NtPMT or HnH6H. Kai et al. (2011b) also reported that the highest production of MTAs was obtained in A. acutangulus hairy root cultures overexpressing AaPMT and AaTRI together, compared with the scopolamine levels in those overexpressing AaPMT or AaTRI. Overexpression of both AaTRI and AaH6H also gave higher production of MTAs in hairy root cultures of A. acutangulus, than overexpression of AaTRI or AaH6H (Kai et al., 2012). Such overexpression of two MTA’s biosynthesis genes is able to be named a “Pushing-Pulling” strategy of metabolic engineering. When an upstream MTA’s biosynthesis gene, such as PMT before TRI and H6H, is overexpressed, its product as precursors for following enzymes is given more, and this pushes more metabolic flux to the downstream reactions. Simultaneously, when a downstream MTA’s biosynthesis gene is overexpressed, more precursors are able to be converted, showing a pulling force in biosynthetic pathway. To be concluded, overexpression of combination of MTA’s biosynthesis genes usually have better effects on promoting MTA’s production than that of a single gene.
Up to date, there are no reports on engineering MTA’s production through overexpression of three or more biosynthesis genes. Very recently, our group finished evaluation on PYKS, CYP82M3, UGT1, LS, two gene combinations (PYKS + CYP82M3, UGT1 + LS), and four gene combination (PYKS + CYP82M3 + UGT1 + LS), in engineering MTA’s production, and found interesting and valuable discoveries (submitted). According to the previously reported researches and our submitted ones, it can be draw a conclusion on simultaneous overexpression of several biosynthesis genes is a prior strategy in engineering MTA’s production. Although fruitful and marked achievements on engineering MTA’s production in hairy root cultures (Table 1), industrial production has not been successfully realized using hairy root cultures, because their cultivation in a large or industrial scale is not available technologically at present. So, it is necessary to develop engineered plants with high-yield MTA’s production.
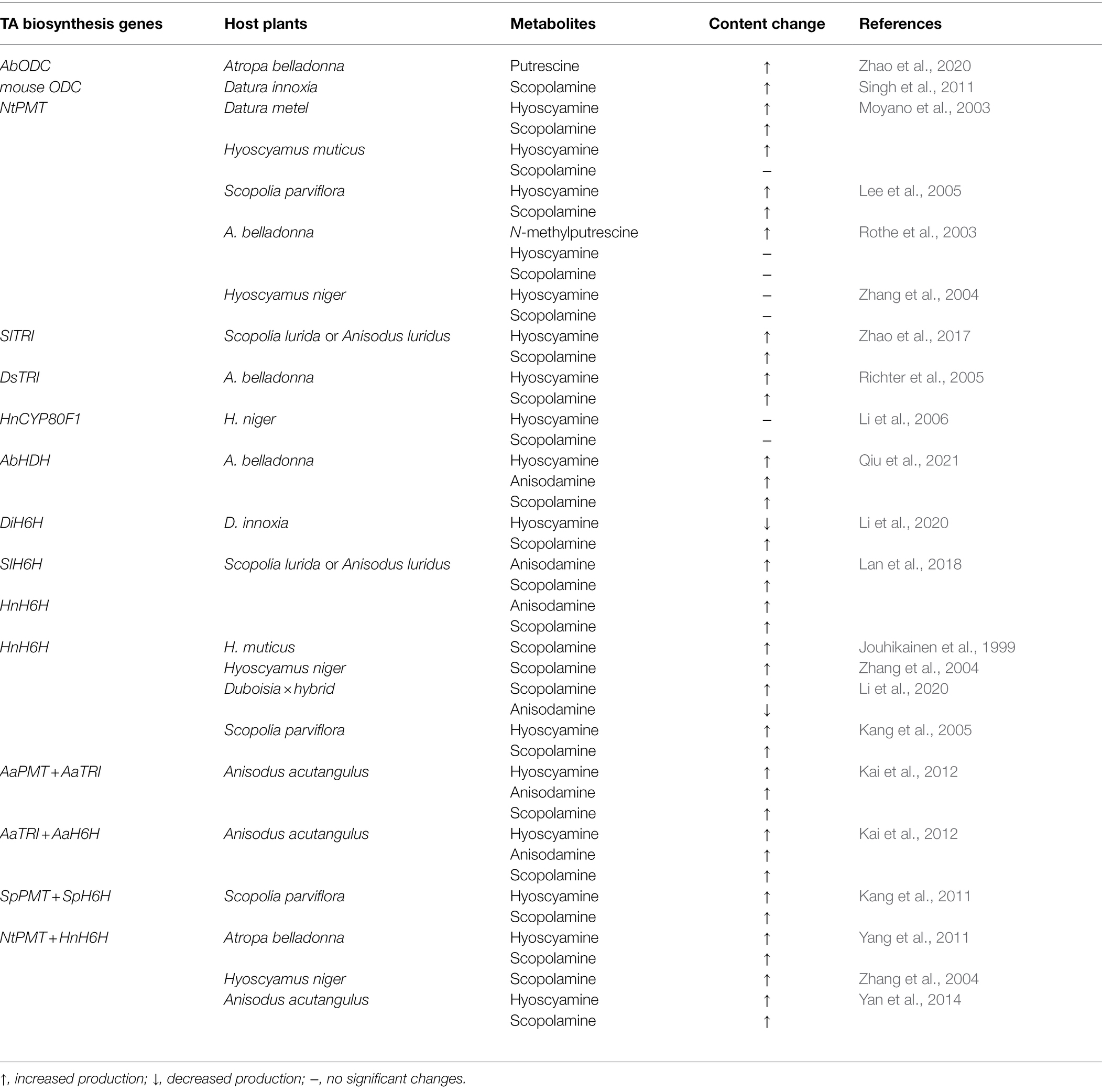
Table 1. Engineering medicinal tropane alkaloid production in plant hairy root cultures through gene overexpression.
Engineering Medicinal Tropane Alkaloid Production in Plants
To date, industrial production of medicinal tropane alkaloids is completely dependent on plant cultivation. So, engineered plants with high yield of hyoscyamine and scopolamine are highly desirable and valuable. For development of such engineered plants, genetic transformation with high efficiency is the most important technology. In fact, generated plants are reported in only a very few plant species producing MTAs, including A. belladonna and Datura innoxia.
Most of engineered plants with high-yield MTAs are reported in A. belladonna with overexpressing HnH6H for enhancement of scopolamine production. In 1992, transgenic plants of A. belladonna with HnH6H overexpression were developed by Yun et al. (1992). They established T0 progeny of HnH6H-overexpressing plants of A. belladonna, and further generated T1 progeny for analyzing the MTA’s production in different organs. The HnH6H-overexpressing plants of A. belladonna were grown under controlled conditions in a growth chamber. Surprisingly, hyoscyamine was almost completely converted to scopolamine in leaves and stems of T1-progeny plants with HnH6H overexpression, whereas only part of hyoscyamine was converted to scopolamine in roots. Such difference indicates that higher efficiency converting hyoscyamine to scopolamine in upground organs than that in underground organs. The scopolamine production was greatly elevated in all tested T1-progeny plants, and the highest scopolamine production was over 1.2% dry weight in leaves of the T1-12 line (Yun et al., 1992). Considering that synergic effects between PMT and H6H, our group generated A. belladonna plants, in which both NtPMT and HnH6H were overexpressed, and the scopolamine production was markedly increased as expected (Wang et al., 2011). Then, we further produced T1 progeny of A. belladonna plants with overexpression of both NtPMT and HnH6H, and investigated MTA’s production under the field conditions in the medicinal plant garden of Tibet Agriculture and Animal Husbandry University (Nyingchi, Tibet, China) and Xinpu Town of Guiyang City (Guizhou Province, China). Similar to the results obtained by Yun et al in HnH6H-overexpssing plants, hyoscyamine was completely converted to scopolamine in aerial organs and high production of scopolamine was detected. In the plants harvested from the field in Xinpu Town of Guiyang City, the highest production of scopolamine was 0.513% DW in leaves, indicating over 12 times higher than in wild-type levels (Xia et al., 2016). Similar results were obtained when A. belladonna plants with overexpressing both NtPMT and HnH6H (T1 progeny) were grown in the field of Tibet Agriculture and Animal Husbandry University, but the highest scopolamine level in leaves nearly reached 1% DW (Quan et al., 2016).
Compared with HnH6H-overexpressing plants generated by Yun et al, the scopolamine production in our transgenic plants was not so high, and possible reasons leading to such difference might have different genotypes of A. belladonna used as initial plant materials for transgenic research, different growth conditions, and some others. Notably, an obvious difference between Yun and our studies is the scopolamine production in roots. The scopolamine levels in secondary roots of HnH6H-overexpressing plants were even much lower than those in wild-type secondary roots, according to Yun et al. (1992). Without exception, overexpression of both NtPMT and HnH6H in greatly increased the scopolamine content of secondary roots in contrast to wild-type plants (Xia et al., 2016). It was postulated that synergic effects overexpressing both NtPMT and HnH6H facilitated the conversion from hyoscyamine to scopolamine in the native organ (secondary roots) synthesizing scopolamine (Zhang et al., 2004; Xia et al., 2016).
The biosynthesis genes including TRI and ODC were also used to engineering MTA’s production in transgenic plants. Zhao et al established transgenic plants of A. belladonna, in which the ODC gene of A. belladonna (AbODC) was overexpressed, and analyzed putrescine, N-methylputrescine, hyoscyamine, anisodamine and scopolamine in roots and leaves (Zhao et al., 2020). The production of putrescine, N-methylputrescine, hyoscyamine and anisodamine were significantly increased in roots of AbODC-overexpressing plants. The levels of putrescine, hyoscyamine, and anisodamine were also elevated in leaves of AbODC-overexpressing plants. The scopolamine production was not significantly altered in roots and leaves, even when AbODC was overexpressed. Overexpression of AbODC increased the production of hyoscyamine anisodamine, but did not promote scopolamine production, indicating that the epoxidation reaction catalyzed by A. belladonna H6H is a key limitation in converting anisodamine to scopolamine (Hashimoto and Yamada, 1986; Kluza et al., 2020). A mouse ODC gene was ectopically expressed in transgenic plants of D. innoxia, the polyamine and scopolamine levels were promoted, but hyoscyamine was not analyzed in this research (Singh et al., 2011). Although the scopolamine production was increased in transgenic plants of D. innoxia, the highest level for scopolamine was extremely low, 0.25 μg g−1 DW (Singh et al., 2011), suggesting that D. innoxia might be not a good candidate for industrially producing scopolamine. Rothe et al developed NtPMT-overexpressing plants of A. belladonna, and found that NtPMT overexpression did not increased the production of scopolamine, hyoscyamine, tropine, pseudotropine, tropinone, and calystegines. It should also be noted that there were only two transgenic plants used for analyzing alkaloid production (Rothe et al., 2003), and this might not be enough to draw a solidified conclusion, because the levels of these metabolites are variable in individual plants.
Notably, all the above transgenic plants with engineering MTA’s biosynthetic pathway are generated using kanamycin as selection pressure, given by neomycin phosphotransferase II (NPTII) as marker gene. As it is well known, the NPTII gene is useless when transgenic plants are obtained and it brings a biosafety risk for commercial usage of engineering plants. When transgenic plants with high-yield MTAs are developed, it is very important to establish transgenic homozygous lines that are highly valuable for agricultural and industrial application. However, no transgenic homozygous plants producing MTAs were reported before 2021. Therefore, our group took a few years to successfully develop transgenic homozygous plants of A. belladonna, with overexpression of both a glyphosate-resistant gene and HnH6H, and without antibiotics-resistant gene (Zhang et al., 2021). After obtaining T0 progeny with one-copy insertion, genetic breeding was used to develop homozygous offsprings. The field trials indicated that homozygous transgenic plants of A. belladonna grew well, resisted the commercial recommended concentration of glyphosate, produced high-yield scopolamine (7.04 mg g−1 DW in leaf). These homozygous transgenic plants of A. belladonna showed much higher commercial value than wild-type ones, because of their glyphosate resistance and high-yield scopolamine.
The CRISPR/Cas9 system is a powerful gene-editing tool, which is also used to generate A. belladonna plants with high-yield hyoscyamine (Zeng et al., 2021). Hyoscyamine is converted to anisodamine, and anisodamine to scopolamine under catalysis of H6H (Hashimoto and Yamada, 1986). Extracts of A. belladonna contain the three above alkaloids with similar structures, and it is expensive to separate each from total alkaloid extracts. Our group generated A. belladonna plants with disruption of the H6H gene through CRISPR/Cas9 system, the H6H-disrupting plants produced hyoscyamine at much higher levels than wild-type ones, because hyoscyamine cannot be converted to anisodamine and scopolamine, and they produced neither anisodamine nor scopolamine. The H6H-disrupting A. belladonna plants are valuable due to high-yield hyoscyamine and convenient separation of hyoscyamine.
Metabolic engineering of MTA’s production usually employs their biosynthesis genes. Very recently, our group has a publication on reporting engineering tropane alkaloid production using a novel calmodulin gene in A. belladonna plants (Zhang et al., 2022). Increased calcium can promote the TA’s production in plants, and accordingly, we postulated that calcium/calmodulin signaling might regulate TA’s biosynthesis and then identified a calcium-binding calmodulin gene (AbCaM1) highly expressed in secondary roots of A. belladonna. Transgenic plants of A. belladonna, in which both AbCaM1 and G2-EPSPS were overexpressed, had strong tolerance to glyphosate and high-yield tropane alkaloids, including hyoscyamine, anisodamine and scopolamine. To our knowledge, this publication is the first report on engineering TA’s production in plants using a regulator protein. Such transgenic homozygous plants of A. belladonna are valuable for industrial production of medicinal TAs, due to their high production of TAs and strong tolerance to herbicide that facilitate manufacturing TAs at low cost.
The problems on Engineering MTA’s Production in Plants
Although the MTA’s biosynthetic pathway has been completely unveiled, using A. belladonna as a model plant producing MTAs. The same enzyme from different MTA-producing plants may have various catalytic efficiency. For example, the H6H enzyme of H. niger shows much higher catalytic efficiency in converting hyoscyamine to scopolamine than the reported others. So, it is necessary to screen MTA’s biosynthesis enzymes with high catalytic efficiency from different plants producing MTAs. According to previous reports on enzymatic assays, it is suggested to pay more attention to the MTA’s biosynthetic enzymes, such as CYP80F1, phenyllactate UDP-glycosyltransferase and littorine synthase, each of which is a key limitation in the MTA’s biosynthesis (Bedewitz et al., 2018; Qiu et al., 2020). Furthermore, over 100 tropane alkaloids are reported in MTA-producing plants, and they compete against the biosynthesis of hyoscyamine and scopolamine. It is largely unknown on these alkaloid biosynthesis, so it is also important to understand their formation, and then genetic operation might be performed in planta to disrupt their biosynthesis and enhance the production of hyoscyamine and scopolamine.
To date, the values of most MTA’s biosynthesis genes are unknown in engineering MTA’s production in plants. It is of importance to have systematic evaluation on the values of MTA’s biosynthesis genes in plant metabolic engineering. And further, their combinations should also be tested in engineering MTA’s production. It can be reasonably predicted that combinations of MTA’s biosynthesis genes will be the main methods used in engineering MTA’s production in the future. Tens of solanaceous plant species are able to produce MTAs, and such abundant resources provide diverse plant chassis for engineering MTA’s production through metabolic engineering. However, of these MTA-producing plants, only the plants of A. belladonna can be efficiently transformed. Lack of genetic transformation at high efficiency is a key technological limitation for modification of MTA’s biosynthetic pathway in plants. So, plant biotechnology, especially transgenic technology, should be strengthened.
The Prospects in Engineering MTA’s Production in Plants
Complete elucidation of MTA’s biosynthetic pathway results in synthesis of hyoscyamine and scopolamine in engineered yeast cells using the approaches of synthetic biology. Professor Smolke built the MTA’s biosynthetic pathway in yeast cells, and successfully synthesized hyoscyamine and scopolamine in engineered yeast cells (Srinivasan and Smolke, 2020). It is a milestone in manufacturing these valuable alkaloids. However, the production of hyoscyamine and scopolamine is extremely low, of which production levels are very far away from the standards of industrial production. Without doubt, the production of MTAs in engineered yeast cells will be gradually promoted, and probably it might reach or even exceed 5 g per liter, the lowest level for industrially producing TAs based on yeast’s fermentation. By the way, synthetic biology’s artemisinin fails in market is a typical case, and the most important reason for this failure is plant-base production of artemisinin is much cheaper than synthetic biology’s.
Surely, the plant biotechnologists have been trying their best to develop novel plant varieties producing MTAs at higher and higher levels, especially through complicated genetic operations composed of overexpression and suppression/gene edition methods. The next-generation strategy on engineering MTA’s production consists of enhancement of MTA’s production by overexpressing MTA’s biosynthesis genes, and simultaneously disruption of competitive metabolite’s production by suppressing or knocking out corresponding biosynthesis genes, in MTA-producing plants. Besides the core target on promoting MTA’s production, horticultural traits, such as biomass especially leaf yield, resistance on stress and pathogens and herbicide resistance, are able to be genetically fortified. Engineered plants, with increased MTA’s production and strengthened horticultural traits, will be the main source for industrial production of hyoscyamine and scopolamine, and engineered yeast cell factory may be an additional source for MTA’s production.
Author Contributions
HG, LZ, and ZL conceived the original idea for the review, and edited and reviewed the manuscript. HG and PH drew the figure. XL and LZ checked the chemical structures and took the pictures of plant species. All authors wrote the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was financially supported by the NSFC projects (U1902212 and 31770335), the Fourth National Survey of Traditional Chinese Medicine Resources, Chinese or Tibet Medicinal Resources Investigation in Tibet Autonomous Region (State Administration of Chinese Traditional Medicine 20191217–540124 and 20200501–542329) and the College Students’ Innovative Entrepreneurial Training Plan Program (202110635108).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Bacchi, C. J., Nathan, H. C., Hutner, S. H., McCann, P. P., and Sjoerdsma, A. (1980). Polyamine metabolism: a potential therapeutic target in trypanosomes. Science 210, 332–334. doi: 10.1126/science.6775372
Bedewitz, M. A., Jones, A. D., D’Auria, J. C., and Barry, C. S. (2018). Tropinone synthesis via an atypical polyketide synthase and P450-mediated cyclization. Nat. Commun. 9, 5281–5213. doi: 10.1038/s41467-018-07671-3
Dräger, B. (2006). Tropinone reductases, enzymes at the branch point of tropane alkaloid metabolism. Phytochemistry 67, 327–337. doi: 10.1016/j.phytochem.2005.12.001
Griffin, J., Lee, S., Caiado, M., Kesten, S., and Price, D. (2008). Comparison of tiotropium bromide and combined ipratropium/salbutamol for the treatment of COPD: a UK general practice research database 12-month follow-up study. Prim. Care Respir. J. 17, 104–110. doi: 10.3132/pcrj.2008.00024
Grynkiewicz, G., and Gadzikowska, M. (2008). Tropane alkaloids as medicinally useful natural products and their synthetic derivatives as new drugs. Pharmacol. Reports 60, 439–463.
Hashimoto, T., Nakajima, K., Ongena, G., and Yamada, Y. (1992). Two tropinone reductases with distinct stereospecificities from cultured roots of Hyoscyamus niger. Plant Physiol. 100, 836–845. doi: 10.1104/pp.100.2.836
Hashimoto, T., and Yamada, Y. (1986). Hyoscyamine 6β-hydroxylase, a 2-Oxoglutarate-dependent Dioxygenase, Alkaloid-Producing Root Cultures. Plant Physiol. 81, 619–625. doi: 10.1104/pp.81.2.619
Hashimoto, T., Yukimune, Y., and Yamada, Y. (1989). Putrescine and putrescine N-methyltransferase in the biosynthesis of tropane alkaloids in cultured roots of Hyoscyamus albus. Planta 178, 131–137. doi: 10.1007/BF00392536
Huang, J. P., Wang, Y. J., Tian, T., Wang, L., Yan, Y., and Huang, S. X. (2021). Tropane alkaloid biosynthesis: a centennial review. Nat. Prod. Rep. 38, 1634–1658. doi: 10.1039/d0np00076k
Jouhikainen, K., Lindgren, L., Jokelainen, T., Hiltunen, R., Teeri, T. H., and Oksman-Caldentey, K. M. (1999). Enhancement of scopolamine production in Hyoscyamus muticus L. hairy root cultures by genetic engineering. Planta 208, 545–551. doi: 10.1007/s004250050592
Kai, G., Liu, Y., Wang, X., Yang, S., Fu, X., Luo, X., et al. (2011a). Functional identification of hyoscyamine 6β-hydroxylase from Anisodus acutangulus and overproduction of scopolamine in genetically-engineered Escherichia coli. Biotechnol. Lett. 33, 1361–1365. doi: 10.1007/s10529-011-0575-y
Kai, G., Yang, S., Luo, X., Zhou, W., Fu, X., Zhang, A., et al. (2011b). Co-expression of AaPMT and AaTRI effectively enhances the yields of tropane alkaloids in Anisodus acutangulus hairy roots. BMC Biotechnol. 11. doi: 10.1186/1472-6750-11-43
Kai, G., Zhang, A., Guo, Y., Li, L., Cui, L., Luo, X., et al. (2012). Enhancing the production of tropane alkaloids in transgenic Anisodus acutangulus hairy root cultures by over-expressing tropinone reductase i and hyoscyamine-6β-hydroxylase. Mol. BioSyst. 8, 2883–2890. doi: 10.1039/c2mb25208b
Kang, Y. M., Lee, O. S., Jung, H. Y., Kang, S. M., Lee, B. H., Karigar, C., et al. (2005). Overexpression of hyoscyamine 6β-hydroxylase(h6h) gene and enhanced production of tropane alkaloids in Scopolia parviflora hairy root lines. J. Microbiol. Biotechnol. 15, 91–98.
Kang, Y. M., Park, D. J., Min, J. Y., Song, H. J., Jeong, M. J., Kim, Y. D., et al. (2011). Enhanced production of tropane alkaloids in transgenic Scopolia parviflora hairy root cultures over-expressing putrescine N-methyl transferase (PMT) and hyoscyamine-6β-hydroxylase (H6H). Vitr. Cell. Dev. Biol. Plant 47, 516–524. doi: 10.1007/s11627-011-9367-2
Kluza, A., Wojdyla, Z., Mrugala, B., Kurpiewska, K., Porebski, P. J., Niedzialkowska, E., et al. (2020). Regioselectivity of hyoscyamine 6β-hydroxylase-catalysed hydroxylation as revealed by high-resolution structural information and QM/MM calculations. Dalt. Trans. 49, 4454–4469. doi: 10.1039/d0dt00302f
Lan, X., Zeng, J., Liu, K., Zhang, F., Bai, G., Chen, M., et al. (2018). Comparison of two hyoscyamine 6β-hydroxylases in engineering scopolamine biosynthesis in root cultures of Scopolia lurida. Biochem. Biophys. Res. Commun. 497, 25–31. doi: 10.1016/j.bbrc.2018.01.173
Lee, O. S., Kang, Y. M., Jung, H. Y., Min, J. Y., Kang, S. M., Karigar, C. S., et al. (2005). Enhanced production of tropane alkaloids in Scopolia parviflora by introducing the PMT (putrescine N-methyltransferase) gene. Vitr. Cell. Dev. Biol. Plant 41, 167–172. doi: 10.1079/IVP2004621
Li, R., Reed, D. W., Liu, E., Nowak, J., Pelcher, L. E., Page, J. E., et al. (2006). Functional genomic analysis of alkaloid biosynthesis in Hyoscyamus niger reveals a cytochrome P450 involved in Littorine rearrangement. Chem. Biol. 13, 513–520. doi: 10.1016/j.chembiol.2006.03.005
Li, J., Van Belkum, M. J., and Vederas, J. C. (2012). Functional characterization of recombinant hyoscyamine 6β-hydroxylase from Atropa belladonna. Bioorganic Med. Chem. 20, 4356–4363. doi: 10.1016/j.bmc.2012.05.042
Li, Q., Zhu, T., Zhang, R., Bu, Q., Yin, J., Zhang, L., et al. (2020). Molecular cloning and functional analysis of hyoscyamine 6β-hydroxylase (H6H) in the poisonous and medicinal plant Datura innoxia mill. Plant Physiol. Biochem. 153, 11–19. doi: 10.1016/j.plaphy.2020.04.021
Liao, Z., Chen, M., Yang, C., Qiu, F., and Zeng, J. (2019). 莨菪醛还原酶及其应用. The patent number “ZL 2019 1 0826205.8”.
Matthew, A. B., Gonzales-Vigil, E., Góngora-Castillo, E., Uebler, J. B., Gonzales-Vigil, E., Wiegert-Rininger, K. E., et al. (2014). A root-expressed l-phenylalanine:4-hydroxyphenylpyruvate aminotransferase is required for tropane alkaloid biosynthesis in Atropa belladonna. Plant Cell 26, 3745–3762. doi: 10.1105/tpc.114.130534
Moyano, E., Fornalé, S., Palazón, J., Cusidó, R. M., Bagni, N., and Piñol, M. T. (2002). Alkaloid production in Duboisia hybrid hairy root cultures overexpressing the pmt gene. Phytochemistry 59, 697–702. doi: 10.1016/S0031-9422(02)00044-4
Moyano, E., Jouhikainen, K., Tammela, P., Palazón, J., Cusidó, R. M., Piñol, M. T., et al. (2003). Effect of pmt gene overexpression on tropane alkaloid production in transformed root cultures of Datura metel and Hyoscyamus muticus. J. Exp. Bot. 54, 203–211. doi: 10.1093/jxb/erg014
Nölke, G., Schneider, B., Agdour, S., Drossard, J., Fischer, R., and Schillberg, S. (2008). Modulation of polyamine biosynthesis in transformed tobacco plants by targeting ornithine decarboxylase to an atypical subcellular compartment. Open Biotechnol. J. 2, 183–189. doi: 10.2174/1874070700802010183
Qiu, F., Yan, Y., Zeng, J., Huang, J. P., Zeng, L., Zhong, W., et al. (2021). Biochemical and metabolic insights into Hyoscyamine dehydrogenase. ACS Catal. 11, 2912–2924. doi: 10.1021/acscatal.0c04667
Qiu, F., Zeng, J., Wang, J., Huang, J. P., Zhou, W., Yang, C., et al. (2020). Functional genomics analysis reveals two novel genes required for littorine biosynthesis. New Phytol. 225, 1906–1914. doi: 10.1111/nph.16317
Quan, H., Xia, K., Zeng, J., Chen, M., Lan, X., and Liao, Z. (2016). Overexpression of NtPMT and HnH6H changed hyoscyamine-rich Atropa belladonna to scopolamine-rich varieties. Acta Pharm. Sin. 51, 1913–1919. doi: 10.16438/j.0513-4870.2016-0275
Richter, U., Rothe, G., Fabian, A. K., Rahfeld, B., and Dräger, B. (2005). Overexpression of tropinone reductases alters alkaloid composition in Atropa belladonna root cultures. J. Exp. Bot. 56, 645–652. doi: 10.1093/jxb/eri067
Rothe, G., Hachiya, A., Yamada, Y., Hashimoto, T., and Dräger, B. (2003). Alkaloids in plants and root cultures of Atropa belladonna overexpressing putrescine N-methyltransferase. J. Exp. Bot. 54, 2065–2070. doi: 10.1093/jxb/erg227
Singh, A., Nirala, N. K., Das, S., Narula, A., Rajam, M. V., and Srivastava, P. S. (2011). Overexpression of odc (ornithine decarboxylase) in Datura innoxia enhances the yield of scopolamine. Acta Physiol. Plant. 33, 2453–2459. doi: 10.1007/s11738-011-0787-8
Srinivasan, P., and Smolke, C. D. (2020). Biosynthesis of medicinal tropane alkaloids in yeast. Nature 585, 614–619. doi: 10.1038/s41586-020-2650-9
Wang, X., Chen, M., Yang, C., Liu, X., Zhang, L., Lan, X., et al. (2011). Enhancing the scopolamine production in transgenic plants of Atropa belladonna by overexpressing pmt and h6h genes. Physiol. Plant. 143, 309–315. doi: 10.1111/j.1399-3054.2011.01506.x
Xia, K., Liu, X., Zhang, Q., Qiang, W., Guo, J., Lan, X., et al. (2016). Promoting scopolamine biosynthesis in transgenic Atropa belladonna plants with pmt and h6h overexpression under field conditions. Plant Physiol. Biochem. 106, 46–53. doi: 10.1016/j.plaphy.2016.04.034
Yan, Z., Pan, X., Qiang, W., and Liao, Z. (2014). Improvement of biosynthesis of Tropane alkaloids in Anisodus acutangulus by co-transformed PMT and H6H. J. Southwest Univ. 36:7
Yang, C., Chen, M., Zeng, L., Zhang, L., Liu, X., Lan, X., et al. (2011). Improvement of tropane alkaloids production in hairy root cultures of Atropa belladonna by overexpressing pmt and h6h genes. Plant Omics 4, 29–33.
Yun, D. J., Hashimoto, T., and Yamada, Y. (1992). Metabolic engineering of medicinal plants: transgenic Atropa belladonna with an improved alkaloid composition. Proc. Natl. Acad. Sci. U. S. A. 89, 11799–11803. doi: 10.1073/pnas.89.24.11799
Zeng, L., Zhang, Q., Jiang, C., Zheng, Y., Zuo, Y., Qin, J., et al. (2021). Development of Atropa belladonna L. plants with high-yield hyoscyamine and without its derivatives using the CRISPR/Cas9 system. Int. J. Mol. Sci. 22, 1–10. doi: 10.3390/ijms22041731
Zhang, L., Ding, R., Chai, Y., Bonfill, M., Moyano, E., Oksman-Caldentey, K. M., et al. (2004). Engineering tropane biosynthetic pathway in Hyoscyamus niger hairy root cultures. Proc. Natl. Acad. Sci. U. S. A. 101, 6786–6791. doi: 10.1073/pnas.0401391101
Zhang, Q., Liang, M., Liu, Y., Yang, C., Zeng, J., Qin, J., et al. (2021). Development of homozygous transgenic Atropa belladonna plants with glyphosate resistance and high-yield scopolamine using metabolic engineering. Ind. Crop. Prod. 171:113953. doi: 10.1016/j.indcrop.2021.113953
Zhang, Q., Liang, M., Zeng, J., Yang, C., Qin, J., Qiang, W., et al. (2022). Engineering tropane alkaloid production and glyphosate resistance by overexpressing AbCaM1 and G2-EPSPS in Atropa belladonna. Metab. Eng. 72, 237–246. doi: 10.1016/j.ymben.2022.03.014
Zhao, T., Li, S., Wang, J., Zhou, Q., Yang, C., Bai, F., et al. (2020). Engineering tropane alkaloid production based on metabolic characterization of ornithine decarboxylase in Atropa belladonna. ACS Synth. Biol. 9, 437–448. doi: 10.1021/acssynbio.9b00461
Keywords: biosynthesis, biotechnology, metabolic engineering, synthetic biology, tropane alkaloids
Citation: Gong H, He P, Lan X, Zeng L and Liao Z (2022) Biotechnological Approaches on Engineering Medicinal Tropane Alkaloid Production in Plants. Front. Plant Sci. 13:924413. doi: 10.3389/fpls.2022.924413
Edited by:
Yongliang Liu, University of Kentucky, United StatesReviewed by:
Zheyong Xue, Northeast Forestry University, ChinaYuan Yuan, China Academy of Chinese Medical Sciences, China
Copyright © 2022 Gong, He, Lan, Zeng and Liao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zhihua Liao, zhliao@swu.edu.cn; Lingjiang Zeng, zengling@swu.edu.cn