
Abstract
This study established a liquid adventitious root culture for Inula crithmoides L. focusing on producing extracts rich in hepatoprotective caffeoylquinic acids through elicitation. Adventitious roots were induced from in vitro-grown plant leaves cultured on MS medium supplemented with 1 mg/L IBA. Growth and secondary metabolites accumulation (phenolics, flavonoids, hydroxycinnamic acids) were monitored weekly to determine the optimal harvesting time. Then, the adventitious roots were elicited for 4 weeks with varying concentrations of yeast extract–YE (50, 100 and 200 mg/L) and extracted with ethanol. The extracts were evaluated for their hepatoprotective effects against ethanol-induced hepatotoxicity and analyzed through high-performance liquid chromatography coupled with electrospray ionization mass spectrometry (HPLC–ESI–MS/MS). Roots of greenhouse-cultivated plants were used to compare the results obtained from in vitro -grown adventitious roots. IBA supplementation induced 100% adventitious root formation, with maximum biomass and phenolic accumulation after 4 weeks. Hydroxycinnamic acid levels increased significantly with YE treatment at all the concentrations, particularly in case of chlorogenic acid and di-O-caffeoylquinic acid isomers. The root extracts displayed significant in vitro hepatoprotective effects, particularly at 50 mg/L YE elicitation. Overall, the elicitation of I. crithmoides liquid root cultures with yeast extract offers a promising, efficient, and cost-effective approach for optimizing the production of valuable caffeoylquinic acids having potential pharmaceutical use.
Key message
Successful establishment of golden samphire adventitious root cultures demonstrates enhanced hepatoprotective caffeoylquinic acid production through yeast extract elicitation.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Inula chrithmoides L. (syn. Limbarda crithmoides L. Dumort), commonly known as golden samphire, is a flowering plant belonging to the Asteraceae family. It is a self-fertile perennial dicotyledonous succulent plant that can grow up to 1 m tall. Its flowering season is from July to August, and the seeds ripen from August to September. It is distributed along the Mediterranean coast, Britain and Western Asian, inhabiting saltmarshes, and maritime cliffs and rocks (Clapham et al. 1962, eHALOPH 2023). Moreover, this is an edible and aromatic halophyte species, where its young leaves or shoots can be eaten raw, cooked, or pickled (Zurayk and Baalbaki 1996; D’Agostino et al. 2022), and it holds ethnomedicinal uses such as treating rheumatic pain and goiter (eHALOPH 2023; Miara et al. 2021). This versatile plant can therefore be used for different purposes, not only as food but also as a source of natural products with health benefits. The cultivation of golden samphire has been optimized in different trials, either using soilless systems under different salinity conditions (Lima et al. 2021) or in neglected salt pans, irrigated using natural estuarine water (Duarte et al. 2022). In Portugal, golden samphire is commercially produced by RiaFresh in a soilless system and sold as a gourmet vegetable.
Many Inula species, including I. crithmoides, endow important therapeutic effects, including antimicrobial (Deriu et al. 2008), anti-inflammatory (Hernández et al. 2007), antioxidant (Kogure et al. 2004), and antihepatotoxic activities (Saygi et al. 2003), related to pharmacologically active compounds such as polyphenols, including many quinic acid derivatives, abundantly present in these plants. The roots of the golden samphire are particularly rich in these molecules, including different isomers of caffeoylquinic acids (Ela et al. 2012), which have been described with potent pharmacological properties, including antioxidant, antibacterial, antiparasitic, anti-inflammatory, anticancer, antiviral, antidiabetic, along with neuro-, hepato- and cardioprotective effects (Liu et al. 2020). Moreover, 5-caffeoylquinic acid (chlorogenic acid) has been recently approved by the China Food and Drug Administration (CFDA) as an anticancer drug for phase I (NCT02728349) and phase II (NCT03758014) clinical trials in glioma patients (Nwafor et al. 2022). Furthermore, the data provided by companies to the European Chemicals Agency (ECHA) under the Classification Labelling and Packaging (CLP) system reveals that 3,5-di-O-caffeoyl quinic acid (also known as Isochlorogenic acid A) has not been classified for any hazards so far (ECHA 2024).
Plant tissue culture is a promising biotechnological tool for plants having potential commercial uses in pharma, cosmetic, or food industries. Specifically, root culture is an effective technique to produce important secondary metabolites that accumulates naturally in such organ, including alkaloids, glycosides, phenolics, flavonoids or terpenoids, allowing for the year-round production of biomass with reduced cost and time. Given the rising demands in the industrial sector for bioactive secondary metabolites, the paramount significance of optimizing metabolite production becomes evident for the pharmaceutical industry (Khanam et al. 2022). The in vitro production of secondary metabolite often includes the use of elicitation techniques to improve the yield (Ramirez-Estrada et al. 2016). Also, the cells or organs are maintained in an optimal medium for biomass growth and then transferred to an optimal production medium for stimulating the synthesis of target metabolites (Ramirez-Estrada et al. 2016; Wawrosch and Zotchev 2021). Such elicitation can be achieved by using different stress factors, including biotic (e.g., proteins, fungus, rhizobacteria, hormones) and/or abiotic (e.g., drought, salinity, light, temperature) (Halder et al. 2019, Alcalde et al. 2022).One common elicitor is the yeast extract (YE), comprising a group of compounds known for their positive impact on plant growth, productivity, microelement composition, and the levels of phytohormones and other plant metabolites (Halder et al. 2019). For instance, its effects have significantly increased the accumulation of phenolic compounds in callus cultures from Annurca apple (Laezza et al. 2024), improved the flavonoid accumulation of adventitious roots from Oplopanax elatus (Jin et al. 2023), and have increased the shoot biomass and lupeol production in Hemidesmus indicus (L.) R. Br. ex. Schult. and Tylophora indica (Burm. F.) Merrill (Pathak et al. 2023). However, as per reviewed literature, there is no report on the in vitro culture of adventitious roots of halophyte species. Thus, this work aimed to establish adventitious roots from leaves of in vitro cultures of Inula crithmoides L., targeting the production of caffeoylquinic acids.
Materials and methods
Establishment of adventitious roots culture
Golden samphire plants were grown in vitro according to the micropropagation protocol detailed by Rodrigues et al. (2023). Then, the leaves (approx. 1.8 cm × 0.25 cm) from in vitro propagated golden samphire plants were excised and transferred to Murashige and Skoog medium (MS) (Murashige and Skoog 1962) containing 30 g/L sucrose, 1% agar, supplemented with 1 mg/L indole-3-butyric acid (IBA). Inoculated leaves sections were then incubated in the dark at 25 ± 1 °C, for 4 weeks. Each treatment was repeated three times, with 10 explants per replicate. After this period, the effect of IBA supplementation on initiation of adventitious roots from explants was assessed by calculating the percentage of rooting. The adventitious roots excised from the leaf explants (0.3 g) were inoculated in Erlenmeyer flasks (100 mL) containing 50 mL of liquid MS media supplemented with the same auxin combination (1 mg/L of IBA) and then cultured on a gyratory shaker at 120 rpm. The adventitious roots were sub-cultured every 4 weeks.
Determination of growth curve for liquid adventitious root culture
From an adventitious root culture with 4-week-old, moisture was removed by drying the roots in sterilized filter paper. Different ratios of biomass were weighted (0.3, 0.5, 0.7, 0.9, 1.2, and 2.1 g) of adventitious roots (in triplicate) and transferred to a 100 mL Erlenmeyer flask containing 50 mL of MS medium supplemented with 1 mg/L of IBA. Once the optimal growth ratio was identified (0.3 g per 50 mL), a growth curve was established by weighing the biomass to determine fresh weight, every week for 5 weeks. The growth index was also calculated by the following formula: (final weight – initial weight) / initial weight. To determine the best time of collection for higher accumulation of secondary metabolites, the total contents of different phenolic groups was estimated for each week (see Sect. "Estimation of phenolic groups content").
Biotic elicitation of liquid adventitious root culture
Four-week-old adventitious root culture was used to establish the biotic elicitation assay. Excess moisture was removed by drying the roots in sterilized filter paper, and 0.3 g of adventitious roots (in triplicate) and transferred to a 100 mL Erlenmeyer flask containing 50 mL of MS medium supplemented with 1 mg/L of IBA and different concentrations of yeast extract (0, 50, 100 and 200 mg/L). After 4 weeks of culture, the experiment was stopped, the roots washed and excess moisture removed, and then freeze dried.
Preparation of the extracts
Freeze dried samples were ground to powder with a pestle and mortar, and then extracted with 96% ethanol (1:40; w/v) in an ultrasonic water bath. The extracts were filtered with filter paper (Whatman No. 4) and the solvent removed in a rotary evaporator. Then, an extracts’ stock solution of 10 mg/mL was prepared in 96% ethanol and stored at -20 °C until further analysis.
Estimation of phenolic groups content
The total contents of different groups of phenolic compounds were determined using 96-well plates, as reported before (Rodrigues et al. 2015). The total phenolic content (TPC) was determined by the method of Folin-Ciocalteu according to Velioglu et al. (1998), total flavonoids (TFC) by the aluminium chloride (AlCl3) colorimetric technique (Zou et al. 2011)., and the total hydroxycinnamic acids using the HCl-ethanol assay as described in Mazza et al. (1999) and adapted to 96-well microplates (Rodrigues et al. 2015). Results were respectively expressed as gallic acid (GA), quercetin (Q) and caffeic acid (CA) equivalents (E) in milligrams per gram (mg/g) of dried extract (dry weight, DW), using a calibration curve with standard solutions at concentrations ranging from 0.002 to 2 mg/mL.
Ultrahigh-Performance Liquid Chromatography Coupled with Electrospray Ionization Mass/Mass Spectrometry (UHPLC-Esi–MS/MS) analyses
The chemical composition of the extracts was determined using a Dionex Ultimate 3000RS (Thermo Scientific) UHPLC equipment. All extracts were filtered before injection using 0.22 μm PTFE syringe filters. The compounds were separated on a Thermo Accucore C18 column (100 mm × 2.1 mm i. d. 2.6 μm) thermostated at 25 °C (± 1 °C). Water (A) and methanol (B) (both were acidified with 0.1% formic acid) were used as eluents. The flow rate was maintained at 200μL/min. The gradient elution performed as follows: 0–3 min. 95% of mobile phase A; 3–43 min. → 0% A; 43–61 min. 0% A; 61–62 min. → 95% A; 62–70 min. 95% A. The UHPLC was connected to a Thermo Q Exactive Orbitrap mass spectrometer equipped with an electrospray ionization source (Thermo Scientific). MS spectra were obtained in both positive and negative ion modes. Settings with defined resolution, i.e., resolution 70000; collision energy: 30 NCE; scan range: 100 to 1500 m/z. Trace Finder 3.1 (Thermo Scientific) software was used to analyse the raw files. The secondary metabolites were identified based on our previous published works and own online databases (Massbank of North America, m/z Cloud etc.). In every case, the exact molecular mass, isotopic pattern, characteristic fragment ions and retention time were used for the identification of the metabolites. The difference between the measured and calculated monoisotopic molecular masses was less than 5 ppm in every case.
For the quantitative determinations, the standard mix in six different concentrations were injected and the metabolite concentrations were calculated based on the calibration curves. For the two unidentified compounds, di-O-caffeoylquinic acid isomers and 3,5-di-O-caffeoylquinic acid were used as standard because they are structural isomers, and their degree of ionization is similar.
Evaluation of the in vitro hepatoprotective properties
Cell culture
The HepG2 (human hepatocellular carcinoma) cell line was maintained in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% heat-inactivated Fetal Bovine Serum (FBS), 1% L-glutamine (2 mM), and 1% penicillin (50 U/mL)/streptomycin (50 µg/mL) and were maintained at 37 °C in humidified atmosphere with 5% CO2.
Cellular viability assay
HepG2 cells were plated in 96-well tissue plates at a density of 5 × 103 cells/well and were incubated for 24 h. Then, the extracts were applied at several concentrations (3.125, 6.25, 12.5, 25, 50 and 100 µg/mL) for 24 h. The 3-(4,5-Dimethylthiazol-2-yl)-2,5 Diphenyltetrazolium bromide (MTT) colorimetric test was used to determine the cellular viability (Biochrom EZ Read 400), as formerly detailed (Rodrigues et al. 2021). The absorbance was measured at 590 nm, and the results were expressed in terms of cellular viability (%) in relation to a control containing ethanol at the same concentration of the samples (0.5%).
Hepatoprotective assay
The non-toxic concentrations (> 80% of cellular viability) of the extracts were evaluated for their capacity to protect hepatic cells from ethanol-induced cytotoxicity. HepG2 cells were plated in 96-well tissue plates at the same density of the cellular viability assay. After an incubation of 24 h, the samples were applied at the non-toxic concentrations (3.125, 6.25, 12.5, 25, 50 and 100 µg/mL) for 24 h. The extracts were removed and 500 mM ethanol solution (allowing for a reduction of 50% of cell viability) was added and incubated for more 24 h. Afterwards, the cellular viability was measured as described in Sect. "Cellular viability assay".
Statistical analyses
The results were expressed as the mean ± standard error (SE), and the experiments were conducted at least in triplicate. Significant differences were assessed by ANOVA and Dunnett's tests. p values lower than 0.05 were considered significant. All the statistical analyses were done using the XLSTAT statistical platform for Microsoft Excel (version 2013, Microsoft Corporation).
Results and discussion
Establishment of adventitious roots cultures: growth curve and phenolics accumulation
Plant roots contain highly bioactive compounds crucial for functions like nutrient absorption, defense, stress response, storage, signaling, and environmental interactions, with potential applications in pharmaceuticals, agriculture, and natural products (Isah 2019; Oladipo et al. 2022). Actually around 60% of traditional herbal preparations contain plant roots as main ingredients in their formulations, which highlights the importance of such plant organ as a source of health promoting ingredients (Hussain et al. 2022).
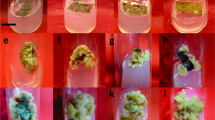
Furthermore, the synthesis and accrual of commercially valuable bioactive secondary metabolites, including phenols, terpenoids, and alkaloids, via adventitious root culture, represents a promising avenue for the eco-friendly generation of pharmaceutical-grade natural products. This is primarily due to their heightened growth rate and consistent metabolite output (Hussain et al. 2022). In this sense, adventitious root cultures of golden samphire were established from leaf explants from in vitro cultured plantlets (Rodrigues et al. 2023), in solid media supplemented with 1 mg/L IBA (Fig. 1A), achieving 100% of adventitious roots formation after 4 weeks of inoculation, being then transferred to liquid media and further cultivated for 5 weeks (Fig. 1B). Biomass production was monitored, along with the total levels of phenolics, flavonoids and hydroxycinnamic acids (Fig. 2). The maximum root biomass accumulation (17.2 g per 50 mL) was observed after 4 weeks of growth (Fig. 2A), showing a growth index of 62.3. Similarly, the maximum content in phenolics (31.31 mg GAE/g DE), flavonoids (29.49 mg QE/g DW), and hydroxycinnamic acids (75.34 mg CAE/g DW) was also observed at the 4th week of culture, remaining significantly similar up to the 5th week (Fig. 2B). Nevertheless, the levels of phenolics, flavonoids, and hydroxycinnamic acids showed a decline in week 2, rebounding in week 3 to levels comparable to those observed in week 1 (Fig. 2B). The roots morphology and color remained the same throughout the experiment (data not shown). Thus, the best collection time for increased biomass and phenolics accumulation was defined as the 4th week of culture.

Four-week adventitious roots in solid MS media containing 1 mg/L of IBA (A), after 1 (B) and 4 weeks (C) of being transferred to the same liquid media

Growth curve (A) and phenolics accumulation (B) per week of culture (W1: week 1; W2: week 2; W3: week 3; W4: week 4; and W5: week 5). Values correspond to mean ± SEM of six independent experiments (n = 6). For each group, columns marked with different letters (a–c) are considered statistically different at p < 0 05 (Tukey’s HSD)
Week 1 represents the end of lag phase, where cells are still metabolically active but not dividing and the growth rate is slow, where cells still synthesize some secondary metabolites in response to stress factors. In turn, the decreased in secondary metabolites observed in week 2, may be related to the degradation of these molecules favoring the synthesis of primary metabolites supporting the optimal growth and development of roots during the exponential growth phase. In contrast, secondary metabolites are increasingly produced during the end or near the stationary phase of growth due to increased stress conditions, such as nutrient depletion (Salam et al. 2023).
Data on adventitious root cultures in golden samphire is lacking; nevertheless, studies on various Asteraceae family species, including Chrysanthemum indicum Linné (Ghimire et al. 2022), Acmella radicans (Jacq.) R.K. Jansen (Bernabé-Antonio et al. 2023), and the halophyte Cineraria maritima Linn. (Banerjee et al. 2004), have reported the addition of IBA to induce adventitious root growth. Moreover, the use of IBA have also been described for other halophytes species as Corema album (L.) D.Don (Alves et al. 2021), Urginea maritima (L.) Baker (Aasim et al. 2008) or Limonium bicolor (Bunge) Kuntze (Yuan et al. 2014). Indeed, IBA functions as an auxin and plays a central role in promoting the formation of adventitious roots (Bai et al. 2020). It is known for its stability and effectiveness over IAA in its ability to induce adventitious roots formation in explants cultured in vitro. Consequently, it is extensively utilized for clonal propagation (Bellini et al. 2014).
Regarding the accumulation of phenolic compounds, it showed an analogous and dependent trend on the accumulation of biomass, where the highest growth and phenolic content was reached from the 4th week onwards. This tendency was also observed for other liquid adventitious root cultures, as for Polygonum multiflorum L. (Ho et al. 2018), or Castilleja tenuiflora Benth. (Gómez-Aguirre et al. 2012), where the highest accumulation of phenolics was achieved by the end of logarithmic and beginning of stationary phases. As mentioned above, this phase involves a slowing down of growth and a shift towards the synthesis of secondary metabolites, noted by the increased phenolic content, typically produced in response to increased stress conditions (Salam et al. 2023). The findings emphasize the importance of growth phase dynamics in influencing the synthesis of secondary metabolites, shedding light on potential strategies for optimizing the production of bioactive compounds in golden samphire. Overall, the establishment of golden samphire adventitious root cultures was achieved with success, demonstrating the ability to gather substantial amounts of biomass and phenolic compounds (primarily hydroxycinnamic acids) within a span of 4 weeks.
Effects of elicitation on adventitious growth and phenolics accumulation
Supplying highly demanding markets can be challenging due to the low production yield of specific metabolites, which can only be produced during specific developmental stages, or in response to particular environmental conditions (Atanasov et al. 2015). But, plant tissue culture tools have been optimized to allow the control of metabolite synthesis under ideal conditions. Elicitation techniques have thus emerged as a highly effective strategy in enhancing the biotechnological synthesis of valuable compounds (Ramirez-Estrada et al. 2016; Wawrosch and Zotchev 2021). However, the selection of proper elicitors and dosage need to be optimized to enhance the product yield (Wawrosch and Zotchev 2021). In this context, adventitious root cultures of golden samphire were elicited by applying yeast extract at three concentrations, followed by the assessment of its effects on biomass yield (Fig. 3A), and on the accumulation of phenolic compounds. The total phenolics levels of ethanolic root extract of greenhouse-grown plants, which were used to establish the in vitro cultures, were also determined, and used for comparison (Fig. 3B).

Biomass (A) and phenolics accumulation (B) of the adventitious roots of I. crithmoides grown into control media (Ctrl; MS + 1 mg/L IBA), elicited with yeast extract (YE) at three different concentrations (50 mg/L,100 mg/L and 200 mg/L), and roots from greenhouse-grown plants. Values correspond to mean ± SEM of six independent experiments (n = 6). For each group, the columns marked with different letters (a–b) are considered statistically different at p < 0 05 (Tukey’s HSD)
The application of the yeast extract in liquid adventitious root cultures did not significantly affect the biomass yield up to the concentration of 100 mg/L (100 – 87%; P < 0.05). However, at 200 mg/L a reduction in the root biomass production was observed, down to 60%, when compared to the control conditions (Fig. 3A). A similar tendency was observed for the accumulation of total phenolics and flavonols content, i.e., adventitious roots grown in media supplemented with yeast extract induced an increase in their levels up to the concentration of 100 mg/L, followed by a decrease at the concentration of 200 mg/L (Fig. 3B). The greenhouse-grown plants exhibited the lowest contents of total phenolics, while the levels of flavonols were similar to those detected in adventitious roots grown in the control medium (Fig. 3B). In turn, the overall amount of hydroxycinnamic acids in roots induced with yeast extract closely resembles the levels found in greenhouse-grown plants but is significantly higher than those in roots from the control experiment (Fig. 3B).
Studies have shown that endophytic fungi play a supportive role for plants by aiding in germination, enhancing shoot growth, and fortifying plants against various stressors, leading to the accumulation of beneficial bioactive secondary metabolites (Wen et al. 2022). Given the common exposure of plant roots to fungi, we selected yeast extract as an elicitor to mimic this interaction, triggering the activation of the plant's defense mechanisms to increase the production of secondary metabolites (Koprivova and Kopriva 2022). Our observations align with this trend, indicating a rise in secondary metabolite production in response to the yeast extract elicitation treatment. Moreover, the application of stressors in plant science often results in biostimulant and elicitor responses, enhancing plant performance and immunity. But these stressors may also induce some toxicity, and the effects occur in a dose-dependent manner, reflecting the concept of “hormesis”, a biphasic dose–response to an environmental agent characterized by a low dose stimulation or beneficial effect and a high dose inhibitory or toxic effect (Mattson 2008, Godínez-Mendoza et al. 2023). Our findings align with this pattern, as we observed a decline in the accumulation of biomass in adventitious roots, accompanied by a reduction in the production of secondary metabolites under elevated elicitor concentrations, which indicate an increase in stress conditions.
Given the interesting content of hydroxycinnamic acids found in golden samphire adventitious root cultures, the extracts were analyzed by UHPLC-ESI–MS/MS to discriminate and quantify the compounds present in the extracts (Table 1 and Fig. 4). As expected, most of the detected compounds were hydroxycinnamic acids, namely chlorogenic (3), caffeic (4), and p-coumaric (6) acids, as well as three di-O-caffeoylquinic acid isomers (8 – 10). Compounds of less represented groups included quinic acid (2), syringaldehyde (5), and ethyl chlorogenate (7). The most abundant compounds in golden samphire adventitious root cultures (2.82 – 7.39 µg/mg of DW) were the 3,5-di-O-caffeoylquinic acid (10), di-O-caffeoylquinic acid isomer 2 (9), and chlorogenic acid, also known as 5-caffeoylquinic acid (3). These molecules were also those more affected by elicitation: the levels of 3,5-di-O-caffeoylquinic acid (10) progressively increased with the yeast extract concentration, from 4.53 µg/mg DW in the roots grown in control medium to 7.39 µg/mg DW in roots elicited with 200 mg/L of yeast extract (Table 1).

Predominant molecules identified in golden samphire adventitious root cultures (Table 1; adapted from PubChem)
The di-O-caffeoylquinic acid isomer 2 (9) content rose from control conditions (3.22 µg/mg DW) to 4.05 µg/mg DW with a yeast extract concentration of 50 mg/L. However, the concentration declines with higher yeast extract concentrations (100 mg/L: 3.19 µg/mg DW; 200 mg/L: 1.97 µg/mg DW). The levels of di-O-caffeoylquinic acid isomer 1 (8) presented a similar trend, but the maximum content was found in roots treated with 100 mg/L of yeast extract (0.678 µg/mg DW). The concentration of chlorogenic acid (3) slightly increased from the control treatment (2.82 µg/mg DW) when compared to the yeast extract elicitation, but without considerable differences between the different yeast extract concentrations (3.25 – 3.55 µg/mg DW). Although less representative, the content of minor compounds was also influenced by elicitation. For example, quinic (2), caffeic (4), and p-coumaric (6) acids, as well as syringaldehyde (5) contents have increased with the increasing concentration of the yeast extract, reaching the highest levels with its application at 200 mg/L (Table 1). In general, all compounds were detected in lower amounts in the roots of greenhouse-grown mother plants.
Our results are in line with others disclosing that the roots of the golden samphire are rich in quinic acid derivatives, such as different isomers of caffeoylquinic acids (Ela et al. 2012). which boosts the interest of the pharmaceutical industry in developing ways to optimize its production with increased yields but reduced production time and cost (Khanam et al. 2022).
Yeast extract is considered a biotic elicitor, rich in nucleotides, proteins, amino acids, sugars, and a variety of trace elements, which positively impact plant growth, productivity, microelement composition, as well as the levels of phytohormones and other plant metabolites (Tao et al. 2023). Elicitation with yeast extract increased the production of several metabolites in different plant species, including rosmarinic acid in Thymus lotocephalus G.López & R.Morales in vitro shoot cultures, isoflavonoids in Pueraria candollei var. mirifica cell cultures, and plumbagin in Plumbago zeylanica L. root callus (Gonçalves et al. 2019; Rani et al. 2020; Singh et al. 2020). Its application was also effective in improving diverse secondary molecules in adventitious root cultures, namely different phenolics in Polygonum multiflorum Thunb (Ho et al. 2018), flavonoids in Oplopanax elatus Nakai (Jin et al. 2023), and tanshinones in Perovskia abrotanoides Kar. (Bayesteh et al. 2021).
Besides, various plant in vitro cultures have demonstrated increased production of caffeoylquinic acids, mainly of chlorogenic acid. Among these, Leonurus sibiricus AtPAP1 transgenic roots, Lonicera macranthoides and Eucomia ulmoides cell suspension cultures stand out for their remarkable productivity over 20 mg/g DW of chlorogenic acid (Skała et al. 2020). However, di- and tri caffeoylquinic acids are less prevalent in plants, which underscores the significance of advancing biotechnological tools to effectively improve and scale up their production and expanding their application in various fields (Skala et al. 2020). However, there are no reports describing this approach for any halophyte species, or even species belonging to the same genus or family of I. chrithmoides.
Effects of elicitation on hepatoprotective properties
Ethanolic extracts from roots of the golden samphire, rich in caffeoylquinic acids, have been reported with strong hepatoprotective properties in model rats (Ela et al. 2012).
Before the hepatoprotective assay, the extracts were tested alone at concentrations ranging from 3.125 to 100 µg/mL to assess their potential cytotoxicity and select the non-toxic concentrations. In this context, none of the extracts exerted significant cytotoxicity on hepatic cells, presenting cellular viabilities amongst 99 – 134% at all concentrations tested (data not shown). Moreover, the incubation of HepG2 cells with 500 mM of ethanol induced a reduction in cellular viability to 46.8% (Fig. 5). To determine the capacity of the adventitious root cultures extract’ to protect cells from ethanol-induced cytotoxicity, cells were pre-treated with the extracts before being challenged with ethanol. Application of the extracts from roots under control conditions and those elicited with yeast extract at 50 mg/L led to increased cellular viability after ethanol treatment. Notably, the latter showed the highest increase, reaching 97%, indicating a complete recovery after ethanol-induced toxicity. However, extracts from roots treated with 100 and 200 mg/L of yeast extract, as well as those from greenhouse-grown plants did not demonstrate significant hepatoprotective ability. As expected, golden samphire root extracts enriched in caffeoylquinic acid derivatives prevented cell death of hepatic cells exposed to ethanol. In fact, different isomers of caffeoylquinic acids isolated from the roots of golden samphire, including 3,5-di-O-caffeoylquinic acid, have been reported with hepatoprotective properties by decreasing the levels of serum glutamic-oxaloacetate transaminase (SGOT), serum glutamic-pyruvic transaminase (SGPT), alkaline phosphatase (ALP), and bilirubin in rats (Ela et al. 2012). Moreover, chlorogenic acid also have shown hepatoprotective effect in lipopolysaccharide (LPS)-treated rats towards glucose-6-phosphate translocase and mitochondrial respiratory chain enzymes (Hemmerle et al. 1997; Chen et al. 2019).

In vitro hepatoprotective effect of ethanol extracts of adventitious roots cultures of I. crithmoides grown into control media (MS + 1 mg/L IBA), elicited with yeast extract (YE) at three concentrations (50 mg/L, 100 mg/L and200 mg/L), and roots from greenhouse-grown plants. HepG2 hepatocarcinoma cells were pre-incubated with the extracts at the concentration of 100 µg/mL for 24 h, followed by the application of ethanol at 500 mM for 24 h. Values correspond to mean ± SEM of six independent experiments (n = 6). Columns marked with an asterisk are considered statistically different from the control (EtOH 500 mM) when analyzed by Dunnett's test (p < 0 05)
Overall, the liquid adventitious root culture of golden samphire has shown enhanced in vitro hepatoprotective capacity than roots from plants grown in soil, mainly attributed to the increased synthesis of caffeoylquinic acids after yeast extract elicitation. This highlights the potential of using in vitro liquid roots culture for an improved production of caffeoylquinic acids for possible pharmaceutical use on alcohol-associated liver toxicity.
Conclusions
This study demonstrates the successful establishment of in vitro induced adventitious root cultures from the medicinal halophyte golden samphire (I. crithmoides) and the enhanced production of caffeoylquinic acids through yeast extract elicitation in liquid condition. The ethanol extracts obtained from elicited root cultures had significant in vitro hepatoprotective properties. In summary, the results highlight the efficacy of adventitious root cultures as a valuable tool for producing bioactive compounds from medicinal plants. This is particularly evident after yeast extract elicitation of I. crithmoides roots, showcasing its potential to enhance the production of hepatoprotective caffeoylquinic acids..
Data availability
Data will be made available on request.
References
Aasim M, Khawar KM, Özcan S (2008) In vitro regeneration of red squill Urginea maritima (L.) Baker. using thidiazuron. Biotechnol Biotechnol Equip 22:925–928. https://doi.org/10.1080/13102818.2008.10817580
Alcalde MA, Perez-Matas E, Escrich A, Cusido RM, Palazon J, Bonfill M (2022) Biotic Elicitors in Adventitious and Hairy Root Cultures: A Review from 2010 to 2022. Molecules 27:5253. https://doi.org/10.3390/molecules27165253
Alves V, Pinto R, Debiasi C, Santos MC, Gonçalves JC, Domingues J (2021) Micropropagation of Corema album from adult plants in semisolid medium and temporary immersion bioreactor. Plant Cell, Tissue Organ Cult 145:641–648. https://doi.org/10.1007/s11240-021-02034-1
Atanasov AG, Waltenberger B, Pferschy-Wenzig E-M, Linder T, Wawrosch C, Uhrin P, Temml V, Wang L, Schwaiger S, Heiss EH (2015) Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol Adv 33:1582–1614. https://doi.org/10.1016/j.biotechadv201508001
Bai T, Dong Z, Zheng X, Song S, Jiao J, Wang M, Song C (2020) Auxin and its interaction with ethylene control adventitious root formation and development in apple rootstock. Front Plant Sci 11:574881. https://doi.org/10.3389/fpls2020574881
Banerjee S, Tripathi J, Verma PC, Dwivedi PD, Khanuja SPS, Bagchi GD (2004) Thidiazuron-induced high-frequency shoot proliferation in Cineraria maritima Linn. Curr Sci 33:1287–1290
Bayesteh B, Abrishamchi P, Asili J, Zaker A (2021) In vitro production of tanshinones in adventitious root cultures of Perovskia abrotanoides Kar under Ag+ ion and yeast extract elicitation. J Plant Res (iranian J Biol) 34:139–151
Bellini C, Pacurar DI, Perrone I (2014) Adventitious roots and lateral roots: similarities and differences. Annu Rev Plant Biol 65:639–666. https://doi.org/10.1146/annurev-arplant-050213-035645
Bernabé-Antonio A, Castro-Rubio C, Rodríguez-Anda R, Silva-Guzmán JA, Manríquez-González R, Hurtado-Díaz I, Sánchez-Ramos M, Hinojosa-Ventura G, Romero-Estrada A (2023) Jasmonic and salicylic acids enhance biomass, total phenolic content, and antioxidant activity of adventitious roots of Acmella radicans (Jacq ) RK Jansen Cultured in Shake Flasks. Biomolecules 13:746. https://doi.org/10.3390/biom13050746
Chen Z, Yang Y, Mi S, Fan Q, Sun X, Deng B, Wu G, Li Y, Zhou Q, Ruan Z (2019) Hepatoprotective effect of chlorogenic acid against chronic liver injury in inflammatory rats. J Funct Foods 62:103540. https://doi.org/10.1016/j.jff.2019103540
Clapham AR, Tutin TG, Warburg EF (1962) Flora of the British Isles, 2nd edn. Cambridge University Press, New York
D’Agostino G, Badalamenti N, Franco P, Bruno M, Gallo G (2022) The chemical composition of the flowers essential oil of Inula crithmoides (Asteraceae) growing in aeolian islands, Sicily (Italy) and its biocide properties on microorganisms affecting historical art crafts. Nat Prod Res 36:2993–3001. https://doi.org/10.1080/14786419.2021.1938040
Deriu A, Zanetti S, Sechi LA, Marongiu B, Piras A, Porcedda S, Tuveri E (2008) Antimicrobial activity of Inula helenium L. essential oil against Gram-positive and Gram-negative bacteria and Candida spp. Int J Antimicrob Agents 31:588–590. https://doi.org/10.1016/jijantimicag.2008.02.006
Duarte B, Feijao E, Pinto MV, Matos AR, Silva A, Figueiredo A, Fonseca VF, Reis-Santos P, Caçador I (2022) Nutritional valuation and food safety of endemic mediterranean halophytes species cultivated in abandoned salt pans under a natural irrigation scheme. Estuar Coast Shelf Sci 265:107733. https://doi.org/10.1016/j.ecss.2021.107733
eHALOPH (2023) Salt-tolerant plants. https://ehaloph.uc.pt/showplant/11358. Accessed 2 August 2023
Ela MAA, El-Lakany AM, Abdel-Kader MS, Alqasoumi SI, Shams-El-Din SM, Hammoda HM (2012) New quinic acid derivatives from hepatoprotective Inula crithmoides root extract. Helv Chim Acta 95:61–66. https://doi.org/10.1002/hlca.201100282
European Chemicals Agency (2024) Substance information. https://echa.europa.eu/substance-information/-/substanceinfo/100.248.993#OTHER_IDENTIFIERScontainer. Access 8 January 2024
Ghimire BK, Kim S-H, Yu C-Y, Chung I-M (2022) Biochemical and physiological changes during early adventitious root formation in Chrysanthemum indicum linné cuttings. Plants 11:1440. https://doi.org/10.3390/plants11111440
Godínez-Mendoza PL, Rico-Chávez AK, Ferrusquía-Jimenez NI, Carbajal-Valenzuela IA, Villagómez-Aranda AL, Torres-Pacheco I, Guevara-González RG (2023) Plant hormesis: Revising of the concepts of biostimulation, elicitation and their application in a sustainable agricultural production. Sci Total Environ 894:164883. https://doi.org/10.1016/j.scitotenv.2023.164883
Gómez-Aguirre YA, Zamilpa A, González-Cortazar M, Trejo-Tapia G (2012) Adventitious root cultures of Castilleja tenuiflora Benth as a source of phenylethanoid glycosides. Ind Crops Prod 36:188–195. https://doi.org/10.1016/j.indcrop.2011.09005
Gonçalves S, Mansinhos I, Rodríguez-Solana R, Pérez-Santín E, Coelho N, Romano A (2019) Elicitation improves rosmarinic acid content and antioxidant activity in Thymus lotocephalus shoot cultures. Ind Crops Prod 137:214–220. https://doi.org/10.1016/j.indcrop.2019.04.071
Halder M, Sarkar S, Jha S (2019) Elicitation: A biotechnological tool for enhanced production of secondary metabolites in hairy root cultures. Eng Life Sci 19:880–895. https://doi.org/10.1002/elsc.201900058
Hemmerle H, Burger H-J, Below P, Schubert G, Rippel R, Schindler PW, Paulus E, Herling AW (1997) Chlorogenic acid and synthetic chlorogenic acid derivatives: novel inhibitors of hepatic glucose-6-phosphate translocase. J Med Chem 40:137–145. https://doi.org/10.1021/jm9607360
Hernández V, Recio MC, Máñez S, Giner RM, Ríos J-L (2007) Effects of naturally occurring dihydroflavonols from Inula viscosa on inflammation and enzymes involved in the arachidonic acid metabolism. Life Sci 81:480–488. https://doi.org/10.1016/j.lfs.2007.06.006
Ho T-T, Lee J-D, Jeong C-S, Paek K-Y, Park S-Y (2018) Improvement of biosynthesis and accumulation of bioactive compounds by elicitation in adventitious root cultures of Polygonum multiflorum. Appl Microbiol Biotechnol 102:199–209. https://doi.org/10.1007/s00253-017-8629-2
Hussain MJ, Abbas Y, Nazli N, Fatima S, Drouet S, Hano C, Abbasi BH (2022) Root cultures, a boon for the production of valuable compounds: A comparative review. Plants 11:439. https://doi.org/10.3390/plants11030439
Isah T (2019) Stress and defense responses in plant secondary metabolites production. Biol Res 52. https://doi.org/10.1186/s40659-019-0246-3
Jin M-Y, Wang M, Wu X-H, Fan M-Z, Li H-X, Guo Y-Q, Jiang J, Yin C-R, Lian M-L (2023) Improving flavonoid accumulation of bioreactor-cultured adventitious roots in Oplopanax elatus using yeast extract. Plants 12:2174. https://doi.org/10.3390/plants12112174
Khanam MN, Anis M, Javed SB, Mottaghipisheh J, Csupor D (2022) Adventitious root culture—An alternative strategy for secondary metabolite production: A review. Agronomy 12:1178. https://doi.org/10.3390/agronomy12051178
Kogure K, Yamauchi I, Tokumura A, Kondou K, Tanaka N, Takaishi Y, Fukuzawa K (2004) Novel antioxidants isolated from plants of the genera Ferula, Inula, Prangos and Rheum collected in Uzbekistan. Phytomedicine 11:645–651. https://doi.org/10.1016/j.phymed.2003.09004
Koprivova A, Kopriva S (2022) Plant secondary metabolites altering root microbiome composition and function. Curr Opin Plant Biol 67:102227. https://doi.org/10.1016/j.pbi.2022.102227
Laezza C, Imbimbo P, D’Amelia V, Marzocchi A, Monti DM, di Loria A, Monti SM, Novellino E, Tenore GC, Rigano MM (2024) Use of yeast extract to elicit a pulp-derived callus cultures from Annurca apple and potentiate its biological activity. J Funct Foods 112:105988. https://doi.org/10.1016/j.jff.2023.105988
Lima AR, Gama F, Castañeda-Loaiza V, Costa C, Schüler LM, Santos T, Salazar M, Nunes C, Cruz RMS, Varela J, Barreira L (2021) Nutritional and functional evaluation of Inula crithmoides and Mesembryanthemum nodiflorum grown in different salinities for human consumption. Molecules 26:4543. https://doi.org/10.3390/molecules26154543
Liu W, Li J, Zhang X, Zu Y, Yang Y, Liu W, Xu Z, Gao H, Sun X, Jiang X (2020) Current advances in naturally occurring caffeoylquinic acids: Structure, bioactivity, and synthesis. J Agric Food Chem 68:10489–10516
Mattson MP (2008) Hormesis defined. Ageing Res Rev 7:1–7. https://doi.org/10.1016/j.arr.2007.08.007
Mazza G, Fukumoto L, Delaquis P, Girard B, Ewert B (1999) Anthocyanins, phenolics, and color of cabernet franc, merlot, and pinot noir wines from British Columbia. J Agric Food Chem 47:4009–4017
Miara MD, Souidi Z, Benhanifa K, Daikh A, Hammou MA, Moumenine A, Sabi IH (2021) Diversity, natural habitats, ethnobotany and conservation of the flora of the Macta marches (North-West Algeria). Int J Environ Stud 78:817–837. https://doi.org/10.1080/00207233.2020.1824867
Murashige T, Skoog F (1962) A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plant 15:473–497. https://doi.org/10.1111/j.1399-3054.1962tb08052.x
Nwafor E-O, Lu P, Zhang Y, Liu R, Peng H, Xing B, Liu Y, Li Z, Zhang K, Zhang Y, Liu Z (2022) Chlorogenic acid: Potential source of natural drugs for the therapeutics of fibrosis and cancer. Transl Oncol 15:101294. https://doi.org/10.1016/j.tranon.2021.101294
Oladipo A, Enwemiwe V, Ejeromedoghene O, Adebayo A, Ogunyemi O, Fu F (2022) Production and functionalities of specialized metabolites from different organic sources. Metabolites 12:534. https://doi.org/10.3390/metabo12060534
Pathak AR, Patel SR, Joshi AG, Shrivastava N, Sindhav G, Sharma S, Ansari H (2023) Elicitor mediated enhancement of shoot biomass and lupeol production in Hemidesmus indicus (L.) R. Br. ex. Schult. and Tylophora indica (Burm. F.) Merrill using yeast extract and salicylic acid. Nat Prod Res 37:1767–1773. https://doi.org/10.1080/14786419.2022.2119388
Ramirez-Estrada K, Vidal-Limon H, Hidalgo D, Moyano E, Golenioswki M, Cusidó RM, Palazon J (2016) Elicitation, an effective strategy for the biotechnological production of bioactive high-added value compounds in plant cell factories. Molecules 21:182. https://doi.org/10.3390/molecules21020182
Rani D, Meelaph T, De-Eknamkul W, Vimolmangkang S (2020) Yeast extract elicited isoflavonoid accumulation and biosynthetic gene expression in Pueraria candollei var mirifica cell cultures. Plant Cell, Tissue Organ Cult 141:661–667. https://doi.org/10.1007/s11240-020-01809-2
Rodrigues MJ, Soszynski A, Martins A, Rauter AP, Neng NR, Nogueira JMF, Varela J, Barreira L, Custódio L (2015) Unravelling the antioxidant potential and the phenolic composition of different anatomical organs of the marine halophyte Limonium algarvense. Ind Crops Prod 77:315–322. https://doi.org/10.1016/J.INDCROP.2015.08.061
Rodrigues MJ, Castañeda-Loaiza V, Monteiro I, Pinela J, Barros L, Abreu RMV, Oliveira MC, Reis C, Soares F, Pousão-Ferreira P (2021) Metabolomic profile and biological properties of sea lavender (Limonium algarvense Erben) plants cultivated with aquaculture wastewaters: Implications for its use in herbal formulations and food additives. Foods 10:3104. https://doi.org/10.3390/foods10123104
Rodrigues MJ, Castañeda-Loaiza V, Fernandes E, Custódio L (2023) A first approach for the micropropagation of the edible and medicinal halophyte Inula crithmoides L. Plants 12:2366. https://doi.org/10.3390/plants12122366
Salam U, Ullah S, Tang Z-H, Elateeq AA, Khan Y, Khan J, Khan A, Ali S (2023) Plant Metabolomics: An Overview of the Role of Primary and Secondary Metabolites against Different Environmental Stress Factors. Life 13:706. https://doi.org/10.3390/life13030706
Saygi Ş, Konuklugil B, Kutsal O, Uzbay İT, Deniz G, Gören Z (2003) Assessment of therapeutic effect of Inula heterolepsis Boiss in alcoholic rats. Phyther Res 17:683–687. https://doi.org/10.1002/ptr.1221
Singh T, Sharma U, Agrawal V (2020) Isolation and optimization of plumbagin production in root callus of Plumbago zeylanica L. augmented with chitosan and yeast extract. Ind Crops Prod 151:112446. https://doi.org/10.1016/j.indcrop.2020.112446
Skała E, Makowczyńska J, Wieczfinska J, Kowalczyk T, Sitarek P (2020) Caffeoylquinic acids with potential biological activity from plant in vitro cultures as alternative sources of valuable natural products. Curr Pharm Des 26:2817–2842. https://doi.org/10.2174/1381612826666200212115826
Tao Z, Yuan H, Liu M, Liu Q, Zhang S, Liu H, Jiang Y, Huang D, Wang T (2023) Yeast extract: characteristics, production, applications and future perspectives. J Microbiol Biotechno 33:151–166. https://doi.org/10.4014/jmb.2207.07057
Velioglu YS, Mazza G, Gao L, Oomah BD (1998) Antioxidant activity and total phenolics in selected fruits, vegetables and grain products. J Agric Food Chem 46:4113–4117
Wawrosch C, Zotchev SB (2021) Production of bioactive plant secondary metabolites through in vitro technologies—status and outlook. Appl Microbiol Biotechnol 105:6649–6668. https://doi.org/10.1007/s00253-021-11539-w
Wen J, Okyere SK, Wang S, Wang J, Xie L, Ran Y, Hu Y (2022) Endophytic Fungi: An Effective Alternative Source of Plant-Derived Bioactive Compounds for Pharmacological Studies. J Fungus 8:205. https://doi.org/10.3390/jof8020205
Yuan F, Chen M, Yang J, Leng B, Wang B (2014) A system for the transformation and regeneration of the recretohalophyte Limonium bicolor. Vitr Cell Dev Biol 50:610–617. https://doi.org/10.1007/s11627-014-9611-7
Zou JY, Chang SKC, Gu Y, Qian SY (2011) Antioxidant activity and phenolic compositions of lentil (Lens culinaris var. Morton) extract and its fractions. Agric Food Chem 59:2268–2276
Zurayk RA, Baalbaki R (1996) Inula crithmoides: a candidate plant for saline agriculture. Arid L Res Manag 10:213–223. https://doi.org/10.1080/15324989609381436
Funding
Open access funding provided by FCT|FCCN (b-on). This study received Portuguese national funds from FCT—Foundation for Science and Technology through projects UIDB/04326/2020 (https://doi.org/10.54499/UIDB/04326/2020), UIDP/04326/2020 (https://doi.org/10.54499/UIDP/04326/2020) and LA/P/0101/2020 (https://doi.org/10.54499/LA/P/0101/2020). M.J.R. was supported by the FCT program contract (UIDP/04326/2020), L.C. by the FCT Scientific Employment Stimulus (CEECIND/00425/2017), and V.C.-L. acknowledge FCT for the PhD grant (2020.04541.BD).
Author information
Authors and Affiliations
Contributions
Luísa Custódio: Conceptualization, formal analysis, investigation, resources, writing – original draft preparation, review and editing, project administration, funding acquisition. Zóltan Cziáky: methodology, formal analysis, writing – review and editing. Viana Castañeda-Loaiza: methodology. Maria João Rodrigues: Conceptualization, methodology, formal analysis, investigation, writing – original draft preparation, review and editing. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Communicated by Agnieszka Szopa.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Custódio, L., Cziáky, Z., Castañeda-Loaiza, V. et al. Establishment and elicitation of liquid adventitious root cultures of Inula crithmoides L. for increased caffeoylquinic acids production and hepatoprotective properties. Plant Cell Tiss Organ Cult 156, 59 (2024). https://doi.org/10.1007/s11240-024-02690-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11240-024-02690-z