Butlletí de la Institució Catalana d’Història Natural
85 (1-4)
Barcelona 2021

Butlletí de la Institució Catalana d’Història Natural
85 (1- 4)
Barcelona 2021
INSTITUCIÓ CATALANA D’HISTÒRIA NATURAL
Editor en Cap
Juli Pujade-Villar, Universitat de Barcelona, Facultat de Biologia, Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals (Secció invertebrats), Barcelona.
Coeditors
Albert Masó, Universitat de Barcelona, Facultat de Biologia, Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals (Secció ecologia), Barcelona
Joan Pino, Centre de Recerca Ecològica i Aplicacions Forestals (CREAF), Universitat Autònoma de Barcelona, Bellaterra. Llorenç Sáez, Unitat de Botànica, Facultat de Ciències, Universitat Autònoma de Barcelona, Bellaterra. Amador Viñolas, Corsorci del Museu de Ciències Naturals de Barcelona, Laboratori de Natura, Coŀlecció d’artròpodes, Barcelona.
L’edició d’aquest Butlletí ha estat possible gràcies al suport de l’Institut d’Estudis Catalans
Figura de la coberta: Espermateca de Parvospeonomus cruillensis n. sp.. Dibuix d’Amador Viñolas.
Aquesta publicació es diposita, per donar compliment a l’Esmena als articles 8, 9, 10, 21 i 78 de el Codi Internacional de Nomenclatura Zoològica (ed. 1999), referents a l’ampliació i perfeccionament dels mètodes de publicació en els repositoris en línia Internet arxive (http://www.archive.org) i Biotaxa (http://www.biotaxa.org/index/index), amb enllaços a la pròpia pàgina de la publicació, en el lloc web: https://ichn2.iec.cat/Butlleti_85.htm i https://publicacions.iec.cat/PopulaFitxa.do?moduleName=revistes_cientifiques&subModu leName=&idColleccio=162
Data de publicació online volum 85 (1): 31 de març de 2021. Pàgines: 3-40.
Data de publicació online volum 85 (2): 30 de juny de 2021. Pàgines: 41-82.
Data de publicació online volum 85 (3): 30 de setembre de 2021. Pàgines: 83-162.
Data de publicació online volum 85 (4): 30 de desembre de 2021. Pàgines: 163-230,
Data de publicació volum 85 (1-4): 30 de desembre de 2021
© Els autors dels articles
Aquesta edició és propietat de la Institució Catalana d’Història Natural (filial de l’Institut d’Estudis Catalans)
Carrer del Carme, 47. 08001 Barcelona
Compost per Amador Viñolas
Imprès a Ediciones Gráficas Rey, SL
ISSN (edició electrònica): 2013-3987
ISSN (edició impresa): 1133-6889
Dipòsit Legal: B 36100-1974
Els continguts del Butlletí de la Institució Catalana d’Història Natural estan subjectes —llevat que s’indiqui el contrari en el text, en les fotografies o en altres il·lustracions— a una llicència Reconeixement - No comercial - Sense obres derivades 3.0 Espanya de Creative Commons, el text complet de la qual es pot consultar a http://creativecommons.org/licenses/by-nc-nd/3.0/es/deed.ca. Així, doncs, s’autoritza el públic en general a reproduir, distribuir i comunicar l’obra sempre que se’n reconegui l’autoria i l’entitat que la publica i no se’n faci un ús comercial ni cap obra derivada.
Distribució de macrolíquens a Catalunya amb noves dades corològiques sobre líquens protegits
Artur Lluent* & Antonio Gómez-Bolea**, ***
* Forestal Catalana S.A. Torrent de l’Olla 218-220 1er. 08012 Barcelona, Spain.
** Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals. Universitat de Barcelona. 08028 Barcelona, Spain.
*** Institut de Recerca de la Biodiversitat (IRBio). Universitat Barcelona. 08028, Barcelona, Spain.
Autor per a la correspondència: Artur Lluent. A/e: artur.lluent@gencat.cat
Rebut: 14.12.2020; Acceptat: 23.03.2021; Publicat: 31.03.2021
Resum
En el present treball s’aporten noves dades coroŀlògiques de 39 tàxons liquènics entre ells la primera cita per Catalunya de Nephroma tangeriense. S’afegeixen noves localitats per Seirophora lacunosa, considerada com en perill d’extinció al catàleg de flora amenaçada de Catalunya i també per 16 de les 20 espècies de líquens incloses com a vulnerables. Per a cada espècie s’ha elaborat un mapa on es mostren les localitats conegudes fins ara i les que s’afegeixen en aquest treball.
Paraules clau: Nephroma tangeriense, líquens, distribució, espècies vulnerables.
Abstract
Distribution of macrolichens in Catalonia with new chorological data on protected lichens
This paper contributes with new chorological data of 39 lichen taxa amongst them the first report for Catalonia of Nephroma tangeriense. New localities are added for Seirophora lacunosa, considered in extinction risk in the catalogue of endangered flora of Catalonia, and in addition 16 of 20 lichen species included as vulnerable. The chorology of each species is represented on a map where it has been differentiated old data and the new data we provide in this paper.
Key words: Nephroma tangeriense, lichens, distribution, threatened species.
Introducció
La informació corològica de les plantes vasculars es pot considerar força detallada a Catalunya, en canvi els líquens han estat molt menys prospectats i en conseqüència hi ha molt poques espècies de les que es pugui dir que se’n té un bon coneixement de la seva distribució a Catalunya.
El concepte de «macrolíquens» és completament subjectiu i s’han proposat diverses definicions. Per uns, fa referència a la forma de creixement del liquen, independentment de la mida. En aquest sentit serien macrolíquens tots els que presenten tal·lus foliaci o fruticulós i microlíquens els que tenen tal·lus crustacis. Pels que defensen aquesta postura ho tindrien complicat per convèncer-nos de que Xalocoa ocellata, que l’hem vist fent taŀlus de 40 cm de diàmetre, és un «microliquen». Per altres el concepte «macroliquen» va relacionat amb la mida del taŀlus, independentment de la forma de creixement. Serien macrolíquens tots els que són identificables a ull nu, sense necessitat de lupa. Pels que defensen aquesta definició, té sentit parlar de taŀlus microfruticolosos i de taŀlus microfoliàcis. En aquest treball, nosaltres utilitzem el concepte «macrolíquens» en aquest darrer sentit.
La dificultat en la identificació d’un tàxon liquènic no sempre està directament relacionada amb la mida o la forma de creixement. En el nostre cas, tenim l’exemple del gènere Bryoria, on per la identificació de les espècies és necessari la utilització de reactius químics i fins i tot és necessari la cromatografia en capa fina per identificar les substàncies liquèniques que caracteritzen les diferents espècies. O bé necessitem d’una lupa, encara que siguin macrolíquens, per observar certs caràcters com és el cas de les espècies de Peltigera
Material i mètodes
Les presents aportacions, majoritàriament són fruit de nombroses sortides de camp, fetes principalment per un dels autors (A. Lluent), a la recerca dels tàxons liquènics considerats en perill d’extinció. Les citacions que donem provenen de material recoŀlectat o bé de l’observació de camp per l’especialista sense material recoŀlectat. En tots dos casos, s’especifica el o els responsables de la citació. En el primer cas, a més s’han fet plecs d’herbari que han quedat dipositats a l’herbari BCN.
3 Butlletí ICHN 85 (1), 2021 GEA, FLORA ET FAUNA GEA, FLORA ET FAUNA
Butlletí de la Institució Catalana d’Història Natural, 85 (1): 3-17. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition) DOI: 10.2436/20.1502.01.67
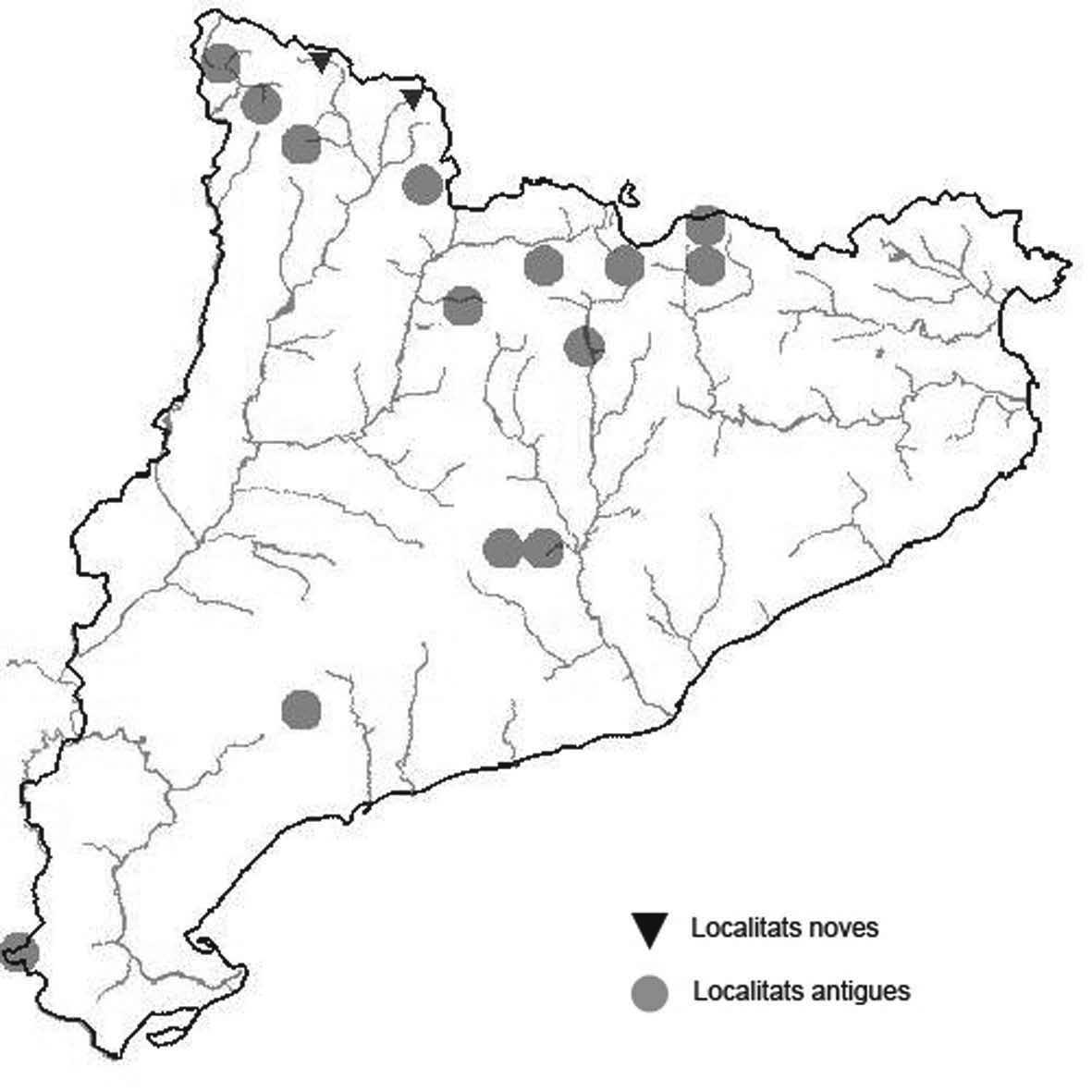
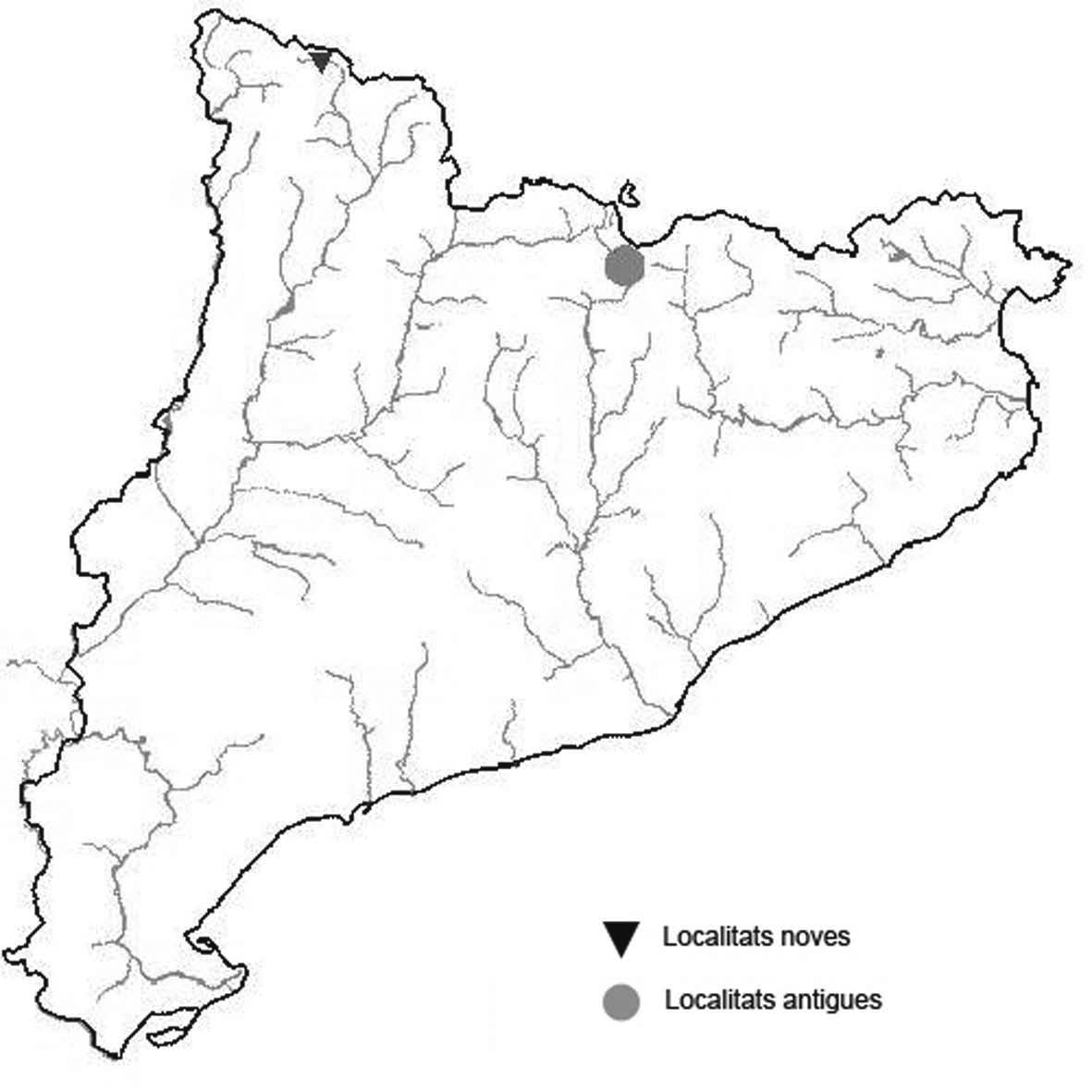
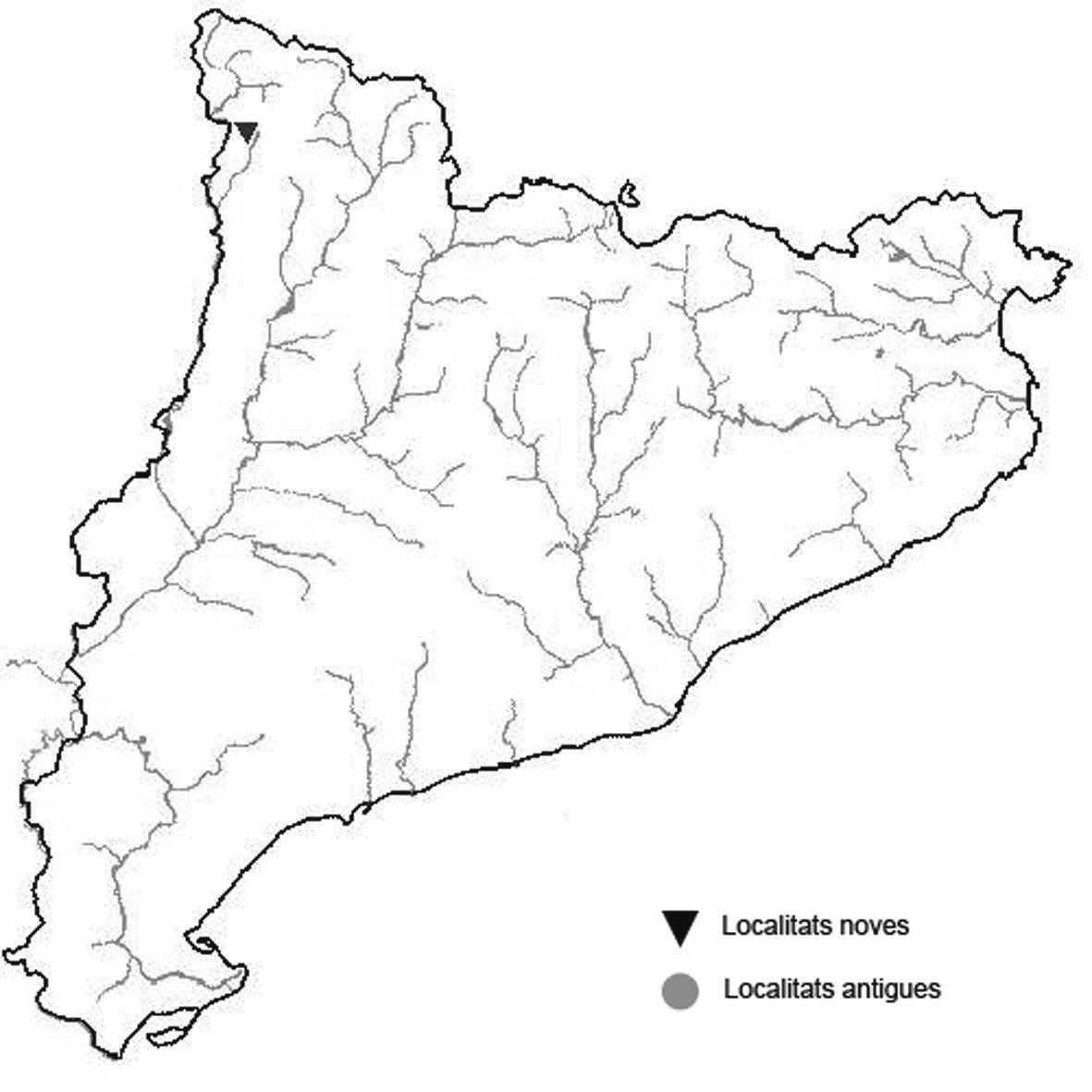
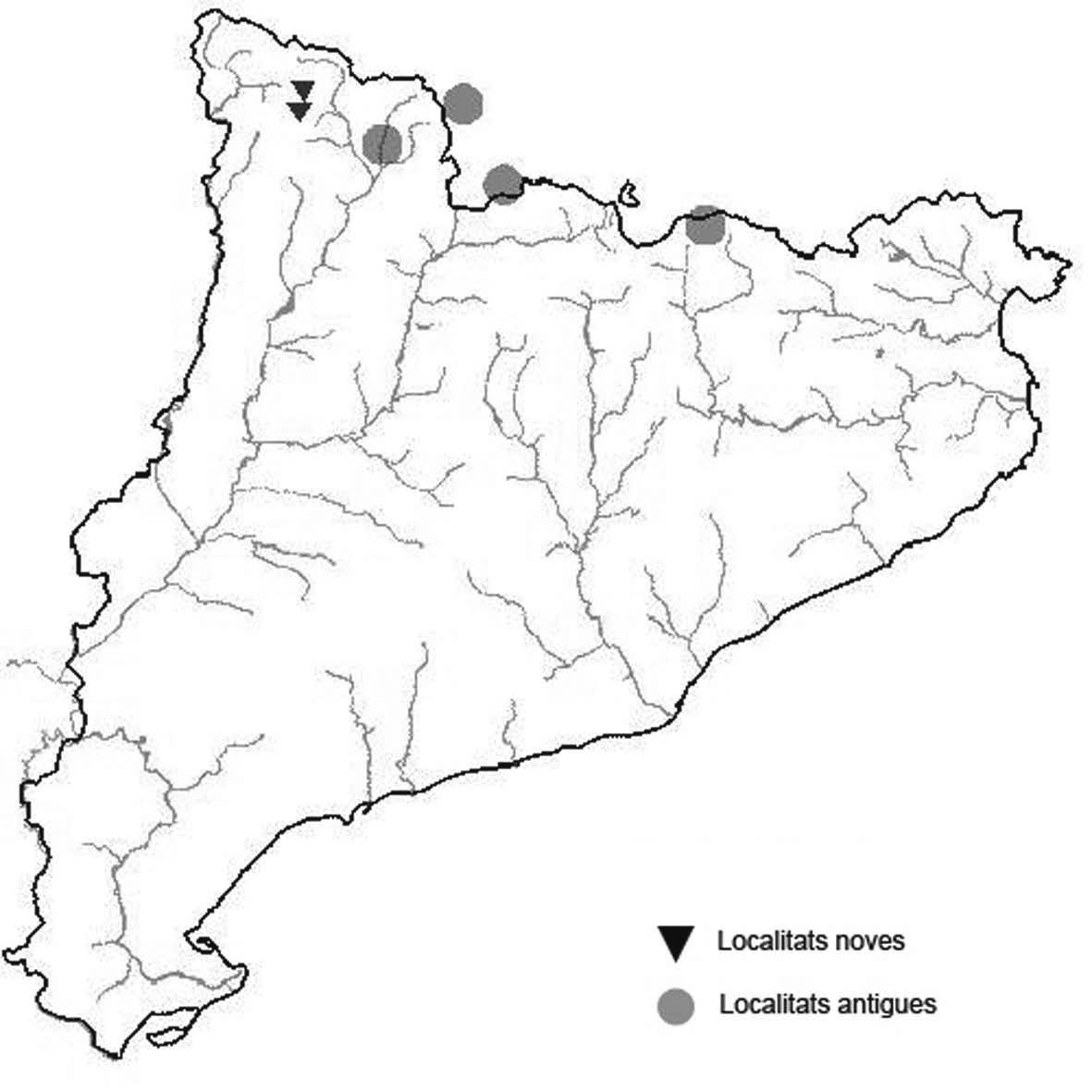
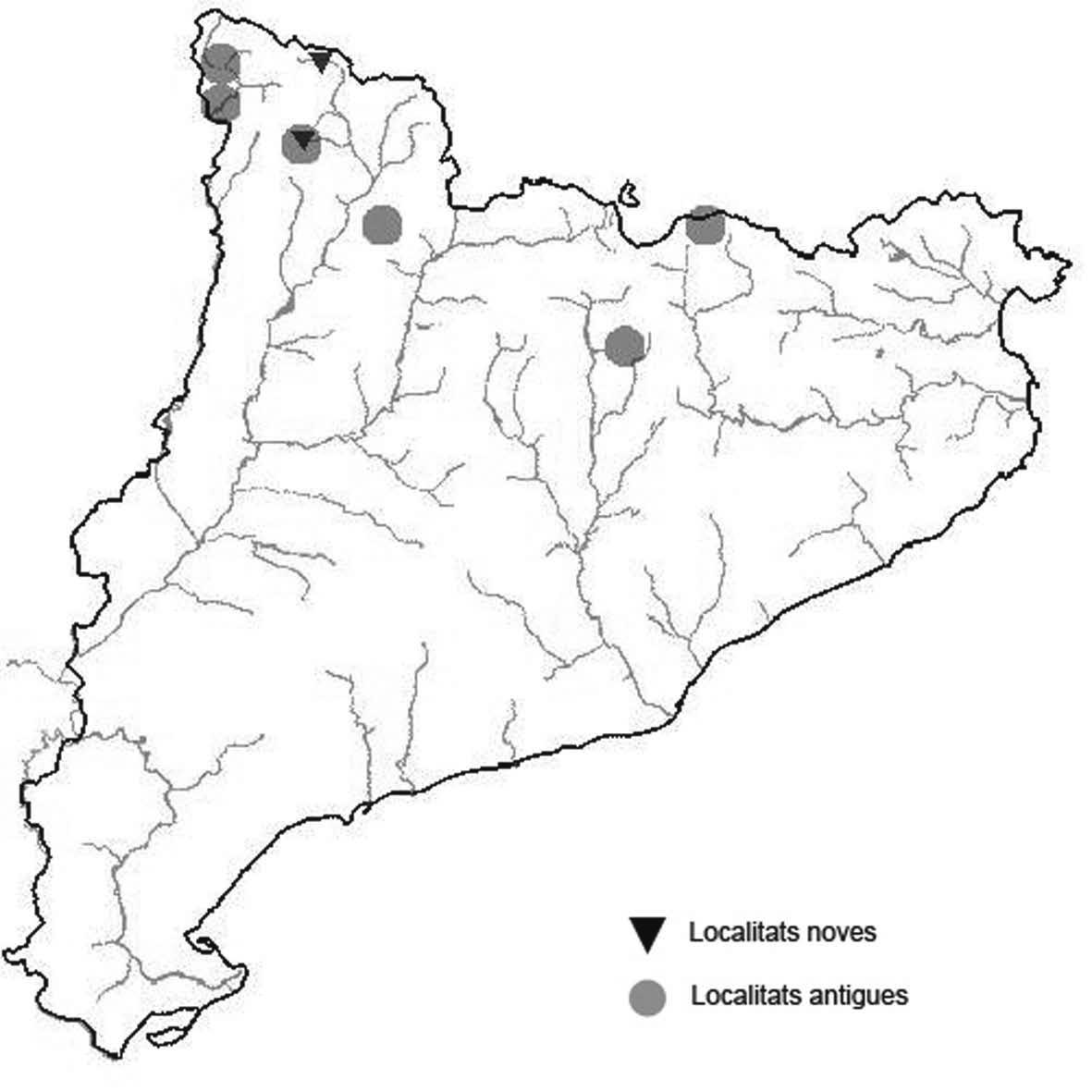
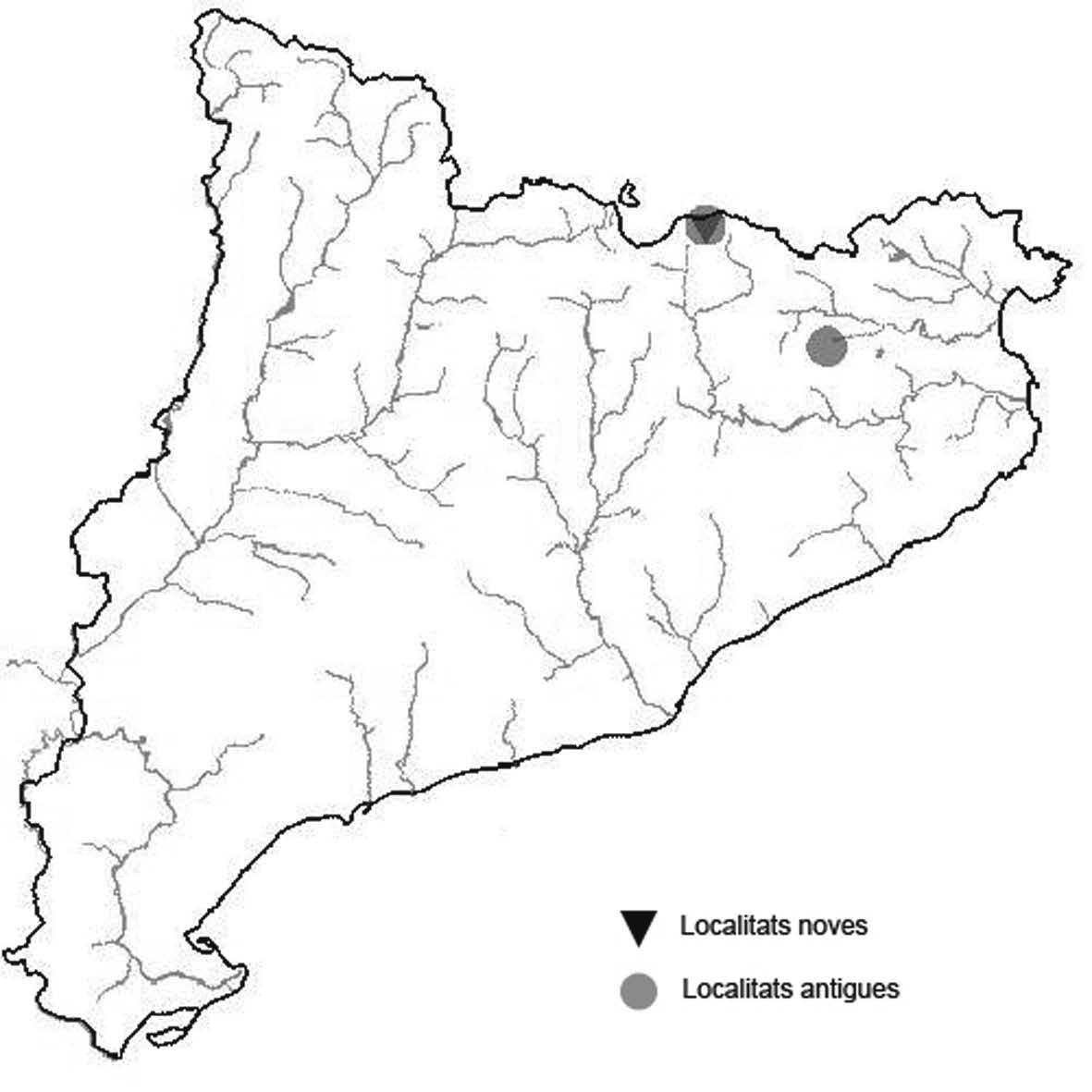
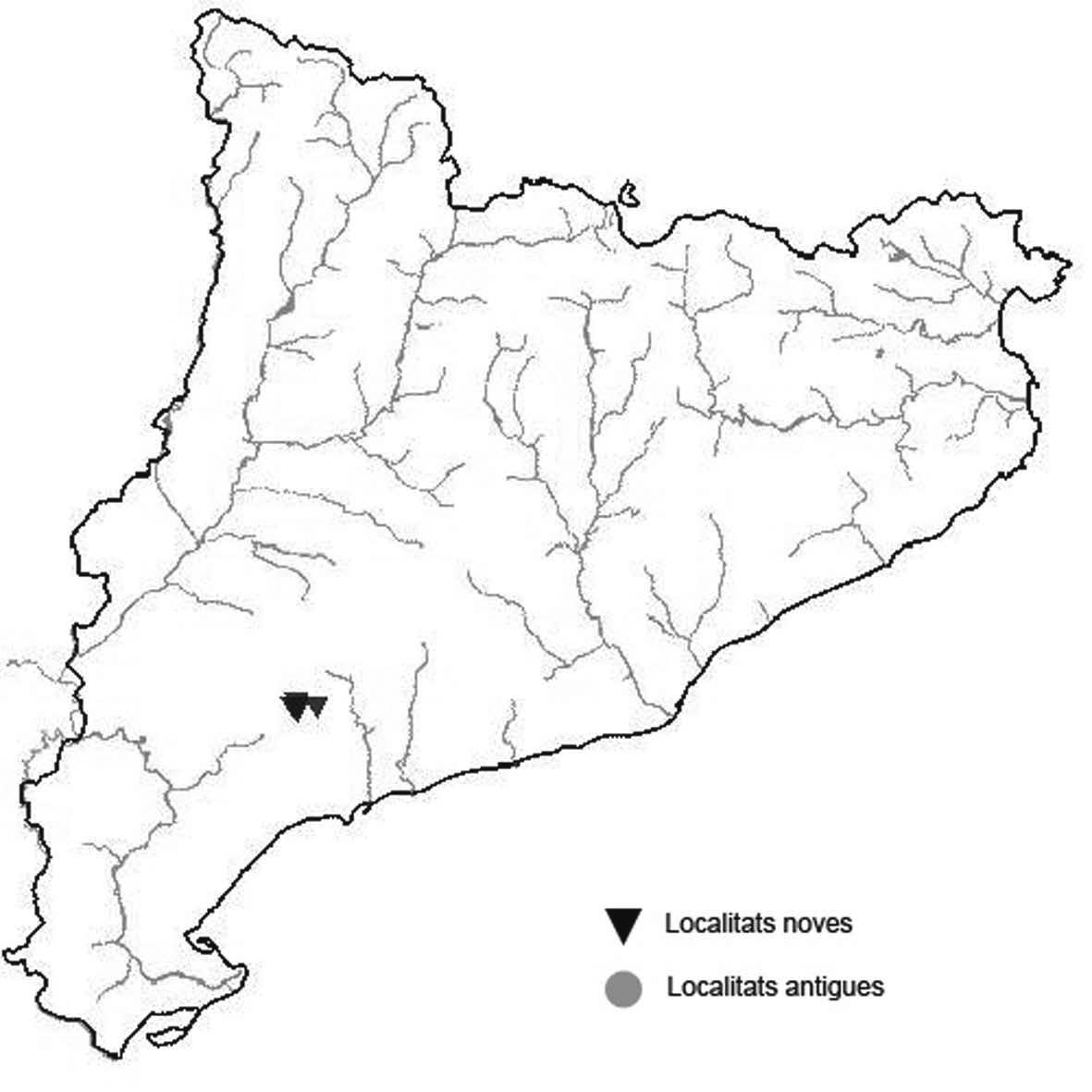
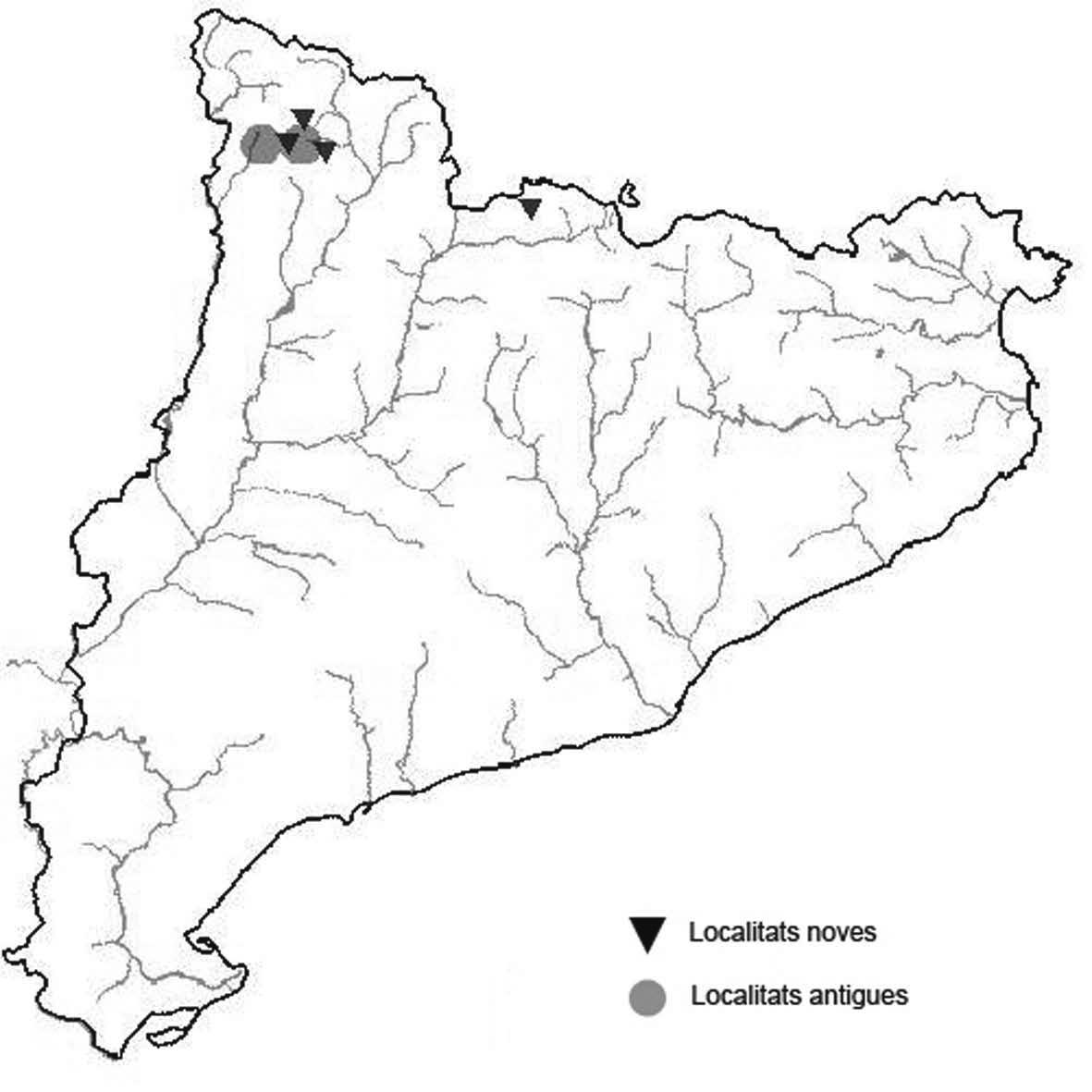
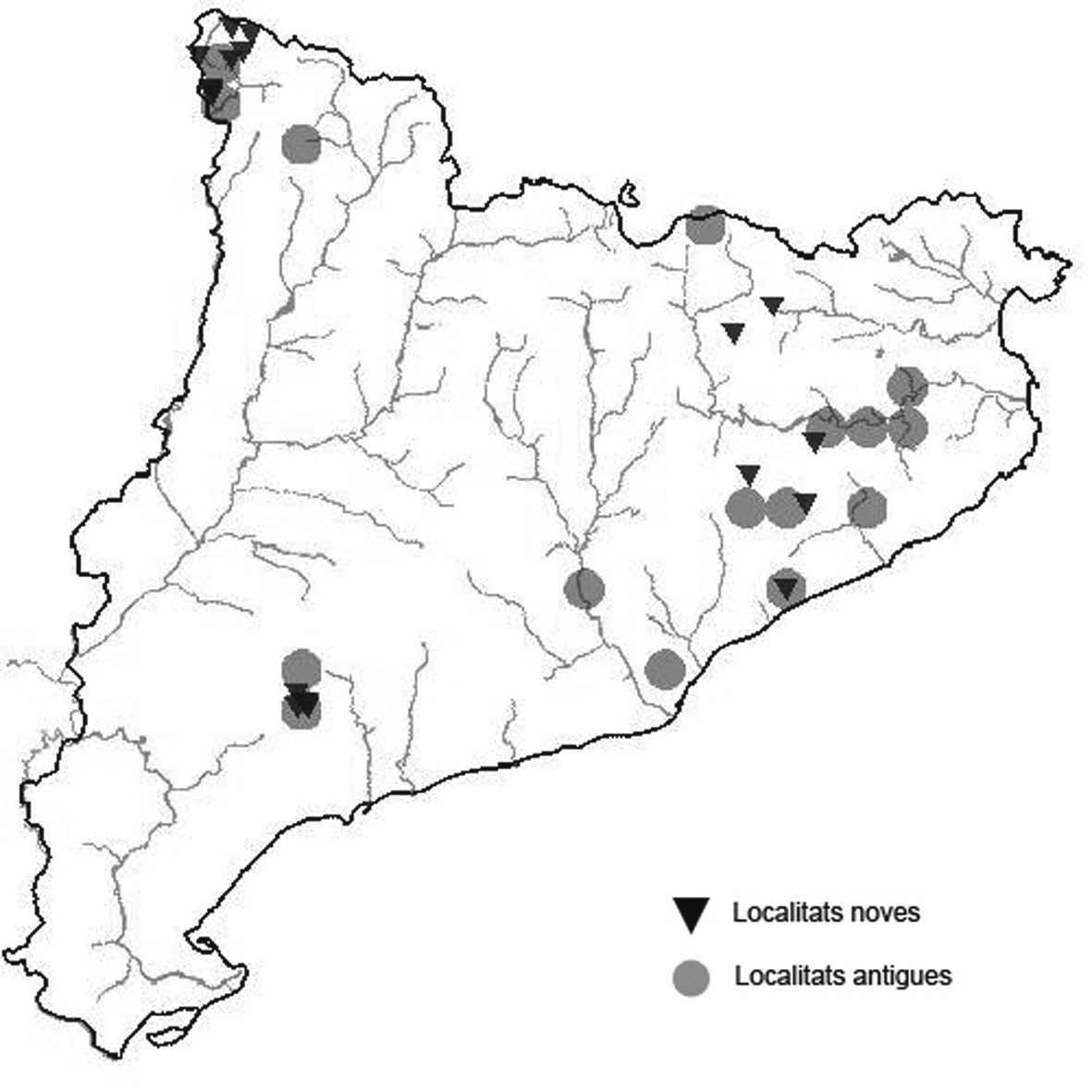
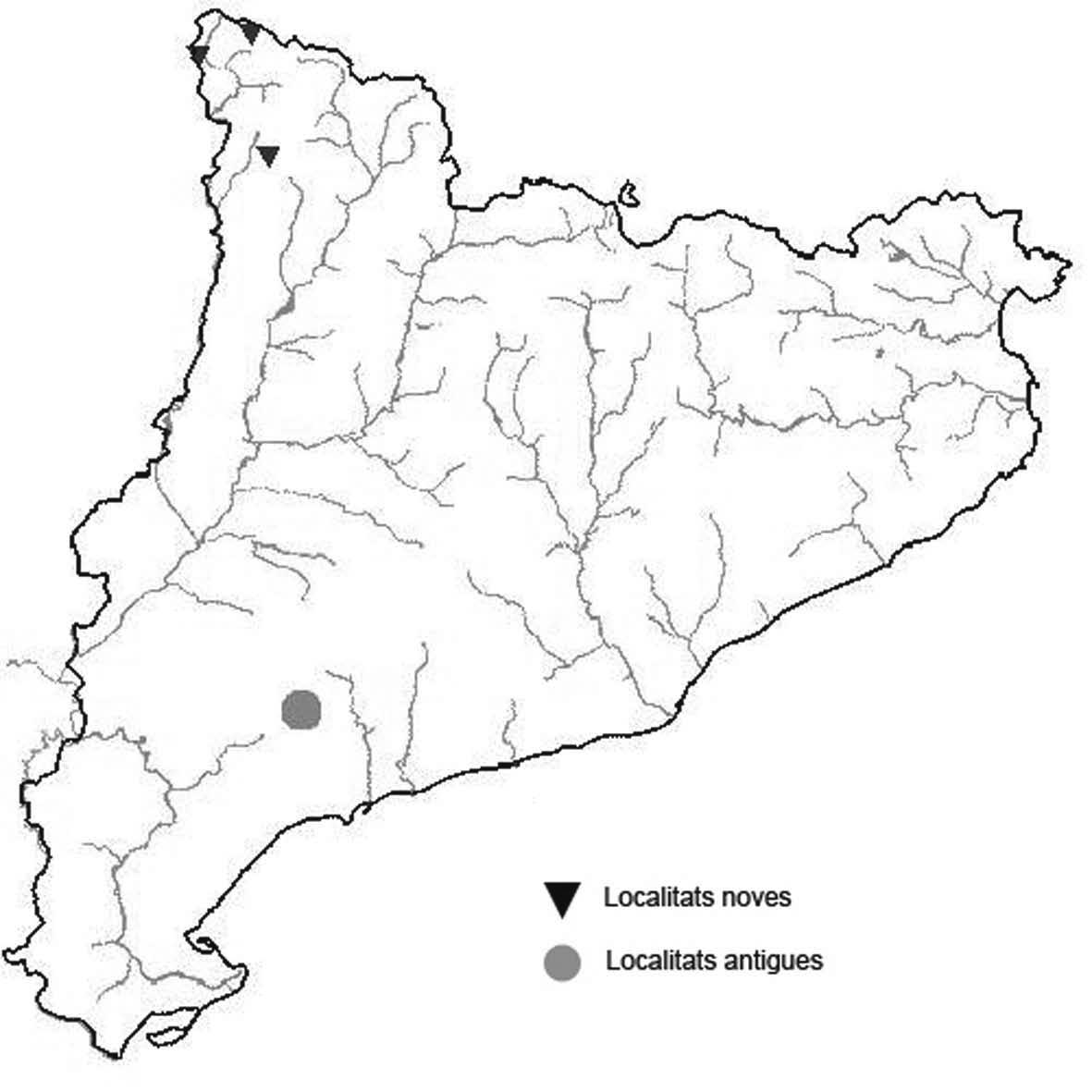
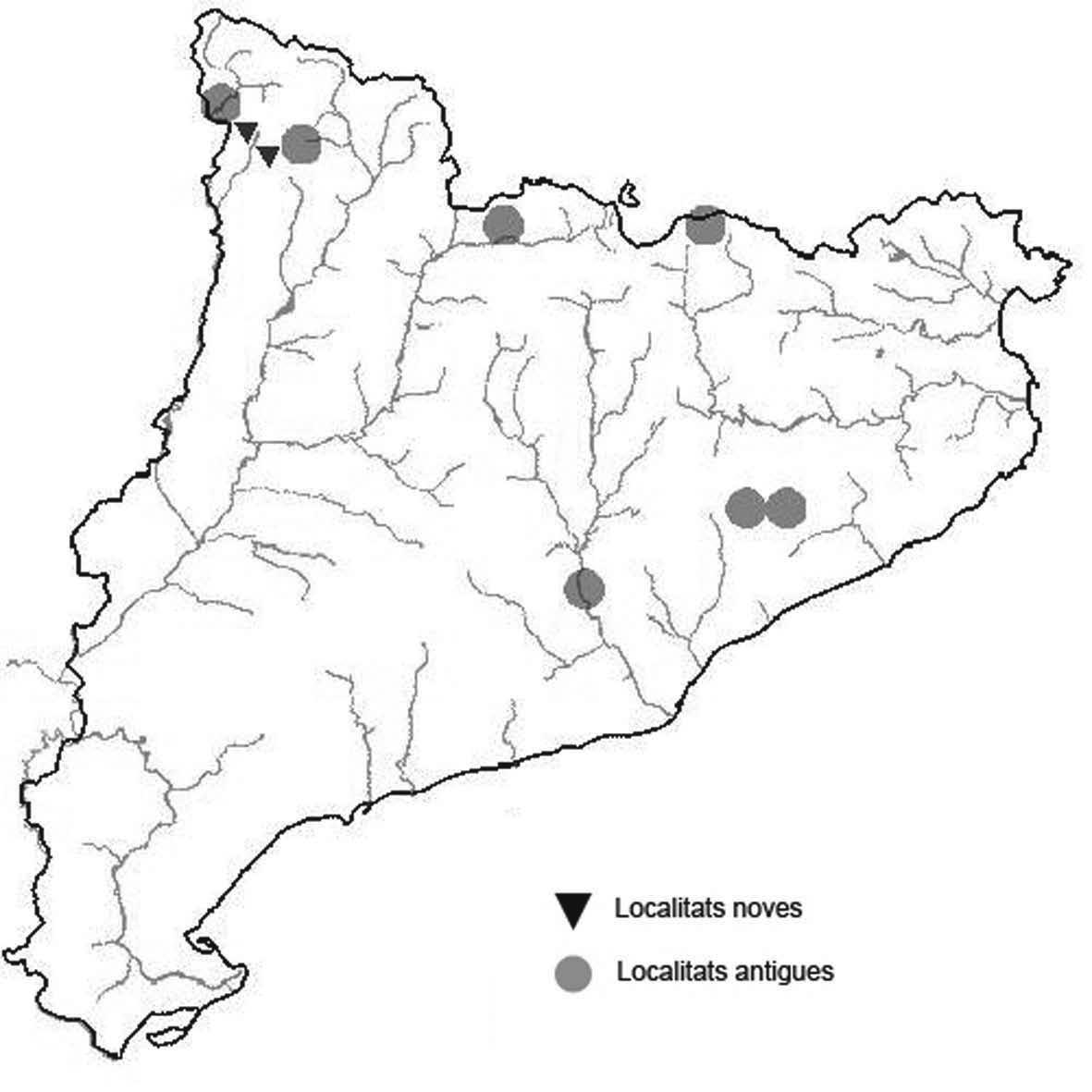
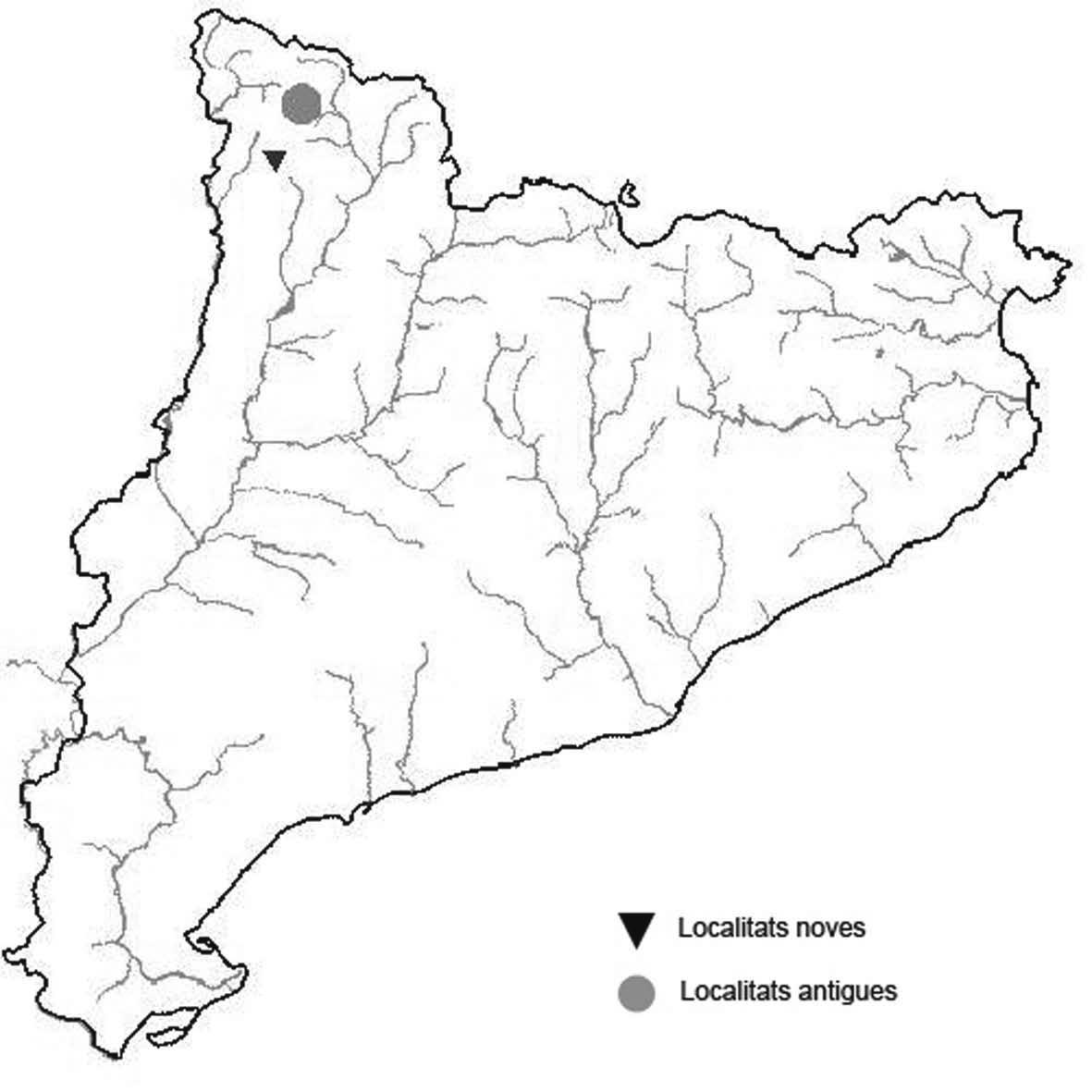
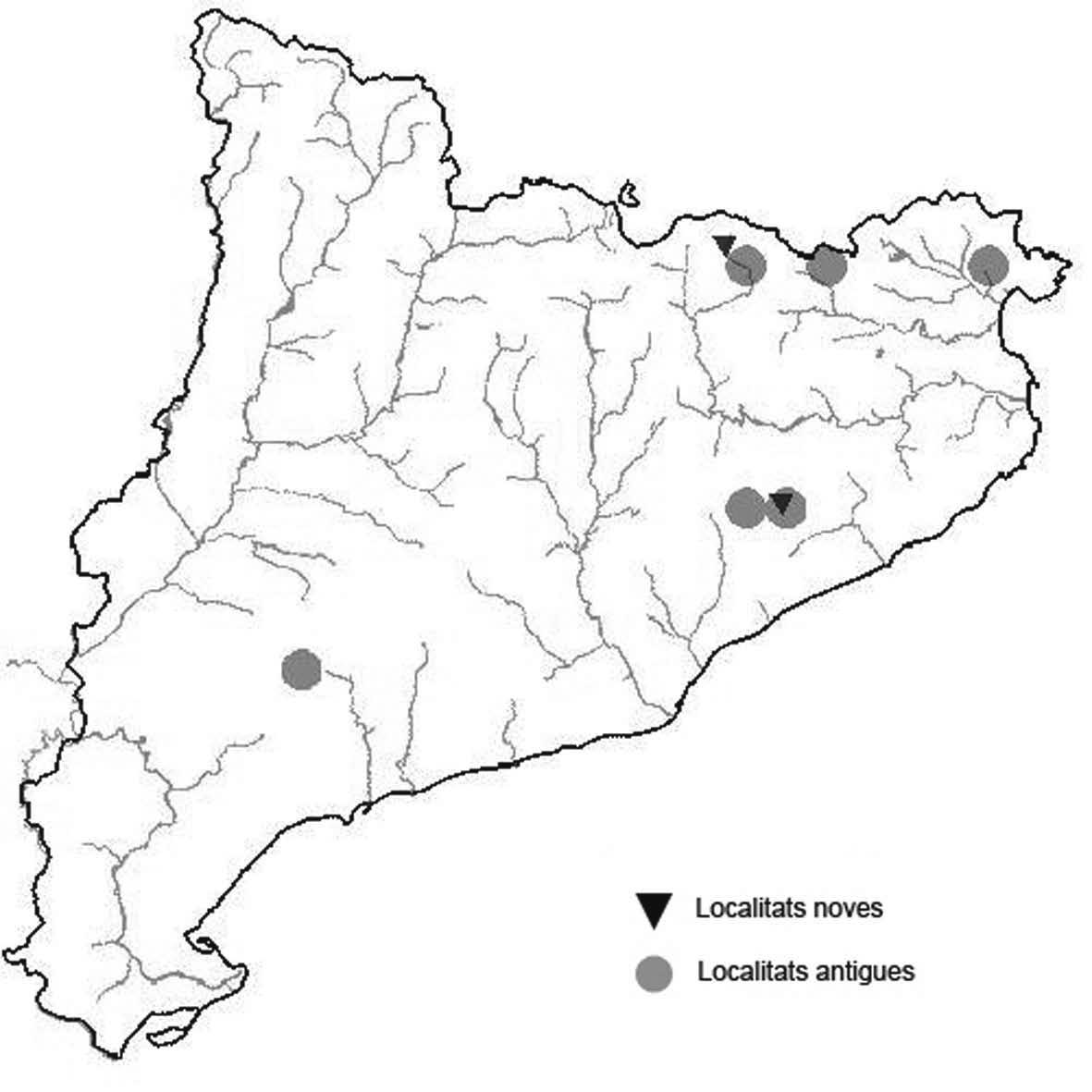
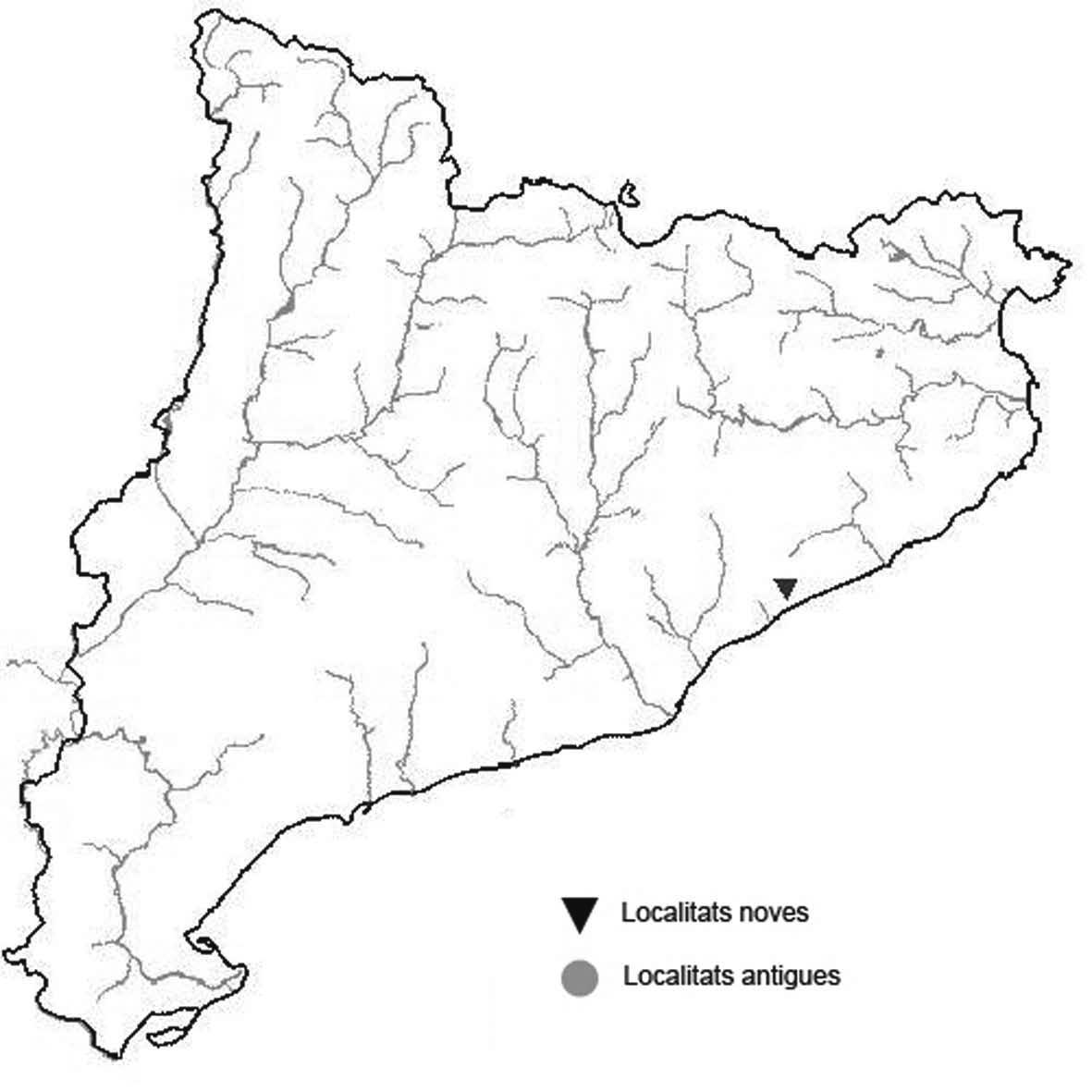
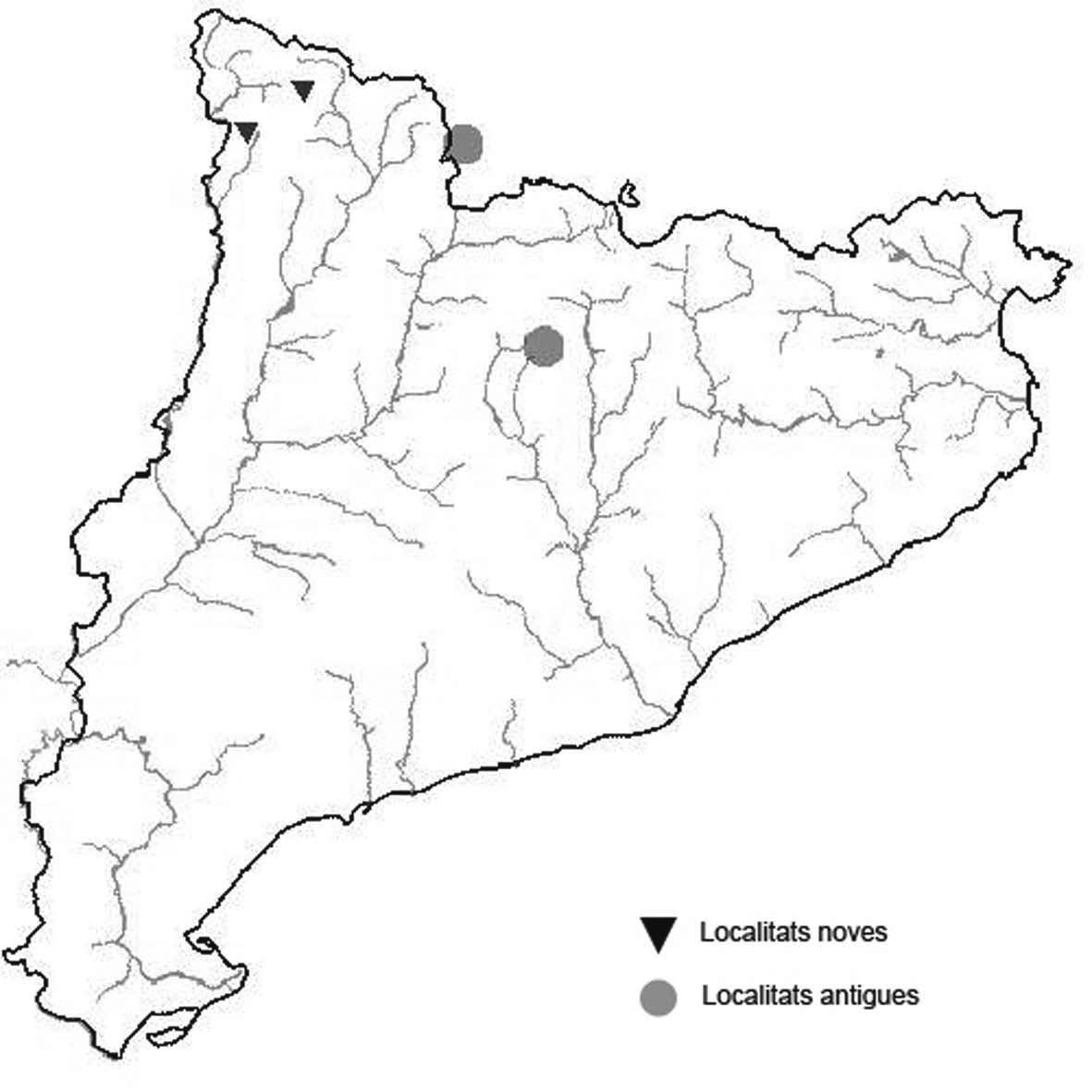
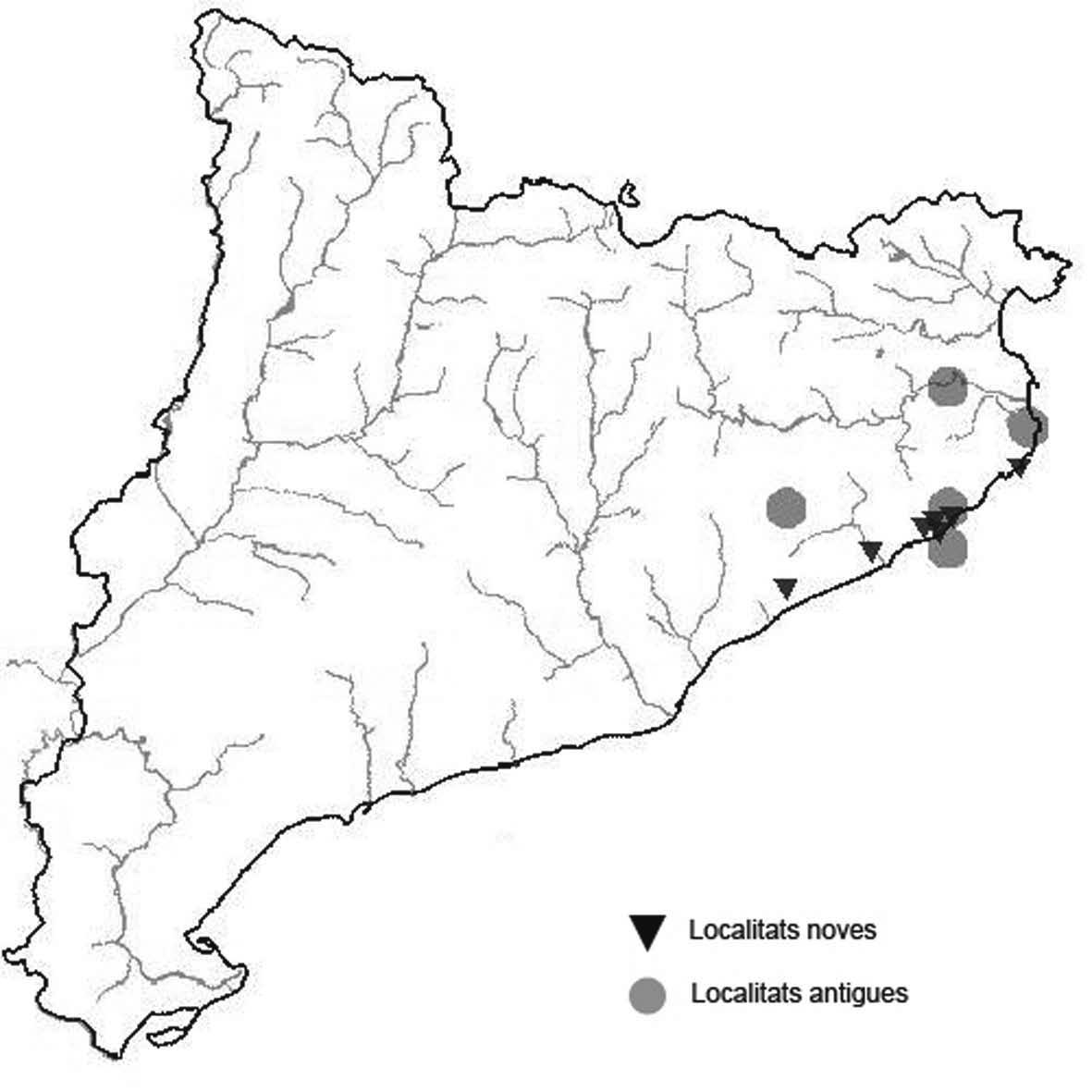
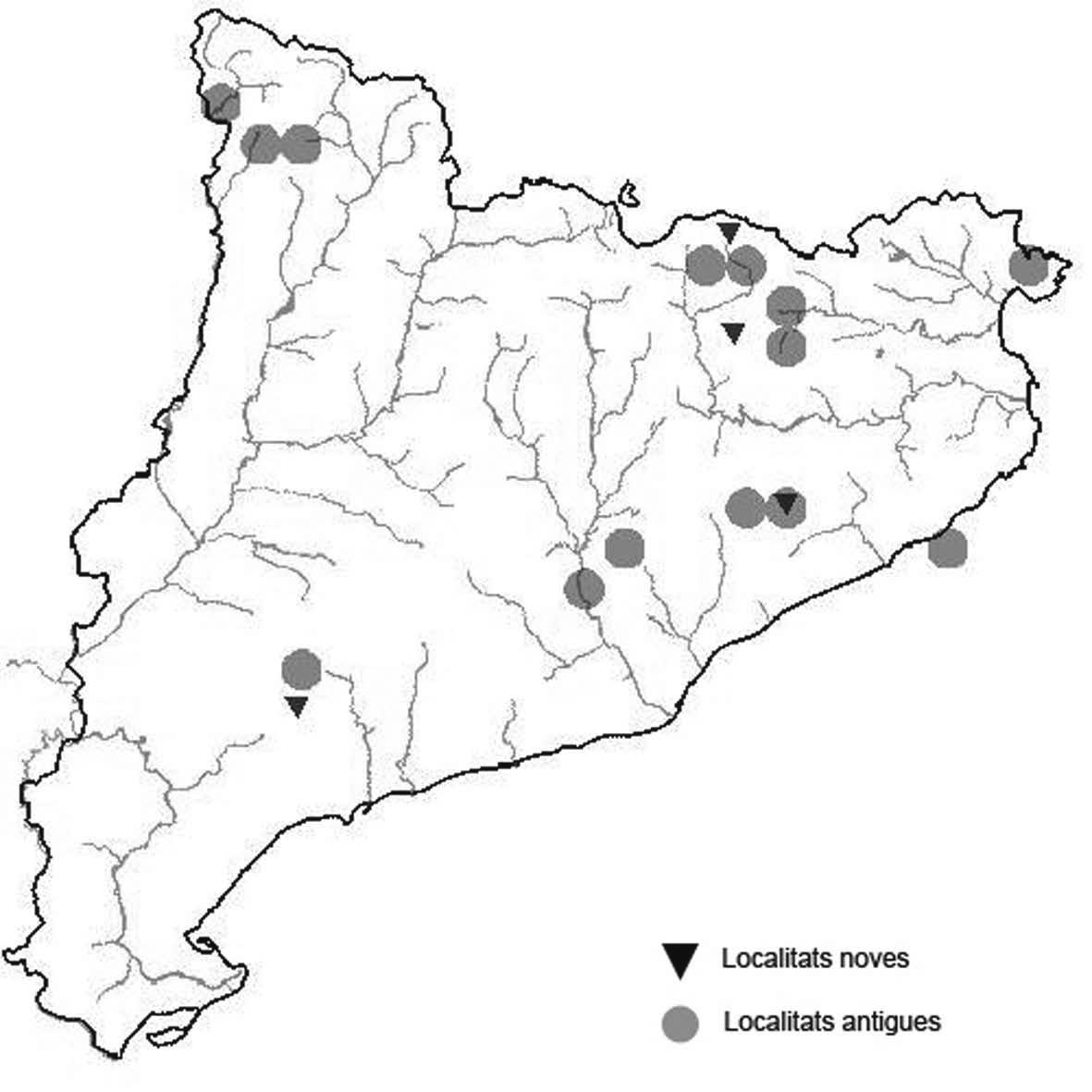
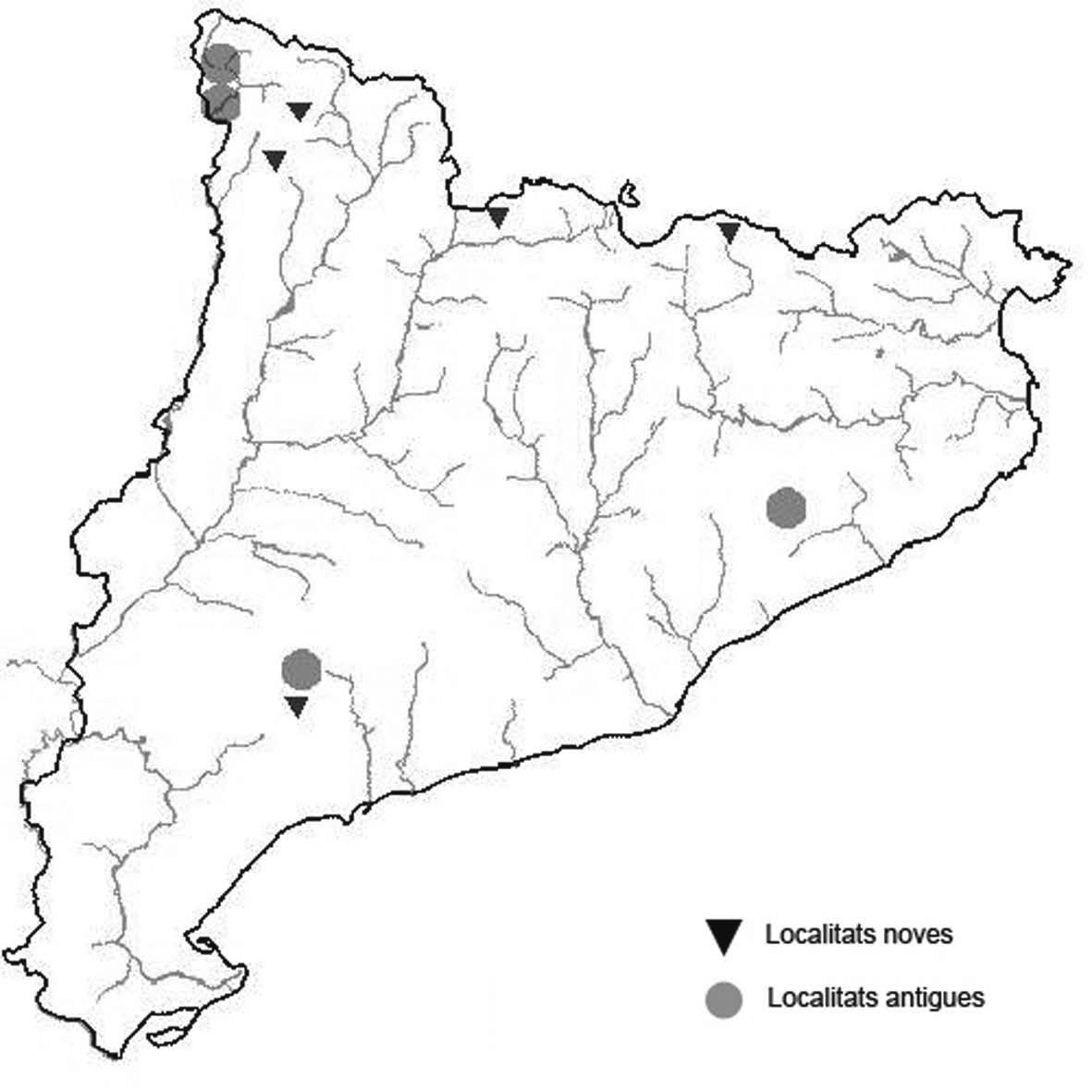
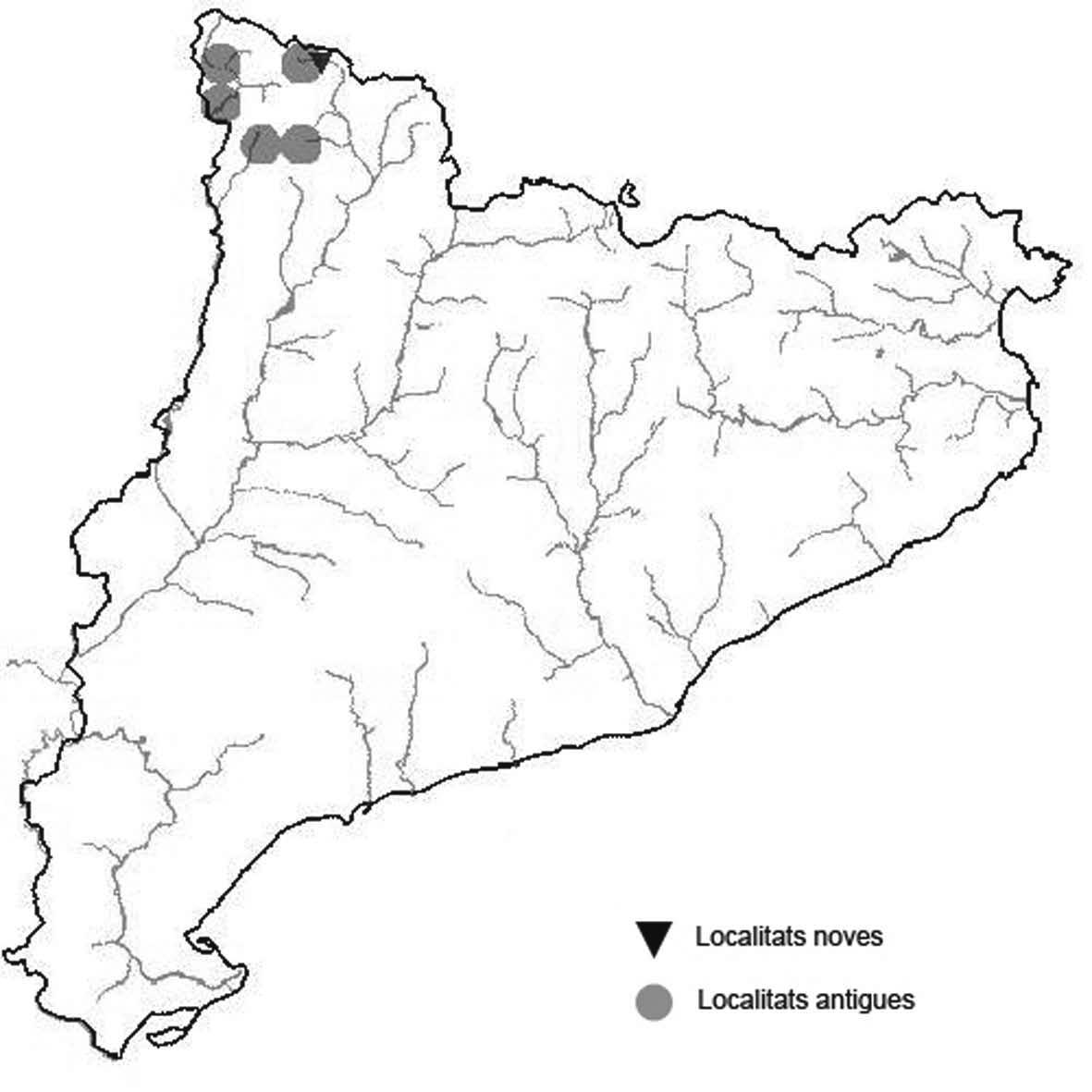
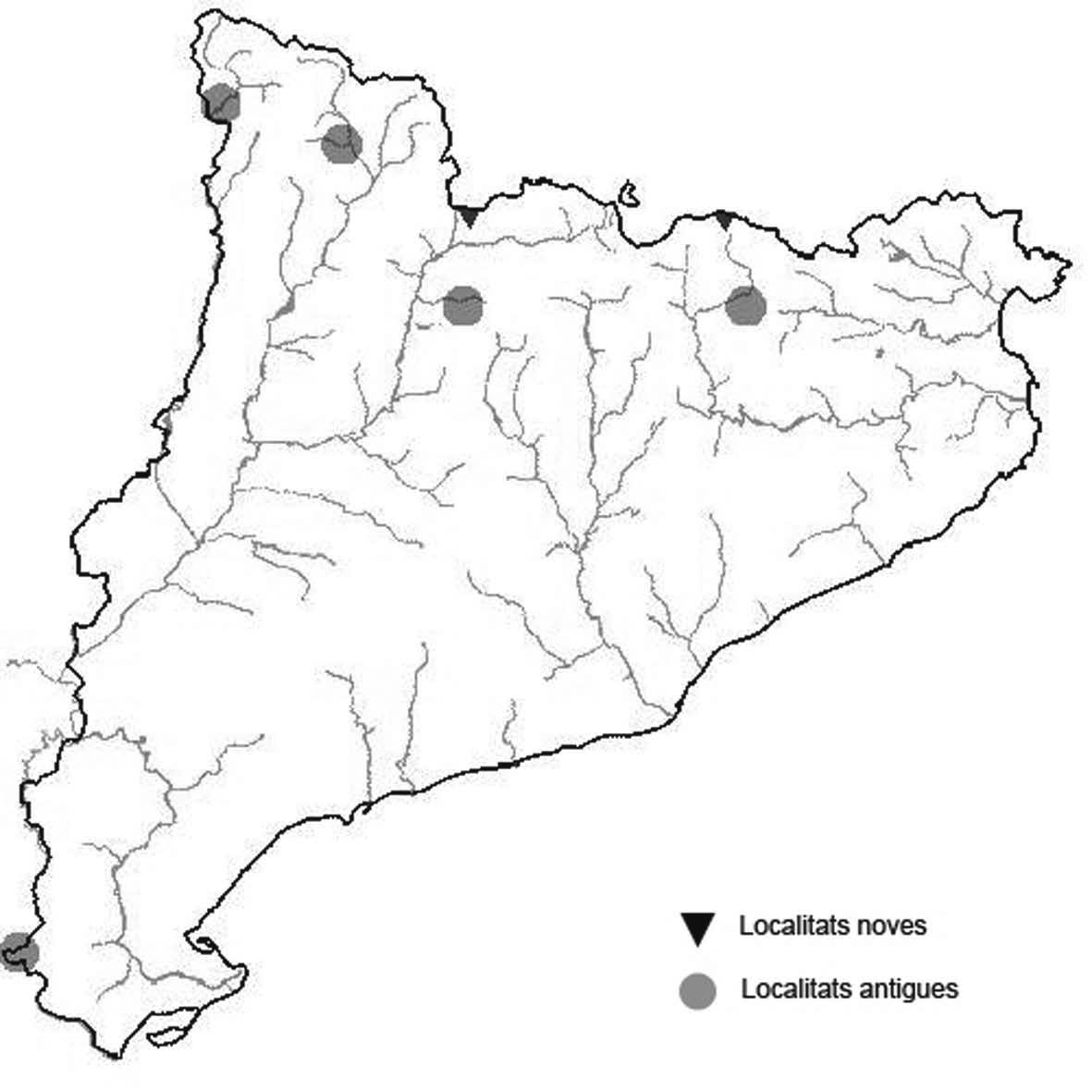
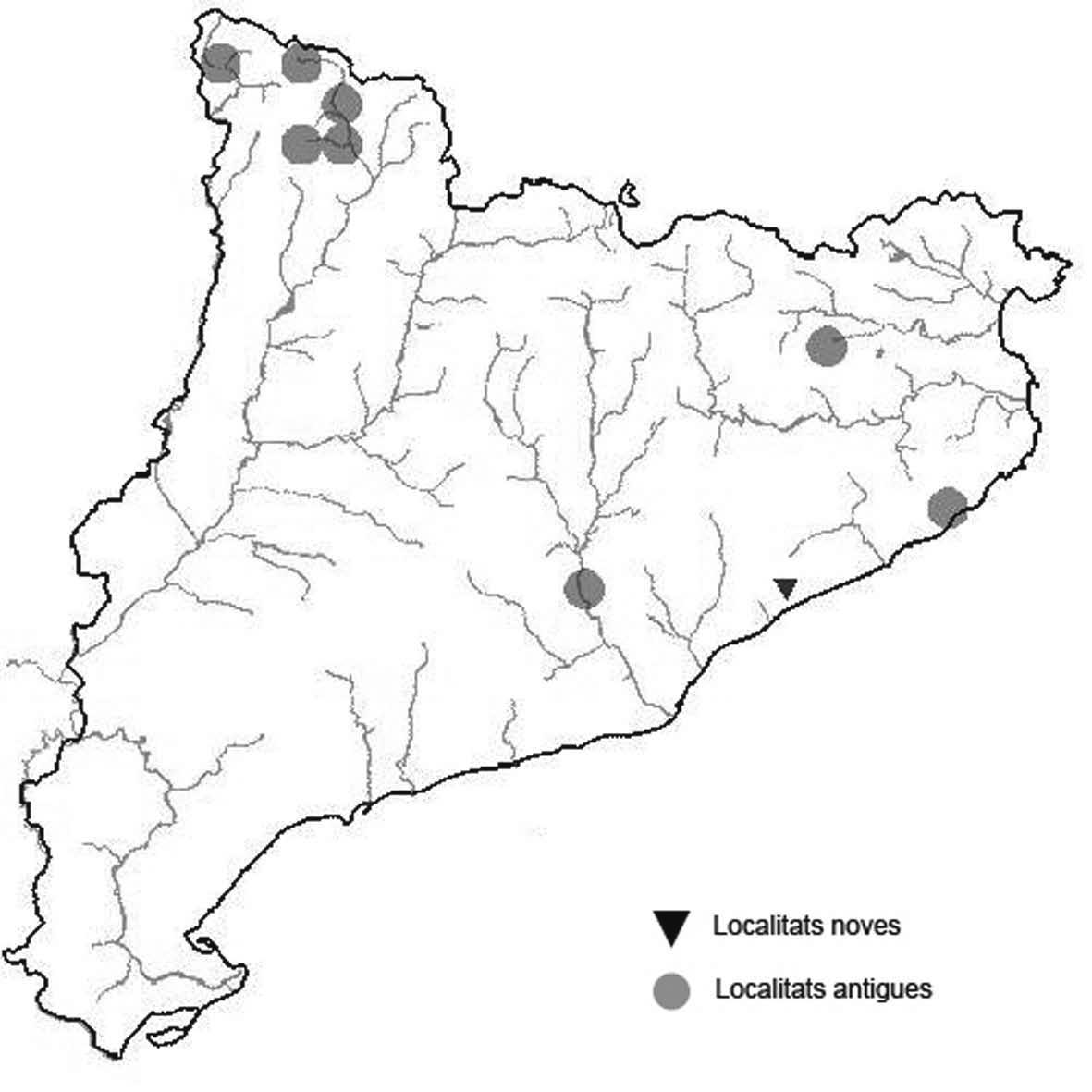
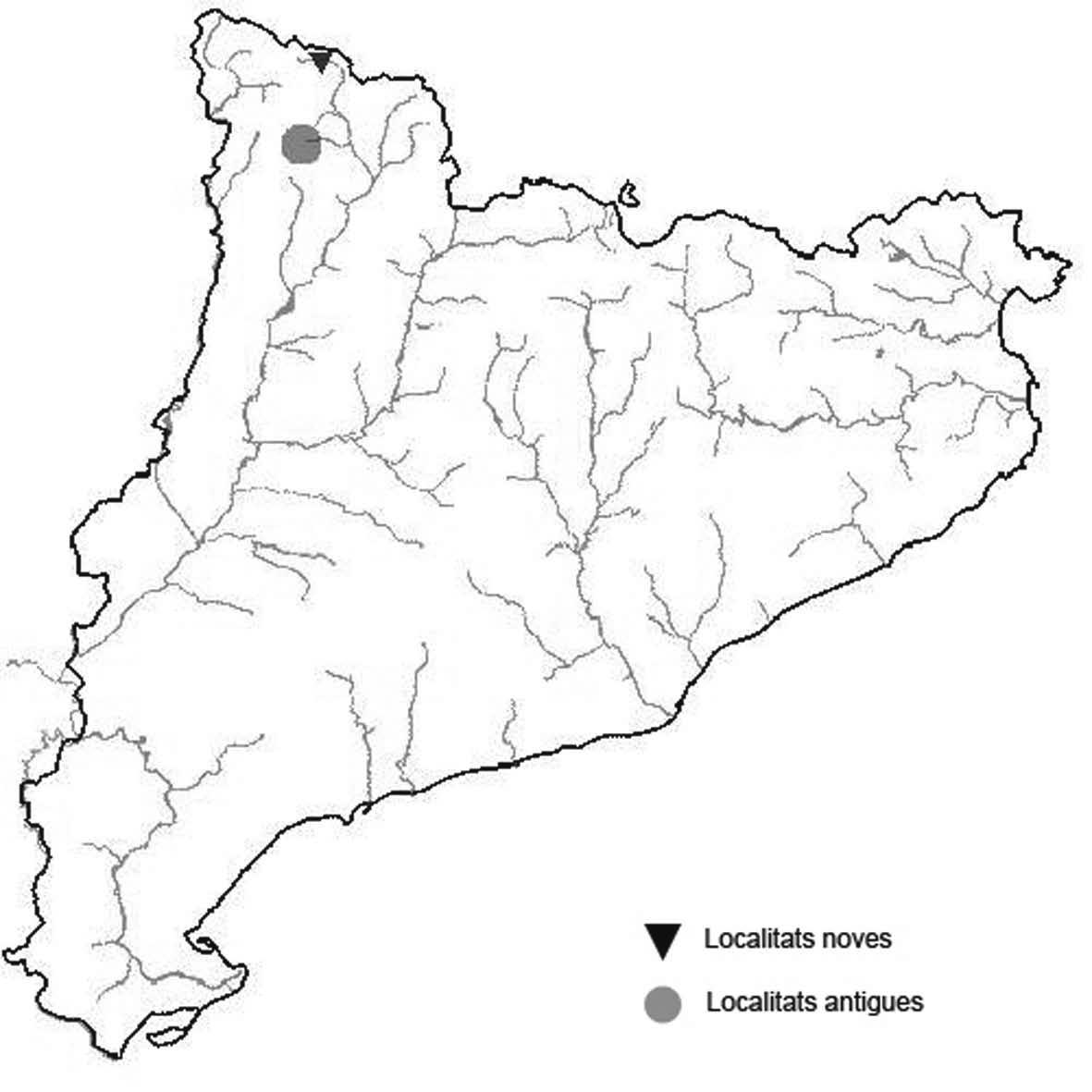
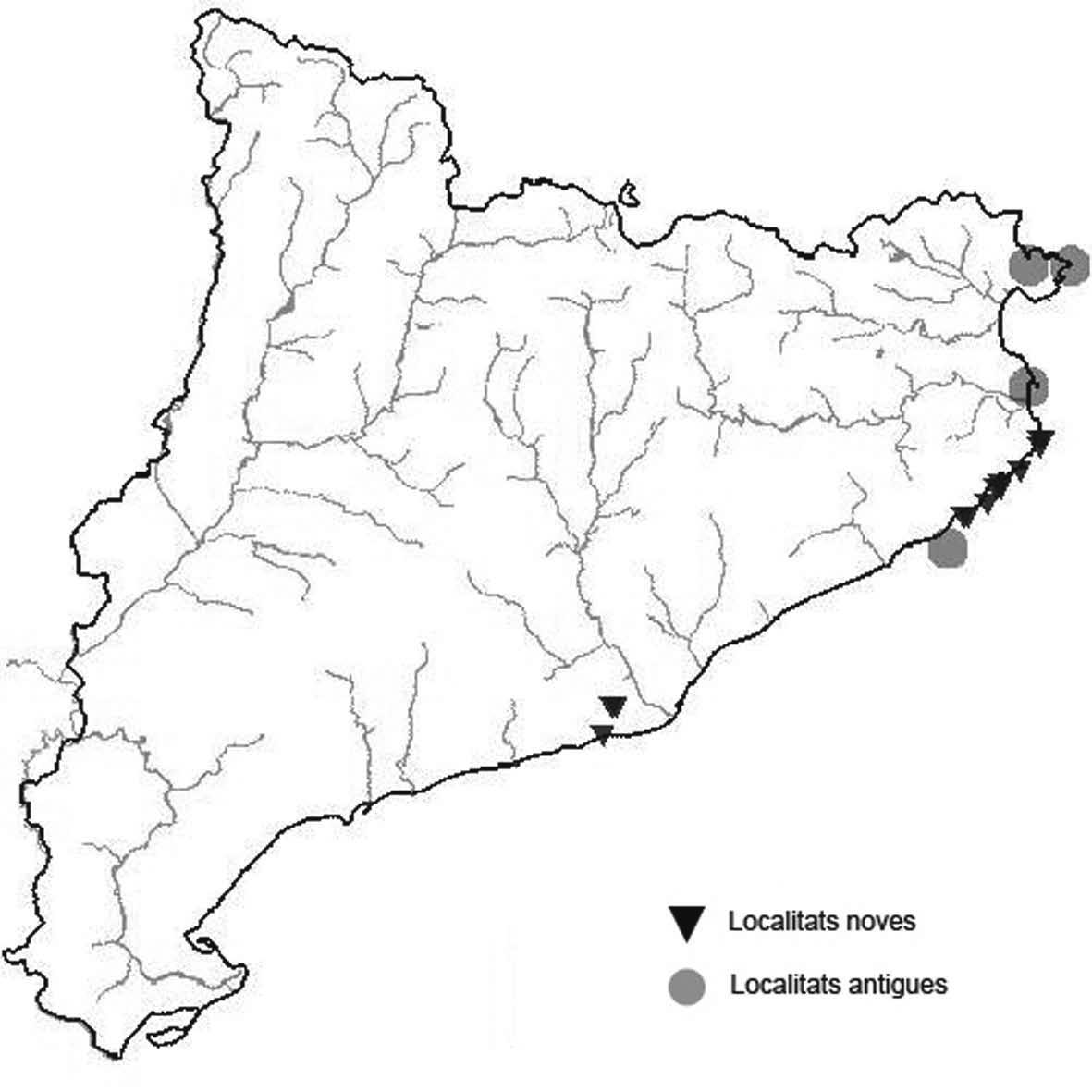
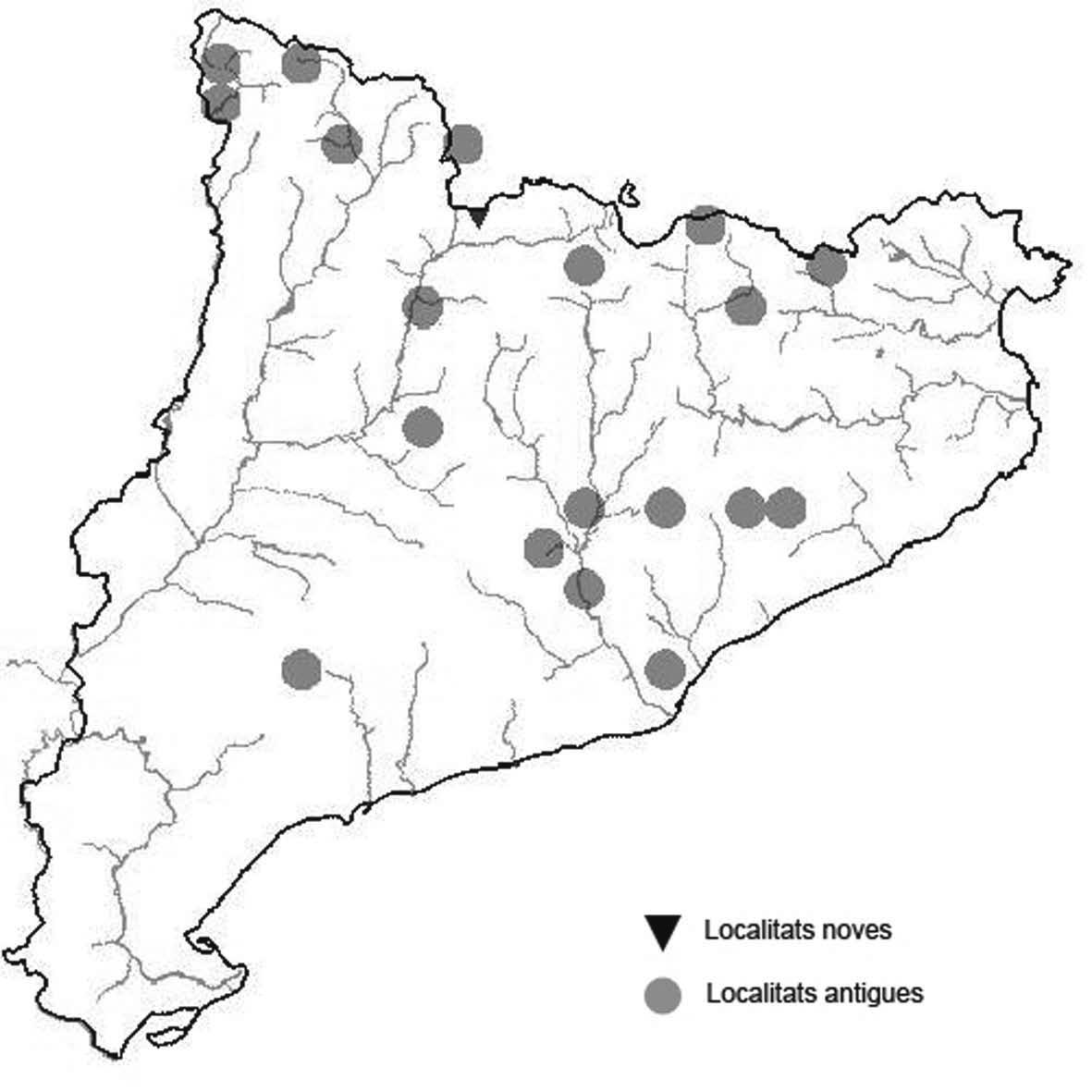
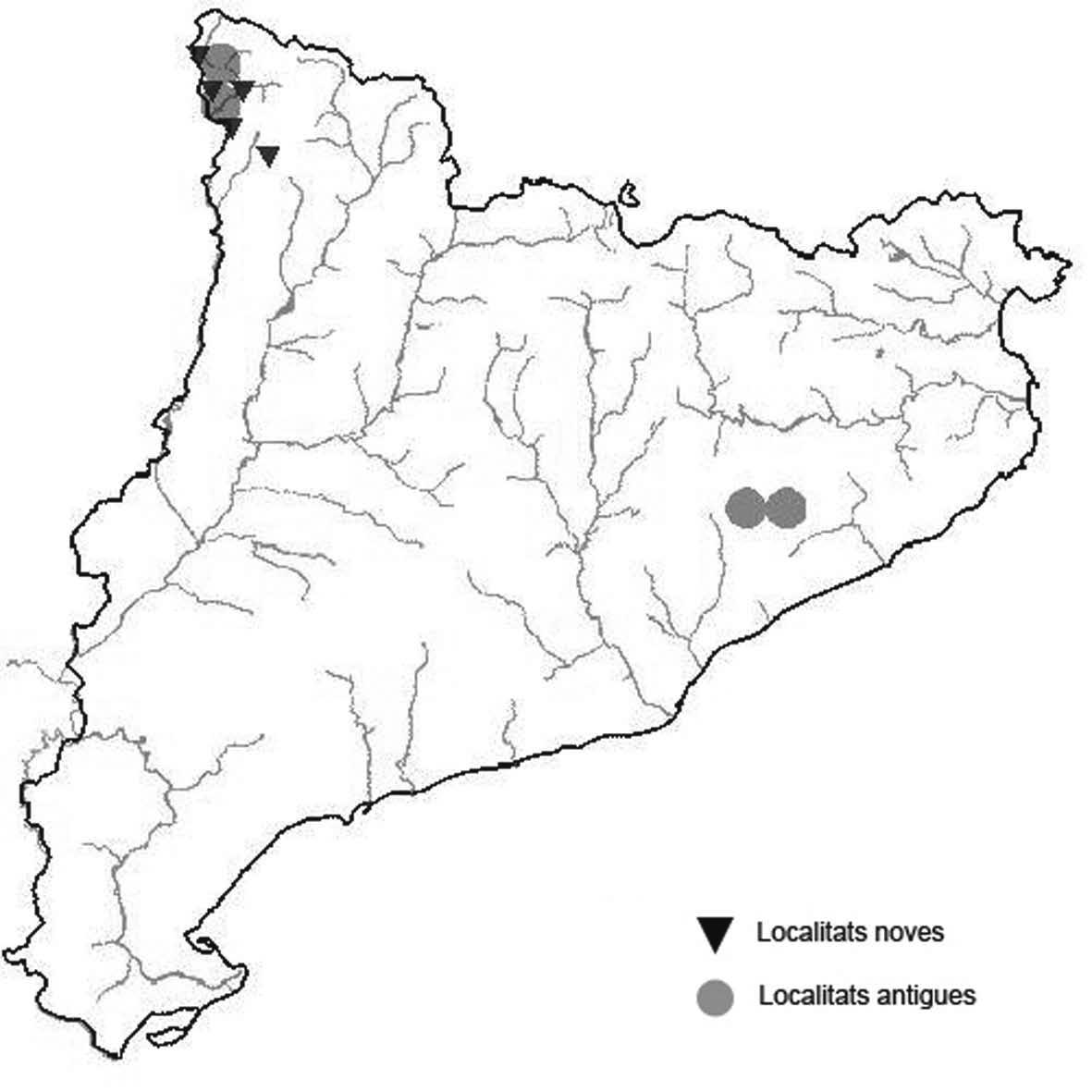
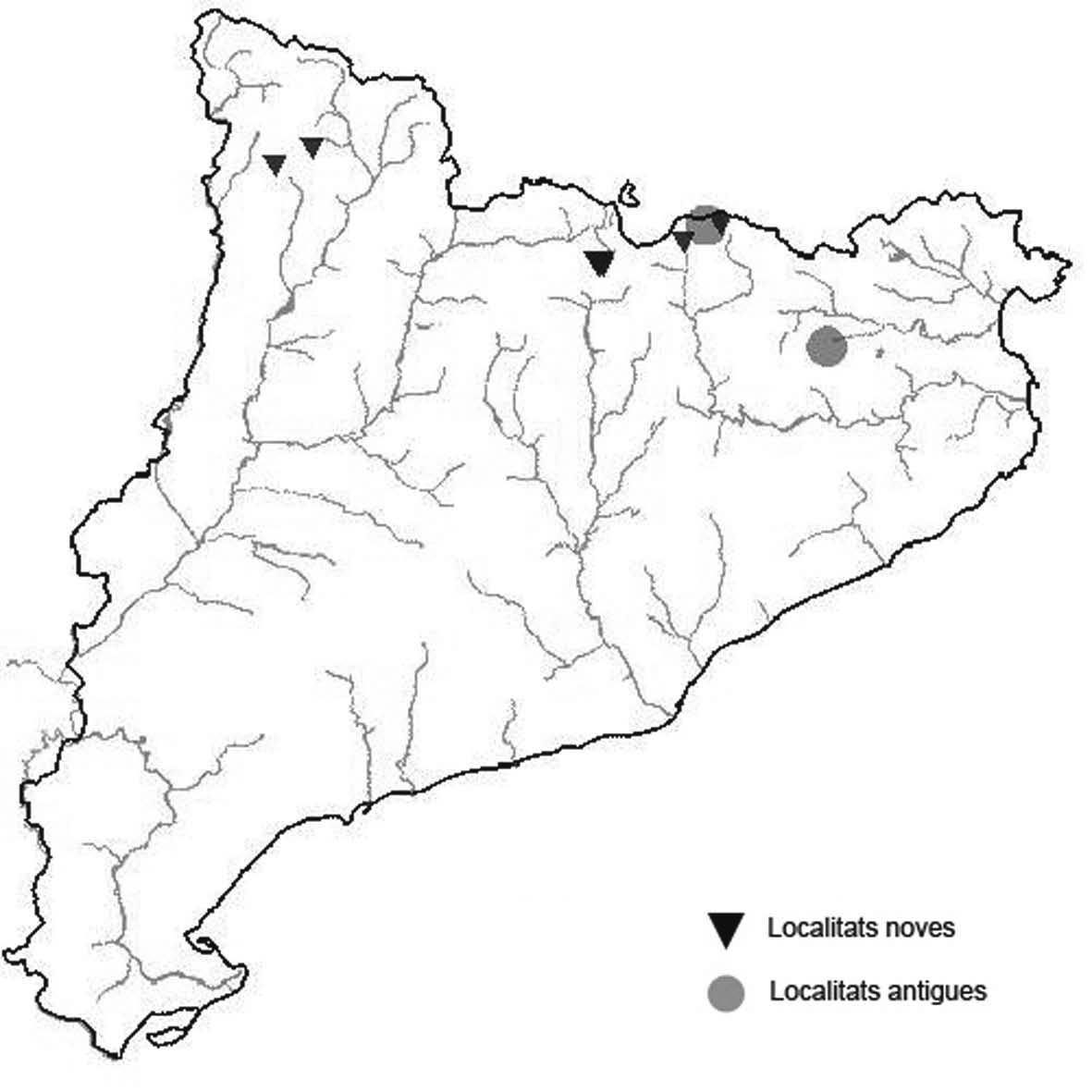
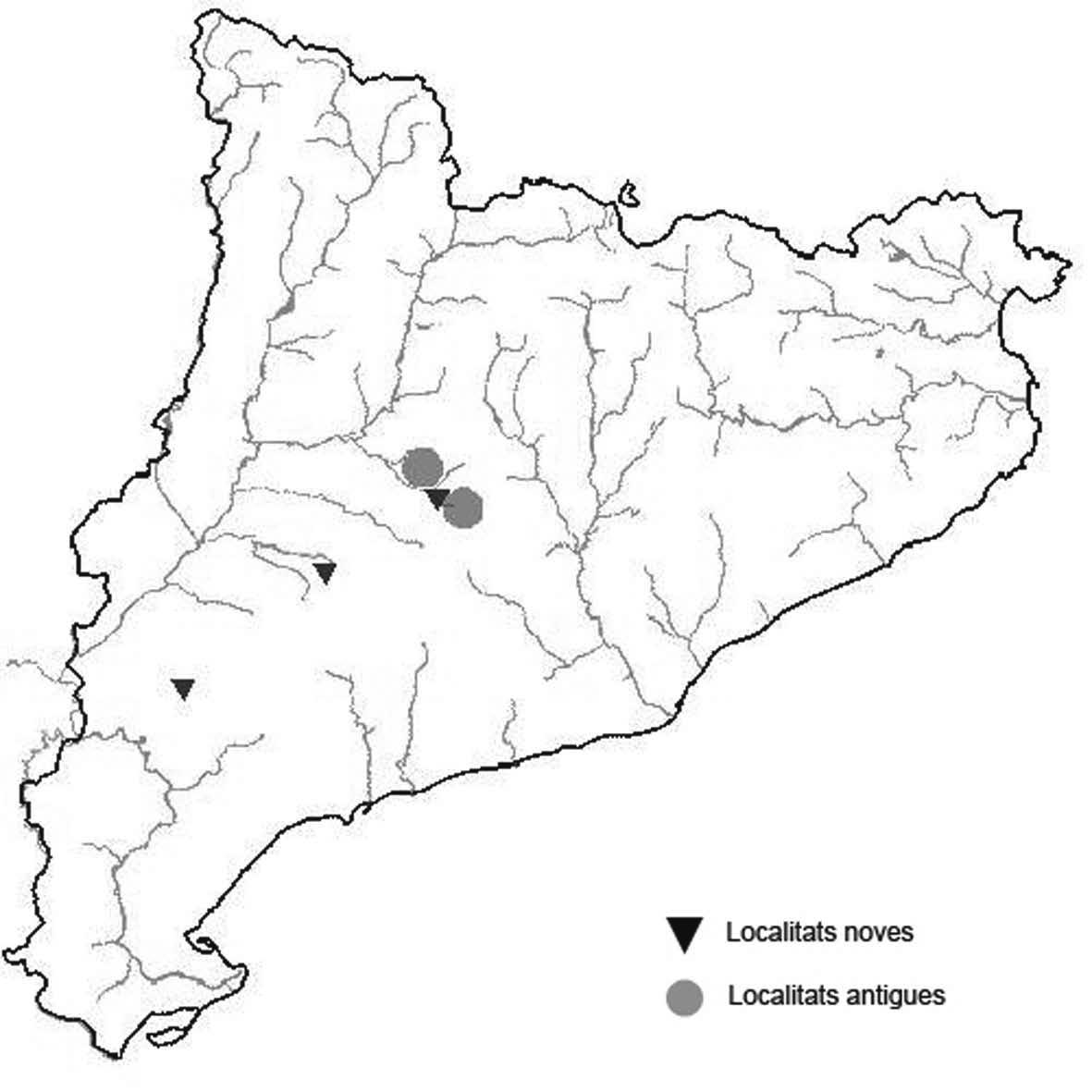
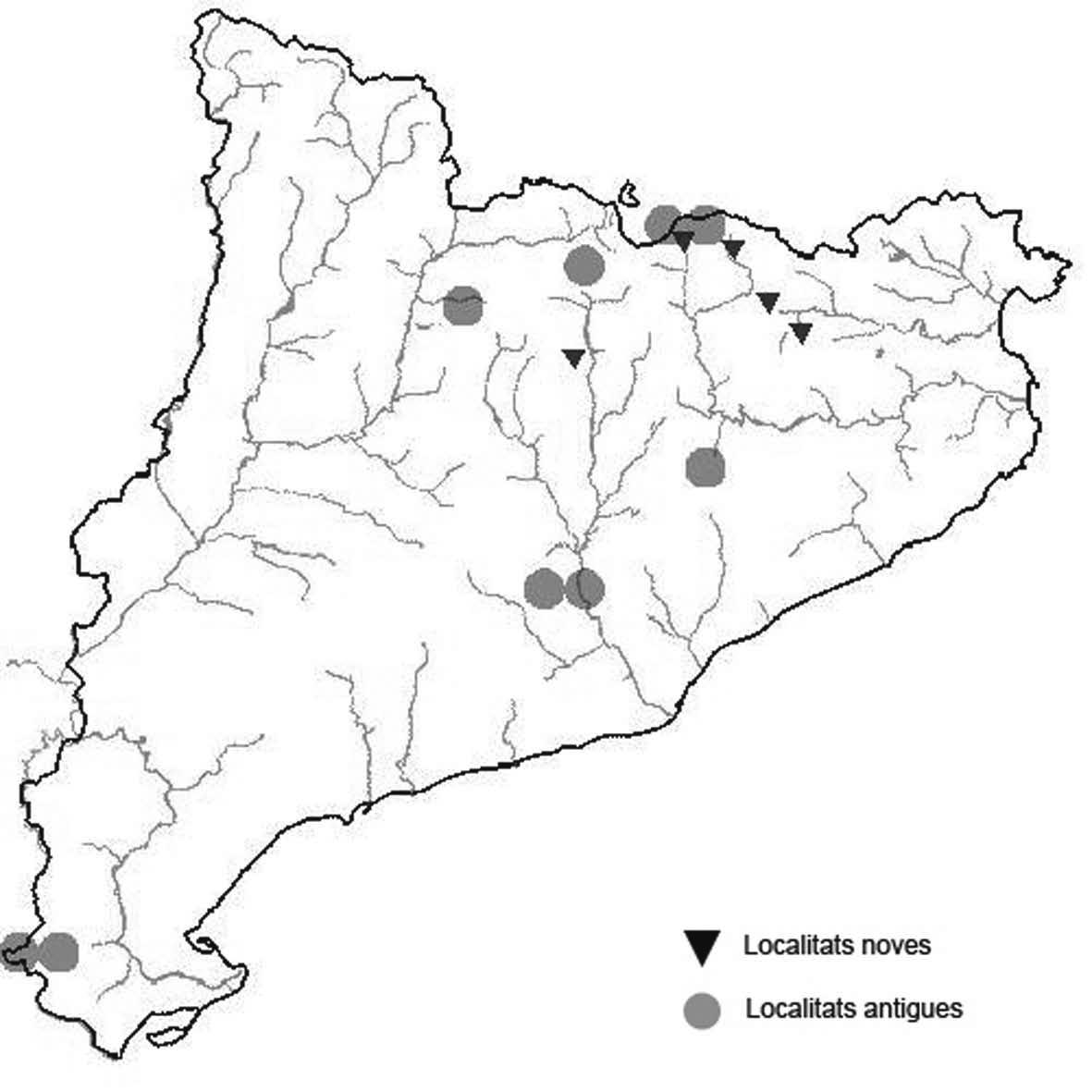
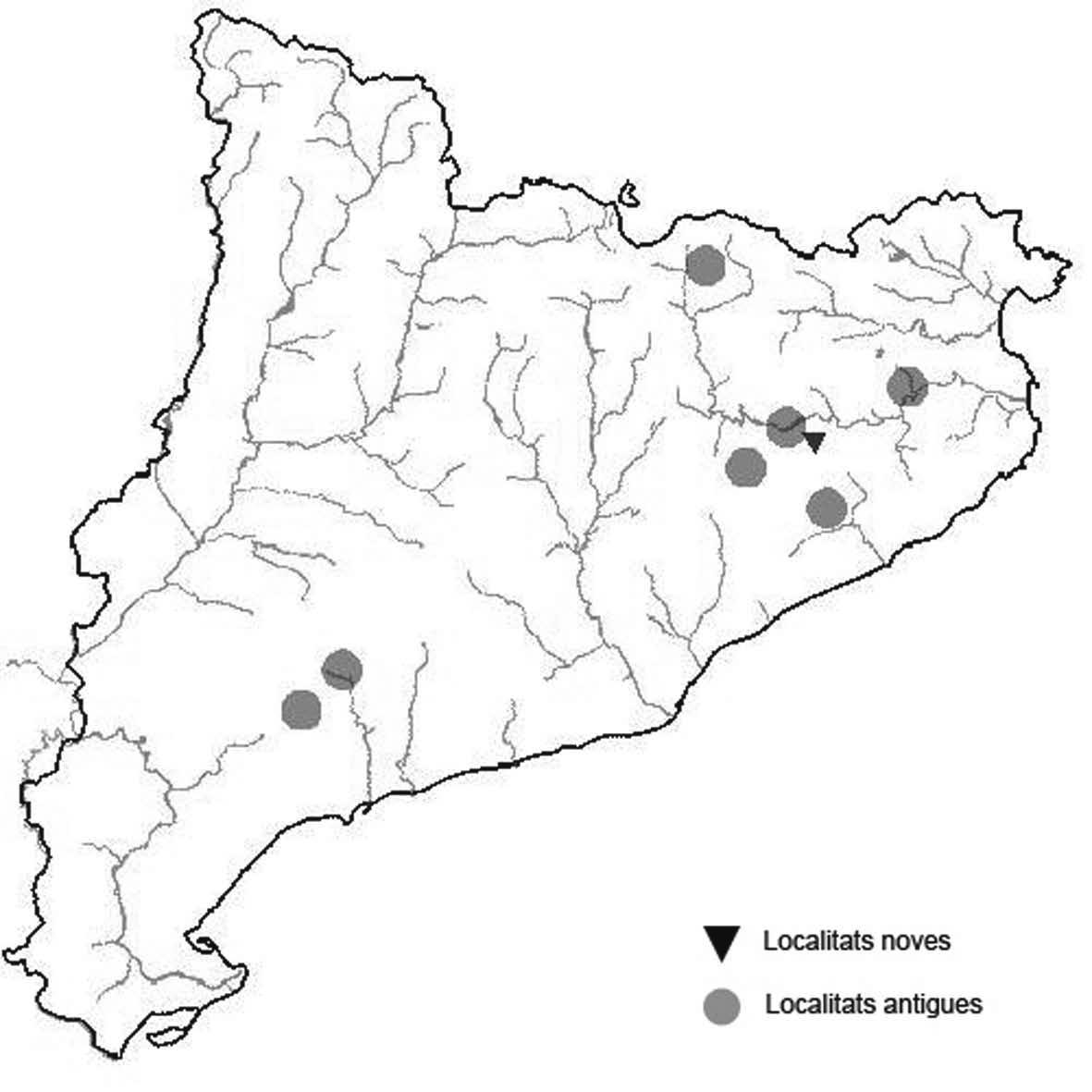
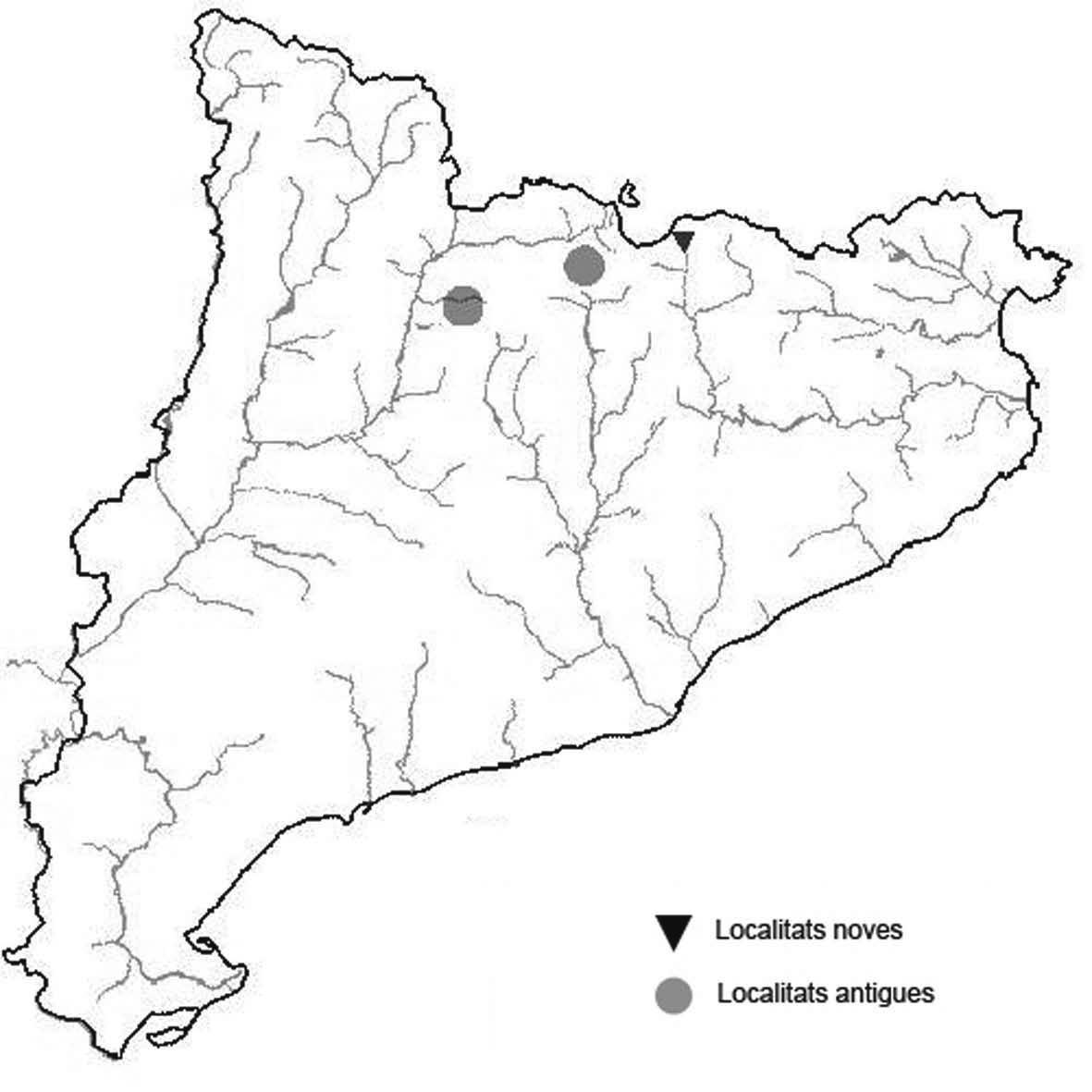
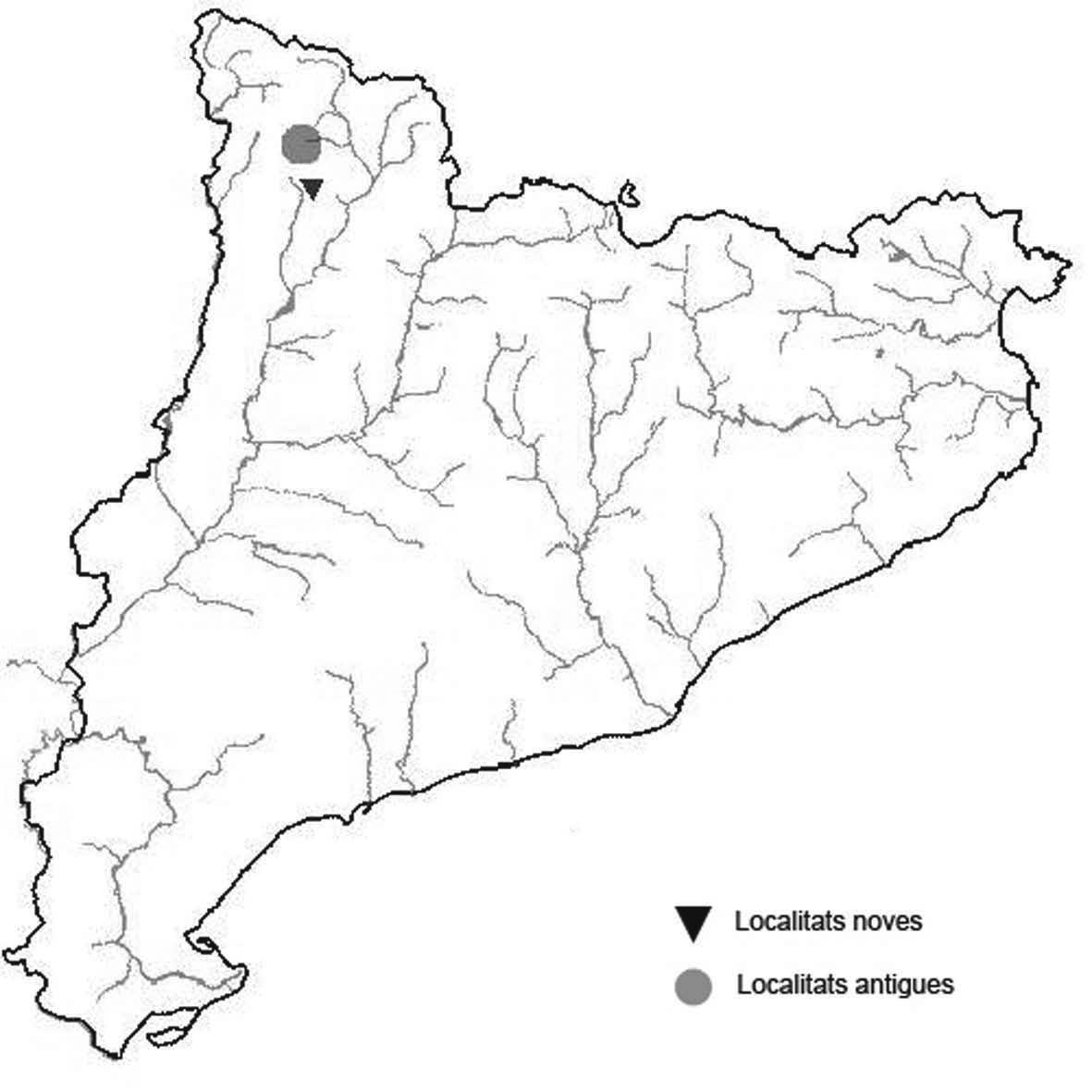
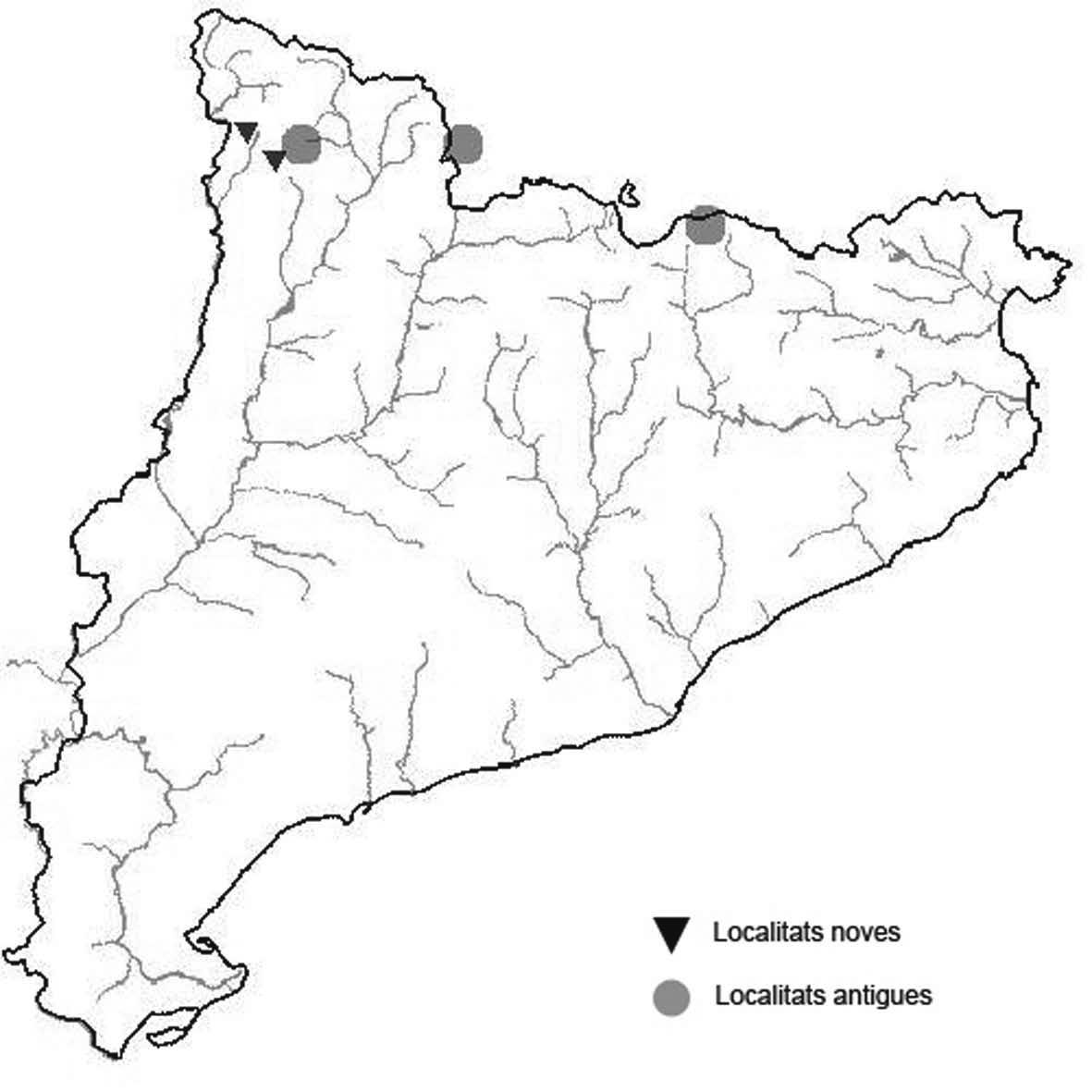
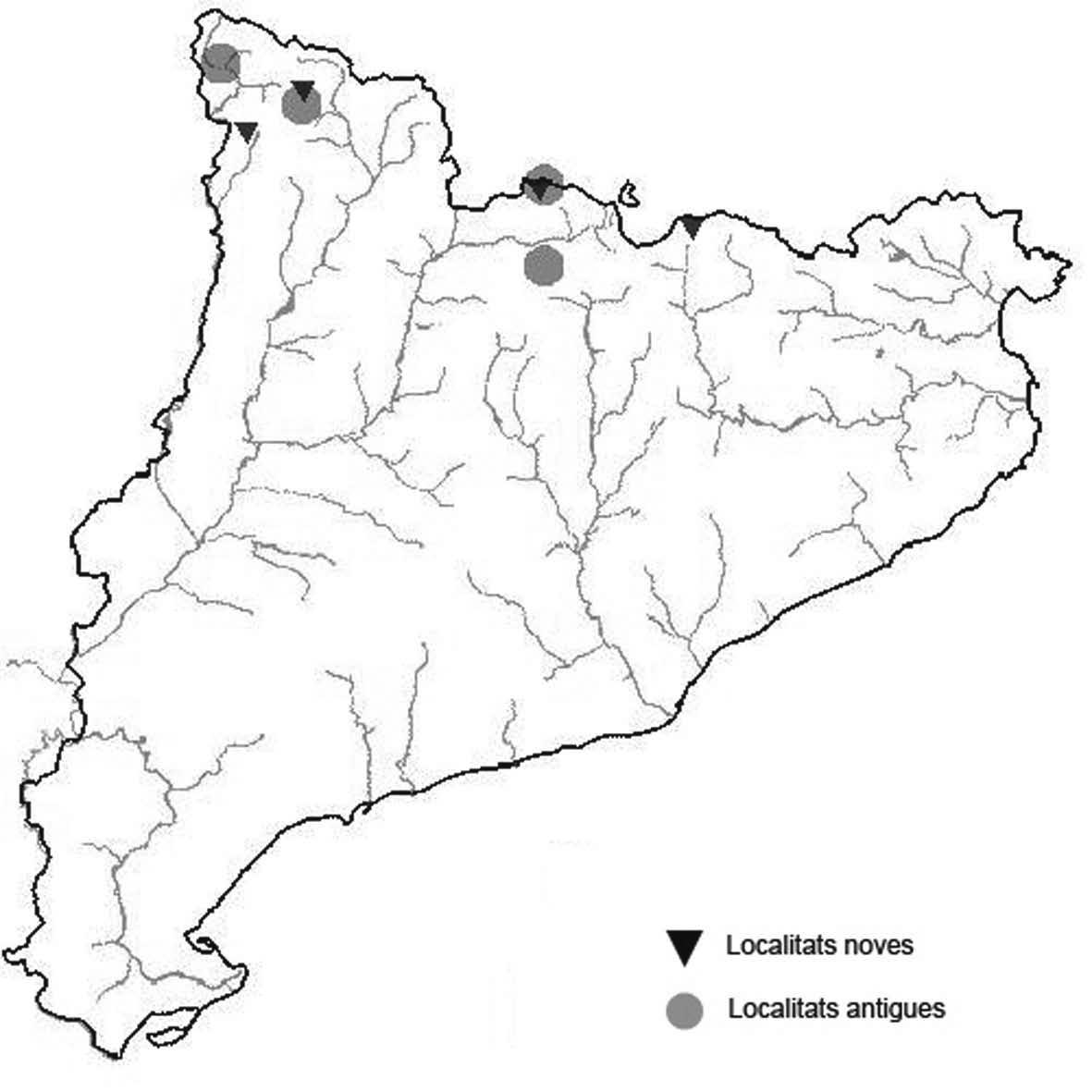
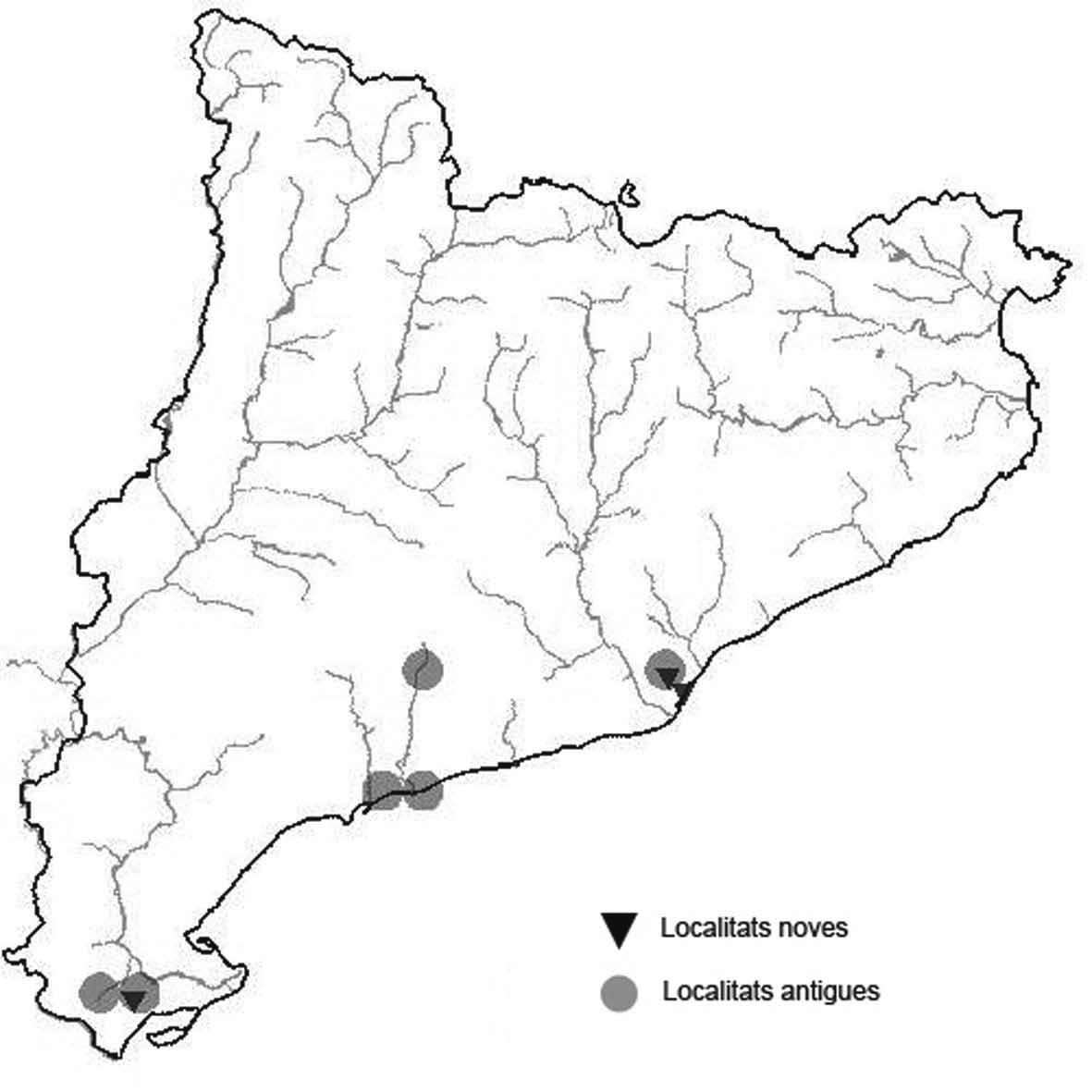
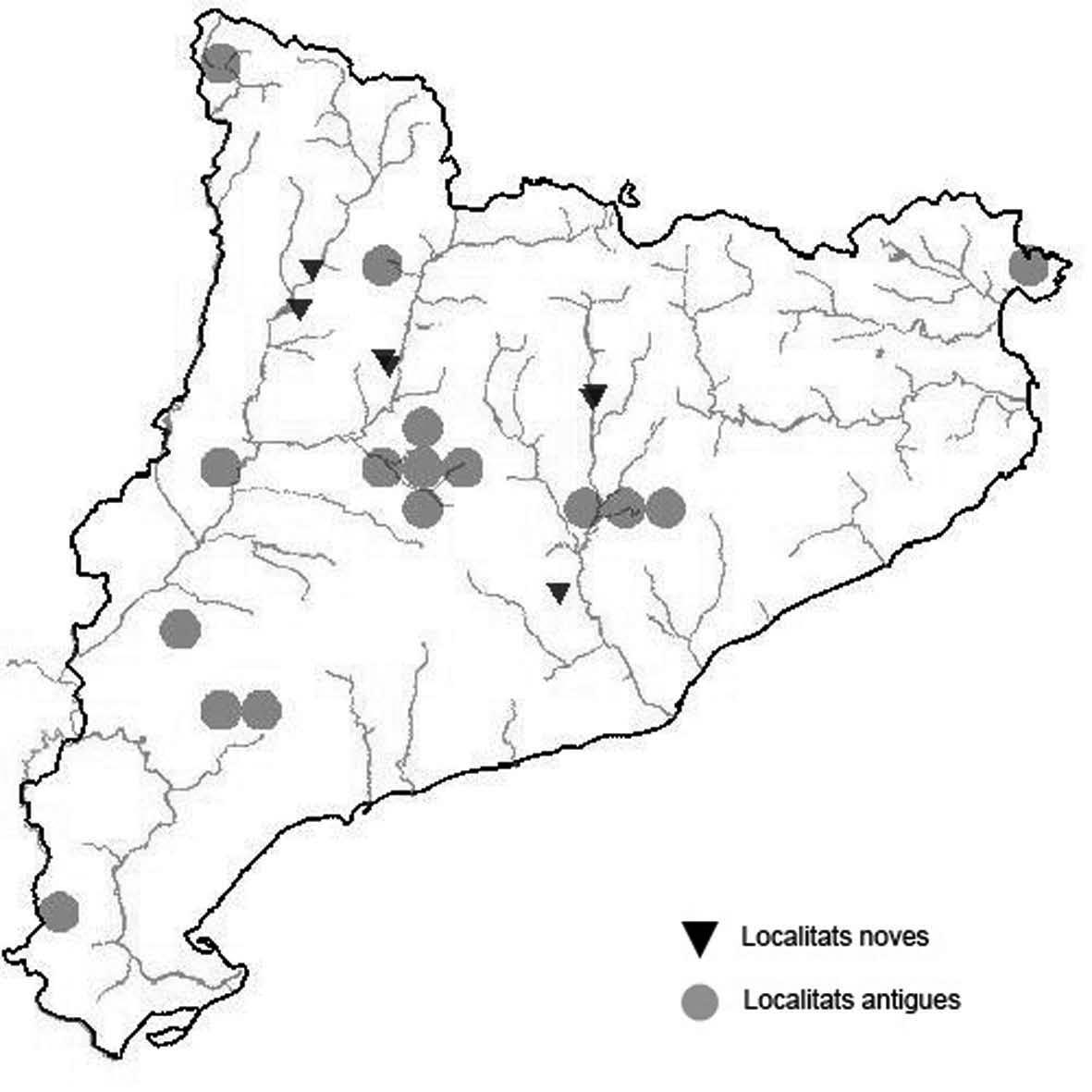
La nomenclatura que seguim és la més actualitzada que tenim per als líquens (http://www.speciesfungorum.org/Names/Names.asp (© 2020 Species Fungorum.), Roux. & coll., 2020 i Nimis & Martellos, 2020). En el cas de canvis nomenclaturals recents s’afegeix també la sinonímia més utilitzada. El llistat de tàxons s’ordena alfabèticament. Per cada tàxon, les localitats s’ordenen alfabèticament pel nom de la comarca, sense l’article, a més es presenta un mapa on representem, de forma diferenciada, les dades anteriors, recollides al Banc de Dades de Biodiversitat de Catalunya (Hladun, 2020), i les aportacions actuals. Amb «**» indiquem la nova cita per Catalunya, i amb «*» marquem els taxons inclosos al catàleg de flora amenaçada.
Resultats
Donem la primera cita per Catalunya de Nephroma tangeriense. Aportem també noves localitats per a 39 tàxons de macrolíquens, en general molt poc citats al nostre país, entre els quals tenim Seirophora lacunosa, considerada com en perill d’extinció a Catalunya, i 16 espècies més que estan incloses com a vulnerables al catàleg de flora amenaçada de Catalunya.
* Acarospora nodulosa (Dufour) Hue var. reagens (Zahlbr.)
Clauzade & Cl. Roux (Fig. 1)
Es tracta d’un líquen terrícola - saxícola que viu sobre guix a terra baixa. Segons Roux & coll. (2020) es tractaria d’un quimiotipus i no d’una varietat.
Anoia, Castellfollit de Riubregós, sobre cal Balaga, crostes liquèniques en timonedes gipsícoles aclarides, 485 m, CG6827, A. Lluent, 1-4-2018. Anoia, Castellfollit de Riubregós, cap Xic, Crostes liquèniques en timonedes gipsícoles, 500-600 m, CG7025, A. Lluent, 26-4-2012. Anoia, Calonge de Segarra, Vinya del Molí, crostes liquèniques en brolles gipsícoles, 495 m, CG7225, A. Lluent, 1-4-2018. Anoia, Calonge de Segarra, Solana de Torremitja, 560 m, CG7426, A. Lluent, 27-2-2019. La Noguera, La Sentiu de Sió, Lo Torricot, brolles sobre guixos, 275 m, CG2129, A. Lluent, 164-2014. La Noguera, Castelló de Farfanya, les Carenilles, crostes liquèniques entremig de pinedes esclarissades, 405 m, CG0933, A. Lluent, 25-6-2016. La Noguera, Camarassa, Valldarnàs, Crostes liquèniques en timonedes gipsícoles, 380 m, CG2536, A. Lluent, 12-6-2016. La Noguera, Camarassa, Tamariu, matollars sobre guixos, 400 m, CG2635, A. Lluent, 12-6-2016. La Segarra, Biosca, camí de lo Coll al cobert del Secà, crostes liquèniques en timonedes gipsícoles, 470 m, CG6431, A. Lluent, 27-2-2019. La Segarra, Sanaüja, costa de les Gesses, crostes liquèniques en timonedes gipsícoles, 450-500 m, CG6035, A. Lluent, 19-1-2013. Segrià, Alfarràs, guixera d’Andaní, crostes liquèniques en timonedes gipsícoles, 350-400 m, BG9735, A. Lluent, 2-7-2012.
Bryoria capillaris (Ach.) Brodo & D. Hawskw. (Fig. 2)
De les tres mostres, BCN-Lich 21886, BCN-Lich 21887 i BCN-Lich 21888, s’ha fet una cromatografia en capa fina
(TLC, abreviatura anglesa) seguint la proposta de Schumm & Elix (2015). En els exemplars BCN-Lich 21886 i BCNLich 21888, l’àcid girofòric era la substància majoritària, però en l’exemplar BCN-Lich 21887 era el barbàtic. Precisament aquesta substància és la que donen com a característica de l’espècie (Velmala et al., 2014). L’exemplar BCN-Lich 21888 té apotecis, fet que segons el mateixos autors, diuen que és molt rar.
Localitat: Pallars Sobirà, Bosc de Marimanha, diversos punts dins del bosc en clarianes, a les branques ben illuminades d’Abies alba, 1680-1820 m, CH3935, leg. A. Lluent. 15-6-2017, det. A. Gómez-Bolea (BCN-Lich 21886, BCN-Lich 21887 i BCN-Lich 21888). En aquesta localitat l’hem trobat acompanyada de Bryoria fuscescens
Bryoria fuscescens (Gyeln.) Brodo & D. Hawksw. (Fig. 3)
De les dues mostres BCN-Lich 21889 i BCN-Lich 21890, s’ha fet una cromatografia en capa fina (TLC) seguint la proposta de Schumm & Elix (2015). En elles hem identificat, com a substància majoritària, l’àcid fumarprotocetràric i traces de protocetràric. Aquest quimiotipus és similar al que Boluda et al. (2015) troben, majoritàriament, a Navacerrada. En un estudi sobre l’ecologia del grup d’espècies de Bryoria entre las que es troba B. fuscescens, a Finlàndia, Myllys et al. (2016) troben que el nostre quimiotipus és especialment abundant a la zona boreal sud.
Localitats: Pallars Sobirà, Lladorre, Pla de Boavi, 1450m, CH6226, sobre Betula pendula, leg. A. Lluent, 2-8-2017, det. A. Gómez-Bolea (BCN-Lich 21889). Pallars Sobirà, Bosc de Marimanha, diversos punts en clarianes, a les branques ben il·luminades d’Abies alba, 1680-1820 m, CH3935, leg. A. Lluent, 15-6-2017, det. A. Gómez-Bolea (BCN-Lich 21890).
Cladonia macrophyllodes Nyl. (Fig. 4)
Anteriorment havia estat citada per Burgaz & Ahti (2009) de la província de Girona, però sense més especificacions i nosaltres la teníem identificada de les muntanyes d’Andorra. Aquesta espècie, que normalment només desenvolupa el tallus primari, l’hem pogut confirmar per la presència de l’atranorina i l’àcid fumarprotocetràric.
Localitats: Alta Ribagorça, Vall de Boí, cresta del Bessiberri Sud, prop del Pas de Trescazes, 2900 m, CH2118, leg. A. Lluent, 24-8-2017, det. A. Gómez-Bolea (BCN-Lich 21892).
* Cladonia mediterranea P.A. Duvign. & Abbayes (Fig. 5)
Liquen terrícola vistós, viu principalment en sols de sauló més o menys argilosos i entre molsa. S’havia recol·lectat per decorar maquetes i pessebres per simular arbres a petita escala.
Localitats: Anoia, sobre el túnel del Bruc, prop de Can solà de la Roca, brolles, 600 m, CG9605, leg. A. Lluent, 11-5-2016, det. A. Gómez-Bolea (BCN-Lich 21893). Baix Empordà, Santa Cristina d’Aro, vora can Dalmau, brolles i boscos clars. 60 m, DG9728, A. Lluent, 20-5-2014. Gironès, Llagostera, carretera de Llagostera a Sant Grau al Km 1, talussos al voltant de la carretera, 180 m, DG9227, A. Llu-
4 Butlletí ICHN 85 (1), 2021 GEA, FLORA ET FAUNA A. LLUENT & A. GÓMEZ-BOLEA


5 Butlletí ICHN 85 (1), 2021
FLORA
FAUNA
MACROLÍQUENS
CATALUNYA AMB NOVES DADES COROLÒGIQUES SOBRE LÍQUENS PROTEGITS
GEA,
ET
DISTRIBUCIÓ DE
A
1 2 3 4 5 6
Figures 1-6. 1) Acarospora nodulosa var. reagens. 2) Bryoria capillaris. 3) Bryoria fuscescens. 4) Cladonia macrophyllodes 5) Cladonia mediterranea. 6) Cladonia mitis
GEA, FLORA ET FAUNA
ent, 18-6-2013. Gironès, Llagostera, Santa Ceclina, brolles de bruc amb sòl molsós, 160 m, DG8827, A. Lluent, 18-62013. Gironès, Llagostera, tossal de Can Bóta, brolles i boscos clars, 150-210 m, DG9434, DG9534 i DG9533, leg. A. Lluent, 15-7-2018, det. A. Gómez-Bolea (BCN-Lich 21894). Maresme, Tordera, vora can Vinyals i bosc de Sant Andreu, pineda esclarissada,. 60-100 m, DG7513, A. Lluent, 143-2018. Selva, Tossa de Mar, vora la resclosa d’Aiguafina, brolles de bruc amb sòl molsós, 70 m, DG9122, A. Lluent, 18-6-2013. Aquesta espècie ja va ser citada d’un lloc proper per Llop & Hladun (2003). Selva, Vidreres, la Goba, pineda esclarissada, 130 m, DG8328, A. Lluent, 18-6-2013. Vallès Occidental, Matadepera, entre la Furriola i la Castellassa del Dalmau, 615 m, DG1911, A. Gómez-Bolea, 27-3-2010.
Cladonia mitis Sandst. (Fig. 6)
D’aspecte semblant a Cladonia arbuscula, es pot diferencia fàcilment per no reaccionar amb el Pd. Alguns autors la consideren només una subespècie de C. arbuscula.
Localitats: Pallars Sobirà, Alt Àneu, estany del Collet de Baciver, carena des de l’estany vers al sud, terrícola en un collet ventat, 2376 m, CH3528, leg. A. Lluent, 8-8-2018, det. A. Gómez-Bolea (BCN-Lich 21895). Pallars Sobirà, Alt Àneu, La Peülla sobre el port de la Bonaigua, en un neretar, 2320 m, CH3423, leg. A. Lluent, 20-9-2018, det. A. Gómez-Bolea (BCN-Lich 21896).
Evernia divaricata (L.) Ach. (Fig. 7)
Localitats: Pallars Sobirà, prop de l’estany de Sant Maurici, sobre Abies alba, 2000 m, CH3516, leg. et det. A. GómezBolea (BCN-Lich 21898). Pallars Sobirà, Naut Aran, Bosc de Marimanha, sobre Abies alba en diversos punts dins del bosc, en clarianes o branques ben iŀluminades, 1680-1820 m, CH3935, leg. A. Lluent, 15-6-2017, det. A. Gómez-Bolea (BCN-Lich 21897).
Glypholecia scabra (Pers.) Müll. Arg. (Fig. 8)
Comparteix hàbitat amb Lecanora dispersoareolata. Confirmem la seva presència a la mateixa localitat d’on havia estat citada 18 anys abans (Hafellner, 1995).
Localitats: Ripollès, Queralbs, Coma de les Mulleres, sota el pic de l’Àliga, roques calcàries verticals, 2250 m, DG3194, leg. A. Lluent, 20-7-2013, det. A. Gómez-Bolea (BCN-Lich 21899).
Lecanora dispersoareolata (Schaer.) Lamy (Fig. 9)
Comparteix hàbitat amb Glypholecia scabra. Constatem la seva presència, 23 anys més tard, a la mateixa localitat en que la van trobar Navarro-Rosinés & Hladun (1990).
Localitats: Ripollès, Queralbs, obaga de coma de les Mulleres, sota el pic de l’Àliga, roques calcàries verticals 2250 m, DG3194, leg. A. Lluent, 20-7-2013, det. A. Gómez-Bolea (BCN-Lich 21900).
* Letharia vulpina (L.) Hue (Fig. 10)
Localitats: Alta Ribagorça, Estany Llong, sobre fusta morta en peu, 2030 m, CH3115, leg. et det. A. Lluent. 22-7-2017,
(BCN-Lich 21901). Cerdanya, Lles de Cerdanya, Serra dels Galls i Bony de l’Os, al costat de la cabana dels caçadors, 2080 m, CG9199, leg. Pere Sala, det. Jaume Arnau, 6-2005, (BCN-Lich 21902). Pallars Sobirà, Alt Àneu, Estany Llong sobre l’estany de Gerber, sobre la part morta d’un pi negre viu, en fusta decorticada, 2320 m, CH3521, G. Giménez, 247-2018. Pallars Sobirà, Espot, bosc de l’obaga dels Estanys de Trescuro, 2055 m, CH4013, leg. et det. E. Carrillo, 20-72004, (BCN-Lich 21903).
* Lethariella intricata (Moris) Krog (Fig. 11)
A Catalunya només s’ha trobat aquesta espècie a la serra de Prades en afloraments del Buntsanstein de la part alta.
Localitats: Baix Camp, Prades, els Pics, en roques exposades i també sota el bosc, ocasionalment sobre els pins, 1080 m, CF3375 i CF3376, C. Casals, 25-11-2016. Baix Camp, Prades, els Colomers, en roques sota el bosc, en aquest punt és escassa, 1050 m, CF3276, C. Casals, 25-11-2016. Baix Camp, Prades, tossal de la Baltassana, en roques prop del cim, 1202 m, CF3276, C. Casals, 1991, població desapareguda probablement per la instaŀlació d’una placa commemorativa. Conca de Barberà, Vimbodí, bosc de can Felip, obaga de la Moleta, a banda i banda de la pista, en roques exposades i també sota el bosc, ocasionalment sobre els pins, 1160 m, CF3377, C. Casals, 25-11-2016. Conca de Barberà, Vimbodí, coll de Cova Fumada, aflorament de roques al mig del bosc, ocasionalment sobre els pins, 1160 m, CF3776, A. Lluent, 25-11-2016. Conca de Barberà, Vimbodí, Mola d’Estat, al cingle que mira al nord, 1125 m, CF3776, A. Lluent, 25-112016. Conca de Barberà, Vimbodí, entre la Mola d’Estat i la Mola dels Quatre Termes, al cingle que mira a l’oest, 1115 m, CF3876, A. Lluent, 25-11-2016.
* Lobaria pulmonaria (L.) Hoffm. (Fig. 12)
Citada per Llenas (1909) com una espècie «comuna a les roques, els arbres i per terra del Montseny, l’Empalme, Celrà, voltants de Girona, Vimbodí, boscos del Monestir de Poblet, etc.», actualment no podem dir que es tracti d’una espècie comuna.
Localitats: Conca de Barberà, Muntanyes de Prades, en aquesta localitat havia estat citada de manera poc precisa (Hladun i Gómez-Bolea, 1984), hem trobat diverses localitats a la obaga del massís entre els 900 i 1200 m, Tossal Gros, sobre Quercus pubescens, un únic taŀlus viu i un de mort, 1080 m, CF3379, A. Lluent, 24-4-2015. Conca de Barberà, Muntanyes de Prades, La Baltassana, talús de la pista que puja al cim, sobre roques, 3 tal·lus grans molt propers, 1175 m, CF3376, A. Lluent, 1-6-2015. Conca de Barberà, Muntanyes de Prades, Vall de Castellfollit, serra de les Boixeres, sota la carena però amb poc pendent, sobre Q. pyrenaica, és la localitat on l’hem vist més abundant hi hem trobat unes desenes de tal·lus dispersos però especialment concentrats en 2 roures, 1130 m, CF3577, A. Lluent, 14-5-2015. Conca de Barberà, Muntanyes de Prades, Vall de Castellfollit, sota el coll d’en Perroi, sobre Q. pyrenaica, 2 tal·lus petits, 1050 m, CF3676, A. Lluent, 29-4-2015. Conca de Barberà, Muntanyes de Prades, Vall de Castellfollit, pista al Titllar a l’alçada
6 Butlletí ICHN 85 (1), 2021
A. LLUENT & A. GÓMEZ-BOLEA

7 Butlletí ICHN 85 (1), 2021 GEA, FLORA ET FAUNA
DE MACROLÍQUENS A CATALUNYA AMB NOVES DADES COROLÒGIQUES SOBRE LÍQUENS PROTEGITS 7 8 9 10 11 12
Figures 7-12. 7) Evernia divaricata; 8) Glypholecia scabra; 9) Lecanora dispersoareolata; 10) Letharia vulpina. 11) Lethariella intricata; 12) Lobaria pulmonaria
DISTRIBUCIÓ
GEA, FLORA ET FAUNA
de la ruta micològica, sobre Q. pyrenaica,. únicament 2 tallus, 910 m, CF3677, A. Lluent, 1-6-2015. La Garrotxa, Vall de Bianya, Baga de la riera de Farró, el Pujolet, el Farró, sobre Q. robur, entre 439 i 446 m, DG5175 (Zapata et al. 2016). Maresme, Canyamars, camí del Pou de Glaç, sobre Q. cerrioides i Q. ilex. 220 m, DG5405, A. Gómez, 2018. Osona, Viladrau, Espinzella, sobre roures i una alzina, 723 m, DG4533, Dolors Rodríguez, 2018. El Ripollès, Milany, sobre Fagus sylvatica, Fraxinus excelsior i Crataegus monogyna, entre 1499 i 1519 m, DG4168, (Zapata et al. 2016). Arbúcies, turó de Montfort, sobre roques amb molses, 670 m, DG5926, D. Rodríguez. Selva, Osor, Sant Miquel de les Formigues, sobre roques amb molses, 1140 m, DG6141, Francesc Canaleta, 3-6-2020. Val d’Aran. Malgrat ser una espècie força freqüent pràcticament no s’ha citat en aquesta comarca. Contribuïm aquí amb algunes localitats per anar completant la corologia d’aquesta espècie a la comarca aranesa. Val d’Aran, Vall de Canejan, sota el Barratge deth Hons dera Coma 1180 m, CH2242, sota el nucli de Sant Joan de Toran, entre la carretera i el riu, 955 m, CH1943, Es Borcalòs, 790 m, CH1744. A. Lluent. 20-102017, freqüent a tota la vall, sobre faig, vern, freixe i altres caducifolis. Val d’Aran, Vall de Varradós, es Prats Vielhs, 1130 m, CH1736. Es Comaretes, entre la carretera i el riu de Varradós, sobre faig principalment, molt abundant tant en els troncs com a les branques principals, 1495 m, CH2038, A. Lluent, 19-10-2017. Val d’Aran, Eth Portillon, dins del recinte de l’Aran Park, sobre faig, til·ler i altres caducifolis i també avet, 1175 m, CH0937, A. Lluent, 21-10-2017. Val d’Aran, Artiga de Lin, als Uelhs deth Joeu, freqüent a tota la vall tan en avets com en faigs on és més abundant, 1395 m, CH1227, CH1228, CH1229. A. Lluent. 21-10-2017. En l’Artiga de Lin ja havia estat citada de per Azuaga & Gómez-Bolea (1996).
Lobarina scrobiculata (Scop.) Nyl. (Fig. 13)
Citada per primera vegada per Llenas (1909) del Montseny, Berga i els boscos del Monestir de Poblet.
Localitats: Alta Ribagorça, obaga de l’estany Llebreta, sobre roques amb molses en un bedollar, 1689 m, CH2612, leg. A. Lluent. 7-9-2017, det. A. Gómez-Bolea (BCN-Lich 21904). Val d’Aran, Eth Portillon, dins del recinte de l’Aran Park, sobre faig, 1120 m, CH0937, A. Lluent, 21-10-2017, det. A. Gómez-Bolea (BCN-Lich 21905). Val d’Aran, Vall de Canejan, sota el Barratge deth Hons dera Coma, 1180 m, CH2242, A. Lluent, 20-10-2017.
* Nephroma parile (Ach.) Ach. (Fig. 14)
Localitats: Ripollès, Vilallonga de Ter, Ribera de Catllar, torrent dels Emprius, lutites negres, fil·lites i pissarres, en un bosc de ribera mixt, sobre Salix caprea, 1450 m, DG3990, (E. Llop, comunicació personal). Selva, vessant NE de les Agudes (Montseny), sobre Fagus sylvatica, 1500 m, DG5326, leg. A. Lluent, 10-4-2012, det. A. Gómez-Bolea (BCN-Lich 21906).
Nephroma resupinatum (L.) Ach. (Fig. 15)
Localitats: Alta Ribagorça, Vall de Boí, cresta de Bessiberris, 2900 m, CH2118, leg. A. Lluent, 24-8-2017, det. A.
Gómez-Bolea (BCN-Lich 21907). Alta Ribagorça, obaga de l’Estany Llebreta, sobre roca molsosa en un bedollar, 1689 m, CH2612, leg. A. Lluent, 7-9-2017, det. A. Gómez-Bolea (BCN-Lich 21908).
** Nephroma tangeriense (Maheu & A. Gillet) Zahlbr. (Fig. 16)
Segons les nostres dades, es tracta de la primera cita per Catalunya. La localitat més propera a la nostra és la del Barranco de la Mosquera, en la Sierra de Espadán, (Castelló) (Burgaz & Martínez, 1999). Burgaz & Martínez (2003), el consideren un element macaronèsic i de l’occident europeu, que presenta una distribució suroccidental a la Península ibèrica.
Localitats: Maresme, Dosrius, Can Canyamars, camí de Can Cames, en un talús de roca granítica molt alterada, orientat al nord, acompanyat per Peltigera polydactylon, 220 m, DG5405, leg. i det. A. Gómez-Bolea, 7-10-2018, (BCN-Lich 21909).
* Nephromopsis cucullata (Bellardi) Divakar, A. Crespo & Lumbsch (= Flavocetraria cucullata (Bellardi) Kärnefelt & A. Thell) (Fig. 17)
Localitats: Alta Ribagorça, Muntanyó de Llacs sobre la Font del Graller, prat rocós en fort pendent orientat al nord, entremig de molses, acompanyat de Thamnolia vermicularis, 2225 m, CH2811, leg. A. Lluent, 21-8-2018, det. A. GómezBolea (BCN-Lich 21910).
Nephromopsis nivalis (L.) Divakar, A. Crespo & Lumbsch (= Flavocetraria nivalis (L.) Kärnefelt & A. Thell) (Fig. 18)
Localitats: Alta Ribagorça, Vall de Boí, cresta de Bessiberris prop del pas de Trescazes, Terrícola entre les roques, 2910 m, CH2118, leg. A. Lluent, 24-8-2017, det. A. Gómez-Bolea (BCN-Lich 21912). Pallars Sobirà, Alt Àneu, Estany del Collet de Baciver, carena des de l’estany vers al sud, terrícola en un collet ventat, 2376 m, CH3528, leg. A. Lluent, 8-8-2018, det. A. Gómez-Bolea (BCN-Lich 21911).
* Parmotrema hypoleucinum (J. Steiner) Hale (Fig. 19)
Localitats: Baix Empordà, Palamós, vora la riera de l’Aubi, en bosc mixt de suros i pins, 15 m, EG1235, A. Lluent, 14-7-2013. Maresme, Can Canyamars, prop de Can Cames, epífit sobre Quercus ilex, 250 m, DG5405, A. Gómez-Bolea, 7-10-2018. Maresme, Tordera, can Vinyals, en bosc mixt d’alzines, suros i roures, 60 m, DG7514, A. Lluent, 17-32017. Lloret de Mar, coll del Cantó, en bosc mixt de d’alzines, suros i pins, 150 m, DG8820, A. Lluent, 22-5-2013. Tossa de Mar, capçalera del torrent de Vallpresona i voltants del castell de Montagut, en bosc mixt d’alzines, suros i pins, 350-450 m, DG9523, A. Lluent, 22-6-2013. Selva, Tossa de Mar, vora can Seca, en bosc mixt d’alzines, suros i pins, 35 m, DG9219, A. Lluent, 22-5-2013. Selva, Tossa de Mar, Al costat de la resclosa d’Aiguafina, en bosc mixt d’alzines, suros i pins, 35 m, DG9122, A. Lluent, 22-5-2013.
8 Butlletí ICHN 85 (1), 2021
A. LLUENT & A. GÓMEZ-BOLEA
9 Butlletí ICHN 85 (1), 2021 GEA, FLORA ET FAUNA
DISTRIBUCIÓ DE MACROLÍQUENS A CATALUNYA AMB NOVES DADES COROLÒGIQUES SOBRE LÍQUENS PROTEGITS 13 14 15 16 17 18
Figures 13-18. 13) Lobarina scrobiculata; 14) Nephroma parile. 15) Nephroma resupinatum; 16) Nephroma tangeriense; 17) Nephromopsis cucullata; 18) Nephromopsis nivalis.
GEA, FLORA ET FAUNA
* Peltigera aphthosa (L.) Willd. (Fig. 20)
Espècie tipus de la secció Peltidea (Miadlikowska & Lutzoni, 2000). Morfològicament es pot confondre amb P. britannica, que també presenta cefalodis. Ambdues espècies tenen a la cara superior un fi toment erecte a l’extrem final del lòbuls i la cara inferior sense venes ben definides. La forma dels cefalodis s’ha fet servir com a criteri per separa-les (Burgaz & Martínez, 2003). Donada la variabilitat en els tipus de cefalodis entre P. aphthosa i P. britannica, i les recents dades moleculars que qüestionen l’estatus filogenètic d’espècie per P. britannica (Miadlikowska et al. 2018), podria ser millor parlar de P. aphthosa en sentit ampli, incloent P. britannica. Llenas (1905), referint-se a Catalunya, cita textualment «Crece muy abundante en el suelo de los bosques musgosos de casi todos los montes» i continua citant tota una sèrie de localitats «Montserrat, Moyà, Arbucias, Viladrau Santa Fé, Celrá, Cadaqués, Ribas, Núria Campeyas, Berga, Olot, bosques del Monasterio de Poblet, etc.» I a més recull cites d’altres botànics. Aquesta observació de principis del segle XX no es correspon amb la realitat que tenim representada en el mapa de distribució.
Localitats: Alta Ribagorça, Estany Llong, a pocs metres de la riba sud, sobre roques amb molses a la mitja l’ombra de nerets, 2005 m, CH3115, leg. A. Lluent, 22-7-2017, det. A. Gómez-Bolea (BCN-Lich 21917). Alta Ribagorça, Muntanyó de Llacs, sobre roques amb molsa cobertes parcialment per nerets, 2125 m, CH2811, leg. A. Lluent, 21-8-2018, det. A. Gómez-Bolea (BCN-Lich 21916). Andorra, La Rabassa, talussos de pista forestal obaga en una pineda de pi negre esclarissada, 1980 m, CG7999, A. Lluent, 10-8-2014. Pallars Sobirà, Espot, estanys de Trescuro, al marge sud de l’estany de baix, sobre roques amb molsa dins de la pineda, 2045 m, CH4012, leg. A. Lluent, 21-9-2018, det. A. Gómez-Bolea (BCN-Lich 21915). Pallars Sobirà, estació d’esquí de Baqueira Beret, La Peülla, 2220 m, CH3524, leg. A. Lluent, 2009-2018, det. A. Gómez-Bolea (BCN-Lich 21914). Pallars Sobirà, Saboredo, 2100 m, CH3321, leg. A. Lluent, 20-092018, det. A. Gómez-Bolea (BCN-Lich 21913).
Peltigera collina (Ach.) Schrad. (Fig. 21)
Sota el nom de Peltigera limbata Delise, Llenas (1905, 1909) fa un comentari sobre la distribució d’aquesta espècie similar al que fa per P. aphthosa, afegint que és la primera vegada que es cita a Catalunya.
Localitats: Conca de Barberà, Vimbodí, La Baltassana, sota el cim al vessant obac en un talús sobre molses, 1295 m, CF3376, A. Lluent, 1-6-2015. Osona, Vidrà, prop del Castell de Milany, 1400 m, DG4168, leg. A. Lluent, 17-7-2018, det. A. Gómez-Bolea (BCN-Lich 21919). Ripollès, baga de Queràs, sobre la soca de Sorbus aucuparia i les roques molsoses de sota, orientació NE, 1775 m, DG4093, A. Lluent, 25-62013. Vallès Oriental, Montseny, Fogars de Monclús, cap els Castellets, 1350 m, DG5426, leg. A. Lluent, 5-2013, det. A. Gómez-Bolea (BCN-Lich 21918).
Peltigera elisabethae Gyeln. (Fig. 22)
Localitats: Pallars Sobirà, Alt Àneu, Bosc de Marimanha, avetosa, 1680-1820 m, CH3935, leg. A. Lluent, 15-6-2017, det. A. Gómez-Bolea (BCN-Lich 21920).
Peltigera leucophlebia (Nyl.) Gyeln. (Fig. 23)
Pertany a la secció Chloropeltigera (Miadlikowska & Lutzoni, 2000). També té cefalodis a la cara superior del taŀlus com P. apthosa, però la cara inferior té venes més o menys ben definides.
Localitats: Alt Urgell, Bescaran, obaga del Paluc, talús de pista forestal, 2005 m, CG8397, A. Lluent, 10-8-2015. Alta Ribagorça, Muntanyó de Llacs, obaga pedregosa amb fort pendent, terrícola, 2275 m, CH2811, leg. A. Lluent, 218-2018, det. A. Gómez-Bolea (BCN-Lich 21921). Conca de Barberà, Vimbodí, La Baltassana, obaga sota el cim, en una pineda de pi roig, sobre sòl amb molses, 1165 m, CF3376, A. Lluent, 1-6-2015. Pallars Sobirà, Alt Àneu, La Peülla sobre el port de la Bonaigua, neretar a 2320 m, CH3423, A. Lluent, 20-9-2018. Ripollès, baga de Queràs, sota el Roc de Queràs, canal humida sobre roques molsoses al mig de l’avetosa en orientació nord, 1850 m, DG4093, A. Lluent. 25-6-2013.
Peltigera neckeri Hepp ex Müll. Arg. (Fig. 24)
Localitats: Alt Urgell, sobre Arcabell, bordes d’Escàs, pineda de pi roig, obaga, 1610 m, CG7697, leg. A. Lluent, 108-2015, det. A. Gómez-Bolea (BCN-Lich 21922). Ripollès, vessant E del Gra de Fajol Petit, sobre roques granítiques molsoses 1995 m, DG3996, A. Lluent, 17-9-2013.
Peltigera polydactylon (Neck.) Hoffm. (Fig. 25)
Llenas (1909) la cita del voltants de Barcelona, Montseny, Olot, Espluga de Francolí i Cap de Creus.
Localitats: Maresme, Can Canyamars, prop de Can Cames, en un talús, de roca granítica molt alterada, orientat al nord, 250 m, DG5405, 7-10-2018, leg. et det. A. Gómez-Bolea, (BCN-Lich 21923).
Peltigera rufescens (Weiss) Humb. (Fig. 26)
Llenas (1909) la cita de Montserrat, Montseny, boscos del Monestir de Poblet, hi afegeix «etc.», com indicant que és comuna.
Localitats: Alt Urgell, sobre Arcabell, al sud del refugi d’Arcavell, pineda de pi negre, 1940 m, CG7897, A. Lluent, 10-8-2015. Pallars Sobirà, Bosc de Marimanha, bosc d’Abies alba, 1700 m, CH3935, leg. A. Lluent, 15-6-2017, det. A. Gómez-Bolea (BCN-Lich 21924).
* Ramalina thrausta (Ach.) Nyl. (Fig. 27)
Per TLC detectem els àcids úsnic i perlatòlic.
Localitats: Pallars Sobirà, Alt Àneu, Vall d’Aran, bosc de Marimanha, diversos punts dins del bosc, en clarianes o branques d’avet ben iŀluminades, 1750 m, CH3935, leg. A. Lluent, 15-6-2017, det. A. Gómez-Bolea (BCN-Lich 21926).
10 Butlletí ICHN 85 (1), 2021
A. LLUENT & A. GÓMEZ-BOLEA

11 Butlletí ICHN 85 (1), 2021 GEA, FLORA ET FAUNA
DISTRIBUCIÓ DE MACROLÍQUENS A CATALUNYA AMB NOVES DADES COROLÒGIQUES SOBRE LÍQUENS PROTEGITS 19 20 21 22 23 24
Figures 19-24. 19) Parmotrema hypoleucinum; 20) Peltigera aphthosa; 21) Peltigera collina; 22) Peltigera elisabethae. 23) Peltigera leucophlebia; 24) Peltigera neckeri
GEA, FLORA ET FAUNA
* Ricasolia amplissima (Scop.) De Not. (= Lobaria amplissima (Scop.) Forssell) (Fig. 28)
Malgrat ser una espècie força freqüent a la Val d’Aran, pràcticament no s’ha citat en aquesta comarca. La trobem per tota la vall tan en avets com en faigs on és més abundant. Contribuïm aquí amb algunes localitats per anar completant la corologia d’aquesta espècie a la comarca aranesa. La localitat de la Vall de Boí és la primera fora de la Vall d’Aran o adjacent a Catalunya i en una ecologia poc freqüent.
Localitats: Alta Ribagorça, Vall de Boí, Obaga de l’Estany Llebreta, a les roques laterals d’una canal, 1786 m, CH2612, leg. A. Lluent, 7-9-2017, det. A. Gómez-Bolea (BCN-Lich 21927). Alta Ribagorça, Vilaller, barranc de Bessiberris, 1775 m, CH1719, leg. A. Lluent, juny 2017, det. A. GómezBolea (BCN-Lich 21928).Val d’Aran, Vall de Canejan, sota el Barratge deth Hons dera Coma, 1180 m, sobre el refugi dera Honderia, 1030 m, principalment sobre faig, CH2028, A. Lluent, 20-10-2017. Val d’Aran, eth Portillon, dins del recinte de l’Aran Park, sobre faig i avet, 1175 m, CH0937, A. Lluent, 21-10-2017. Val d’Aran, Artiga de Lin, als Uelhs deth Joeu, 1395 m, CH1228, A. Lluent. 21-10-2017, d’aquesta localitat ja havia estat citada per Azuaga & Gómez-Bolea (1996).
* Roccella phycopsis (Ach.) Ach. (Fig. 29)
És relativament freqüent al litoral de Selva i l’Empordà. Destaquen les localitats del massís del Garraf per el seu isolament i la distància al mar.
Localitats: Baix Empordà, Begur, entre les cales de Aiguablava i des Tamariu, diversos punts al litoral rocós i també en pins un xic a l’interior, 5-100 m, EG1742, EG1842, X. Santjoan, 30-12-2018. Baix Empordà, Calonge, Cap de les Penyes Blanques, sobre roques protegides de les onades del mar a la vora del camí de ronda, 5-10 m, EG0730, A. Lluent, 1-7-2018. Baix Empordà, Palamós, castell de Sant Esteve de Mar, a les parets del propi castell i a les roques litorals entre el castell i la cala s’Alguer, 5-20 m, EG1234, X. Sanjuan, 28-2-2019. Baix Empordà, Palamós, entre la cala dels Capellans i la platja de Torre Valentina, sobre roques litorals, 5-10 m, EG0731, X. Sanjuan, 15-12-2018. Baix Empordà, Palafrugell, litoral entre Tamariu i Llafranc en diversos indrets, sobre roques però també en pins, 5-30 m, EG1740, A. Lluent, 15-7-2014. Baix Empordà, Platja d’Aro, cala Pedrosa, roques litorals i també alguns tal·lus en pins, 5-10 m, EG0426, X. Sanjuan, 6-10-2019. Baix Empordà, Platja d’Aro, cales de Pi i Belladona, roques litorals, 5-10 m, EG0630, X. Sanjuan, 28-2-2019. Baix Empordà, Sant Feliu de Guíxols, punta del Canyet, sobre roques litorals, 5-25 m, DG9823, X. Sanjuan, 5-7-2017. Baix Empordà, Sant Feliu de Guíxols, cala Romeguer, sobre roques litorals, 2-10 m, DG9923, X. Sanjuan, 5-7-2017. Garraf, Sitges, Pic del Martell, paret d’escalada, 300 m, DF0969, A. Gómez-Bolea, febrer 2015. Baix Llobregat, Begues, Vessant N del Bosc de Can Grau, sobre conglomerats i gresos silícics, verticals o quasi, 342 m, DF1176, L. Sáez, 23-4-2016. Baix Llobregat, Begues, Roca del Barret, sobre gresos sobre conglomerats i gresos silicis, verticals o quasi, 295 m, DF1276, L. Sáez, 23-4-2016.
* Seirophora contortuplicata (Ach.) Frödén (= Xanthaptychia contortuplicata (Ach.) S.Y. Kondr. & Ravera; = Teloschistes contortuplicatus (Ach.) Clauzade & Rondon) (Fig. 30)
Localitats: Alta Ribagorça, Vall de Boí, Muntanyó de Llacs, a l’extrem nord de la pleta d’Erdo, en fissures de roques calcàries, 2275 m, CH2810, leg. A. Lluent, 27-82018, det. A. Gómez-Bolea (BCN-Lich 21933). Berguedà, Bagà, al sud del coll de la Vall, en escletxes d’un petit aflorament rocós, 2360 m, DG0885, leg. A. Lluent, 2-7-2019, det. A. Gómez-Bolea (BCN-Lich 21932). Berguedà, Bagà, Serrat Gran, en escletxes d’afloraments rocosos dispersos. 2320 m, DG0985, A. Lluent, 22-6-2020. Cerdanya, Das, Coma Oriola, a l’est de la canal, roques extraplomades calcàries, 2205 m, DG0886, leg. A. Lluent, 28-7-2016, det. A. Gómez-Bolea (BCN-Lich 21931). La Cerdanya, Das, Coma Oriola, a l’oest de la canal, petits afloraments rocosos en escletxes profundes i concavitats, 2250 m, DG0786, A. Lluent, 28-7-2016. La Cerdanya, Alp, a banda i banda de coma Pregona, en escletxes de roques calcàries, 2290 m, DG0986, A. Lluent, 10-8-2020. Pallars Sobirà, Espot, Cim de l’Encantat Gran, 2745 m, CH3714, A. Gómez-Bolea, 23-08-2001. Ripollès, Queralbs, al nord del Torrent de Puigmal, en extraploms calcaris, 2250 m, DG2991, leg. A. Lluent, 6-8-2013, det. A. Gómez-Bolea (BCN-Lich 21929). Ripollès, Setcases, Solell de la Coma de l’Orri, en fissures de roques calcaries extraplomades, 2395 m, DG3895, leg. A. Lluent, 7-8-2013, det. A. Gómez-Bolea (BCN-Lich 21930). Fins ara al Ripollès només es coneixia a la coma de les Mulleres (Navarro et. al. 1990, Gaya et. al. 2009) on també l’hem pogut observar. A la localitat del Torrent de Puigmal només hem vist 4 tal·lus, a la coma de l’Orri és més abundant. Les localitats de Coma Oriola, Coma Pregona, Serrat Gran i de coll de la Vall representen una novetat per a l’eix Cadí-Moixeró i per a la Cerdanya i el Berguedà, on colonitza fissures i extraploms de roques calcàries encarades al sud.
* Seirophora lacunosa (Rupr.) Frödén (= Teloschistes lacunosus (Rupr.) Savicz) (Fig. 31)
Considerat en perill d’extinció a Catalunya, només ha estat citat de Torà (Gaia, 2009) i de Castellfollit de Riubregós (Llimona, 1973) tot i que també és conegut a la serra d’Almenara (ICHN), de la Sentiu de Sió (J.A. Conesa com. per.) i la serra Llarga (J.A. Conesa com. pers.). Aquestes són les primeres localitats que no és troben en guixos. A més la de la Granadella és la primera al sud de la plana de Lleida.
Localitats: Anoia, Castellfollit de Riubregós, sobre cal Balaga, en mates en una brolla gipsícola, 485 m, CG6827, leg. A. Lluent, 1-4-2018, det. A. Gómez-Bolea (BCN-Lich 21934). Les Garrigues, Les Sorts, a l’est de La Granadella, en brolles calcícoles, 515 m, CF0580,. A. Lluent, 27-6-2013. Molt escassa i en regressió possiblement per la pertorbació causada per conills. Urgell, Tossal de Montalbà, Vilagrassa, en matollars esclarissats sobre gresos poc salins, 350 m, CG4009, A. Mayoral, 12-7-2015.
12 Butlletí ICHN 85 (1), 2021
A. LLUENT & A. GÓMEZ-BOLEA
13 Butlletí ICHN 85 (1), 2021 GEA, FLORA ET FAUNA
DISTRIBUCIÓ DE MACROLÍQUENS A CATALUNYA AMB NOVES DADES COROLÒGIQUES SOBRE LÍQUENS PROTEGITS 25 26 27 28 29 30
Figures 25-30. 25) Peltigera polydactylon; 26) Peltigera rufescens; 27) Ramalina thrausta; 28) Ricasolia amplissima; 29) Roccella phycopsis; 30) Seirophora contortuplicata
GEA, FLORA ET FAUNA
14 Butlletí ICHN 85 (1), 2021
A. LLUENT & A. GÓMEZ-BOLEA 31 32 33 34 35 36
Figures 31-36. 31) Seirophora lacunosa; 32) Solorina bispora Nyl. subsp. bispora; 33) Solorina saccata; 34) Solorina spongiosa; 35) Sticta limbata; 36) Thamnolia vermicularis.
Solorina bispora Nyl. subsp. bispora (Fig. 32)
Localitats: Ripollès, Queralbs, sobre la font de la Dou, en escletxes ombrívoles amb terra entre els cingles, 2175 m, DG2991, leg. A. Lluent, 6-8-2013, det. A. Gómez-Bolea (BCN-Lich 21935).
Solorina saccata (L.) Ach. (Fig. 33)
Llenas (1909) la cita de les esquerdes i les roques de Montserrat, on nosaltres encara no l’hem pogut retrobar. També la cita de Sant Llorenç, Moià, Espluga de Francolí, Ribas i Camprodon.
Localitats: Berguedà, Berga, obaga de Queralt, en acumulacions de terra entre roques calcàries, 1150 m, DG0262, A. Lluent, 18-8-2018. Garrotxa, Vall de Bianya, El Callís, talús amb molses, 538 m, DG5076, E. Llop, 5-4-2014. Garrotxa, Sant Cristòfol les Fonts, 500 m, DG5868, herbari BC, Antoni de Bolòs, 1918 (E. Llop comunicació personal). Ripollès, Queralbs, sobre la font de la Dou, en escletxes ombrívoles amb terra entre els cingles, 2175 m, DG2991, leg. A. Lluent,
6-8-2013, det. A. Gómez-Bolea (BCN-Lich 21936). Ripollès, Setcases, talussos vora la carretera vora el pont del Catllar en acumulacions de terra entre roques protegides de la pluja. 1165 m, DG4189, A. Lluent, 25-6-2018.
Solorina spongiosa (Ach.) Anzi (Fig. 34)
Localitats: Pallars Sobirà, Llessui, a la Cabaneta, en terra molt humida, 2150 m, CH3704, leg. A. Lluent, 6-9-2017, det. A. Gómez-Bolea (BCN-Lich 21937).
* Sticta limbata (Sm.) Ach. (Fig. 35)
Localitats: Selva, Osor, Sant Miquel de les Formigues, sobre roques amb molses, 1140 m, DG6141, Francesc Canaleta, 3-6-2020.
Thamnolia vermicularis (Sw.) Schaer. (Fig. 36)
Localitats: Alta Ribagorça, Muntanyó de Llacs, sobre la Font del Graller, en prat rocós en fort pendent orientat al nord,
15 Butlletí ICHN 85 (1), 2021 GEA, FLORA ET FAUNA
Figura 37-39. 37) Vulpicida juniperinus; 38) Waynea stoechadiana; 39) Xalocoa ocellata
LÍQUENS PROTEGITS 37 38
DISTRIBUCIÓ DE MACROLÍQUENS A CATALUNYA AMB NOVES DADES COROLÒGIQUES SOBRE
39
GEA, FLORA ET FAUNA
Entremig de molses, 2225 m, CH2811, A. Lluent, 21-8-2018. Alta Ribagorça, Vall de Boí. cresta de Bessiberris, entre el pas de Trescazes i el Bessiberri sud, 2915 m, CH2118, leg. A. Lluent, 24-8-2017, det. A. Gómez-Bolea (BCN-Lich 21938).
Vulpicida juniperinus (L.) J.-E. Mattsson & M. J. Lai (Fig. 37)
Rarament trobem aquesta espècie amb apotecis, com en la localitat del Collet de Baciver.
Localitats: Alta Ribagorça, Vall de Boí, cresta de Bessiberris, entre el pas de Trescazes i el Bessiberri sud, 2915 m, CH2118, leg. A. Lluent, 24-8-2017, det. A. Gómez-Bolea (BCN-Lich 21939). Cerdanya, Lles de Cerdanya, Vall de la Llosa prop de la cabana dels Esparvers, sobre Juniperus communis, 2100 m, CH9304, 3-5-1986, leg. i det. A. GómezBolea (BCN-Lich 21940). Pallars Sobirà, Alt Àneu, estany del Collet de Baciver, carena des de l’estany vers al sud, terrícola en un collet ventat, 2376 m, CH3528, leg. A. Lluent, 8-8-2018, det. A. Gómez-Bolea (BCN-Lich 21941). Ripollès, Queralbs, obaga de la Coma de les Mulleres, com a terrícola en prats acidòfils, 2425 m, DG3194, leg. A. Lluent, 1-82013, det. A. Gómez-Bolea (BCN-Lich 21942). Les formes terrícoles s’havien separat sota el nom de Vulpicida tubulosus (Schaer.) J.-E. Mattsson & M. J. Lai. A partir d’estudis moleculars del gènere fets per Saag. et al. (2014), sense cap estudi morfològic ni anatòmic de les espècies seqüenciades, proposen sinonimitzar aquesta espècie amb V. juniperinus.
* Waynea stoechadiana (Abbassi Maaf & Cl. Roux) P. Clerc & Cl. Roux (= Hypocenomyce stoechadiana Abbasi Maaf & Cl. Roux) (Fig. 38)
Localitats: Barcelonès, Barcelona, Jardí Botànic Històric, sobre Fraxinus pennsylvanica. 60 m, DF2979, A. GómezBolea, 20-4-2018. Barcelonès, Barcelona, jardins de Can Sentmenat, sobre Quercus ilex, 175 m, DF2583, A. GómezBolea, 27-10-2019. Montsià, Amposta, barranc de Coll Llarg, sobre un parell de garrofers vells separats uns 30 m, 140 m, BF9303, A. Lluent, 24-7-2017.
* Xalocoa ocellata (Fr.) Kraichak, Lücking & Lumbsch (Fig. 39)
Localitats: Anoia, El Bruc, prop del Castell, sobre conglomerats, 500 m, CG9804, A Gómez-Bolea, 2-2019. Berguedà, Gironella, sobre l’ermita de Sant Marc, afloraments de roques més o menys plans, 500-525 m, DG0652, A. Lluent, 253-2018. Berguedà, Gironella, sobre cal Paleta, afloraments de roques més o menys plans, 460 m, DG0753, A. Lluent, 25-3-2018. Pallar Jussà, Aramunt, Aramunt Vell al voltant de la torre, afloraments de roques a terra, 640 m, CG3474, A. Lluent, 15-4-2018. Pallars Sobirà, Baix Pallars, boca nord del túnel de Custoia, sobre roques inclinades però no verticals, 570 m, CG3784, A. Lluent, 15-9-2017.
Agraïments
Volem agrair a en Xavier Sanjuan, Gerard Gímenez, Llorenç Sáez, Dolors Rodriguez i en Francesc Canaleta ha-
ver-nos cedit les seves citacions per a poder-les publicar en aquest article. Al Dr. Esteve Llop per l’aportació de dades i els comentaris.
Referències
Azuaga, T. & Gómez-Bolea, A. 1996. Lichens et champignons lichénicoles récoltés dans la région du Val d’Aran (Pyrénées), Espagne. Epiphytes et terricoles.; Bulletin Informations Association Française de Lichénologie, 21 (1): 39-47.
Boluda, C. G.; Rico, V. J., Crespo, A., Divakar, P. K. & Hawksworth, D. L. 2015. Molecular sequence data from populations of Bryoria fuscescens s. lat. in the mountains of central Spain indicates a mismatch between haplotypes and chemotypes. The Lichenologist 47 (5): 279-286.
Burgaz, A. R. & Ahti, T. 2009. Cladoniaceae. Flora Liquenológica Ibérica. Vol. 4. Sociedad Española de Liquenología (SEL). Madrid. ISSN: 1696-0513. 111 p.
Burgaz, A. R. & Martínez, I. 1999. The genus Nephroma Ach. in the Iberian Peninsula. Cryptogamie, Mycologie, 20 (3): 225-235.
Burgaz, A. R. & Martínez, I. 2003. Peltigerales: Lobariaceae, Nephromataceae, Peltigeraceae. Flora liquenológica Ibérica. Sociedad Española de Liquenología (SEL). 42 p.
Diari Oficial de la Generalitat de Catalunya Núm. 685420.4.2015.
Gaya, E. 2009. Taxonomical revision of the Caloplaca saxicola group (Teloschistaceae, lichen-forming Ascomycota). Bibliotheca Lichenologica, 101: 1-191.
Hafellner, J. 1995. Towards a better circumscription of the Acarosporaceae (lichenized Ascomycotina, Lecanorales). Cryptogamic Botany, 5: 99-104.
Hladun, N. 2020. Mòdul LiqueCat. Banc de Dades de Biodiversitat de Catalunya. Generalitat de Catalunya i Universitat de Barcelona. Disponible a: http://biodiver.bio.ub.es/biocat/homepage.html
ICHN. 2015. Itinerari Serra d’Almenara: Ermita de Sant Jaume. Disponible a: https://ichn2.iec.cat/WebSortides/SERRA_ALME–NARA /SERRA_ALMENARA_pagines/Almenara_punt2.htm.
Llenas, M. 1905. Enumeración y distribución de Peltigeráceos de Cataluña. Boletin Real Sociedad Española de Historia Natural, 5: 168-175.
Llenas, M. 1909. Ensaig d’una flora liquènica de Catalunya. Butlletí de la institució Catalana d’Història Natural, 6 (1-6): 1-39.
Llop, E.; Fernandez, S.; Figueras, G.; Muñiz, D. & Llimona, X. 2012. Aproximació al coneixement de la flora liquènica i dels fongs liquenícoles dels altiplans i conques centrals de Catalunya: Sector segàrric. Butlletí de la institució Catalana d’Història Natural, 77: 39-59.
Llop, E. & Hladun, N. L. 2003. Aportació al coneixement de la flora liquènica del massís de Cadiretes (Girona, NE península Ibèrica). Butlletí de la institució Catalana d’Història Natural, 71: 39-50.
Llop, E., Barbero, M., Hladun, N., Navarro, P. & Gómez, A. 2018. Diversity and ecology of lichens and lichenicolous fungi in PNAESM. Butlletí de la institució Catalana d’Història Natural, 82: 121-132.
Miadlikowska, J. & Lutzoni, F. 2000. Phylogenetic revision of the genus Peltigera (lichen-forming ascomycota) based on morphological, chemical, and large subunit nuclear ribosomal DNA data. International Journal of Plant Sciences 161: 925-958.
Miadlikowska, J., Magain, N., Pardo-de la Hoz, C. J., Niu, D., Goward, T., Sérusiaux E. & Lutzoni, F. 2018. Species in section
16 Butlletí ICHN 85 (1), 2021
A. LLUENT & A. GÓMEZ-BOLEA
Peltidea (aphthosa group) of the genus Peltigera remain cryptic after molecular phylogenetic revision. Plant and Fungal Systematics 63 (2): 45-64.
Myllys L, Lindgren H, Aikio S, Häkkinen, L. & Högnabba, F. 2016. Chemical diversity and ecology of the genus Bryoria section Implexae (Parmeliaceae) in Finland. The Bryologist, 119: 29-38.
Llimona, X. 1973. Las comunidades de líquenes de los yesos de España. Tesis Doctoral Univ. Barcelona. 342 p.
Navarro-Rosinés, P. & Hladun, N. L. 1990. Flora liquénica de las rocas carbonatadas del valle de Nuria (Pirineos, Cataluña). Monografías del Instituto Pirenaico de Ecología, 5: 75-83.
Nimis, P. L. & Martellos, S. 2020: ITALIC - The Information System on Italian Lichens. Version 6.0. University of Trieste, Dept. of Biology.
Roux C. & coll., 2020.– Catalogue des lichens et champignons lichénicoles de France métropolitaine. 3e édition revue et augmentée (2020). Édit. Association française de lichénologie (AFL), Fontainebleau. 1769 p.
Saag L., Mark K., Saag A. & Randlane T. 2014. Species delimitation in the lichenized fungal genus Vulpicida (Parmeliaceae, Ascomycota) using gene concatenation and coalescent-based species tree approaches. American Journal of Botany, 101 (12): 2169-2182.
Schumm, F. & Elix, J. A. 2015. Atlas of Images of Thin Layer Chromatograms of Lichen Substances. Herstellung und Verlag: Books on Demand GmbH, Norderstedt ISBN: 978-3-73926103-4. 584 p.
Velmala, S., Myllys, L., Goward, T., Holien, H. & Halonen, P. 2014. Taxonomy of Bryoria section implexae (Parmeliaceae, Lecanoromycetes) in North America and Europe, based on chemical, morphological and molecular data. Annales Botanici Fennici, 51: 345-371.
Zapata, J., Rabasa, M., Vilasís, D., Llop, E. & Oliver, X. 2016. Caracterització i seguiment de localitats de Lobaria pulmonaria (L.) Hoffm. a la Garrotxa i el Ripollès. Pòster presentat a les IV Jornades de conservació de Flora i Funga. Olot.
17 Butlletí ICHN 85 (1), 2021
GEA, FLORA ET FAUNA
DISTRIBUCIÓ DE MACROLÍQUENS A CATALUNYA AMB NOVES DADES COROLÒGIQUES SOBRE LÍQUENS PROTEGITS
L’associació Bartsia trixaginis-Scirpoidetum holoschoeni a
Catalunya
Jordi Bou Manobens* & Lluís Vilar Sais*
* LAGP-Flora i Vegetació. Institut de Medi Ambient. Universitat de Girona. Campus de Montilivi. 17003 Girona.
Autor per a la correspondència: Jordi Bou: A/e: jordi.bou.manobens@gmail.com
Rebut: 12.02.2021; Acceptat: 01.03.2021; Publicat: 31.03.2021
Resum
Els prats dels sorrals fixats del litoral del Bartsia trixaginis-Scirpoidetum holoschoeni O. Bolòs 1962, constitueixen una comunitat de gran interès naturalístic per la seva singularitat. La distribució coneguda fins el moment a Catalunya es limitava al Baix Llobregat, on Bolòs (1962) va descriure l’associació a partir d’un sol inventari fet el 1950. En el decurs d’un estudi de la vegetació dunar de les comarques de Girona s’han trobat noves localitats per aquesta associació a l’Alt Empordà i el Baix Empordà, arribant així a un total de sis localitats. L’escrit presenta 12 inventaris inèdits que permeten analitzar la comunitat des dels punts de vista fitosociològic i ecològic. Aquests prats de rereduna tenen una distribució i extensió molt limitada degut a la destrucció de l’hàbitat i a la pèrdua de les dinàmiques naturals. La variabilitat de l’associació permet diferenciar fins a tres varietats ecològiques noves, sempre marcades per la presència d’espècies dels Thero-Brachypodietea. Es tracta d’una comunitat efímera, de transició entre les unitats dunars de l’Ammophilion arundinaceae i la vegetació mediterrània llenyosa, amb un elevat interès de conservació i un alt grau d’amenaça. Mitjançant les observacions i inventaris fets, l’escrit posa en rellevància aquests prats i la necessitat d’aprofundir en el seu coneixement per tal de poder preservar-los dins d’un litoral profundament transformat.
Paraules claus: Hàbitat d’Interès Comunitari, Empordà, fitosociologia, vegetació dunar, prat, Bartsia trixago, successió ecològica.
Abstract
The Bartsia trixaginis-Scirpoidetum holoschoeni association in Catalonia
The coastal fixed sands grassland of Bartsia trixaginis-Scirpoidetum holoschoeni O. Bolòs 1962, comprises a community of great ecological interest due to its unique character. Up to now, its known distribution in Catalonia has been limited to the county of Baix Llobregat, where Bolòs (1962) described the association on the basis of a single inventory made in 1950. In the course of a dune vegetation study conducted in the province of Girona, new localities for this association were identified in the Alt Empordà and Baix Empordà counties, bringing the total number of localities to six. This paper presents 12 unpublished inventories that have enabled us to analyse the community from phytosociological and ecological perspectives. These backdune meadows are very limited in both distribution and extent, due to habitat destruction and the loss of natural dynamics. The variability of the association allows us to distinguish up to three new ecological varieties, always marked by the presence of Thero-Brachypodietea species. The community is ephemeral, in transition between Ammophilion arundinaceae dune units and Mediterranean vegetation, and is of great conservation interest as well as being highly threatened. Based on the observations and inventories made, this paper highlights the importance of these meadows and the need to increase our knowledge of them, in order to preserve them within a profoundly transformed coastline.
Key words: Habitat of Community Interest, Empordà, phytosociology, dune vegetation, grassland, Bartsia trixago, ecological succession.
Introducció
Els ecosistemes dunars del litoral català són un element singular del patrimoni natural del territori, i ofereixen nombrosos serveis ecosistèmics (Defeo et al., 2009; Everard et al., 2010; Valls et al., 2017; Rodríguez-Revelo et al., 2018), entre els que destaquen ser una barrera de protecció pels hàbitats del litoral davant els temporals i l’erosió (Judge et al., 2003; McLachlan & Brown, 2006; Pries et al., 2008; Taylor et al., 2015). Per altra banda, la flora que contenen aquests ambients és molt específica, degut a les condicions ambientals particulars d’aquest hàbitat. De fet, la vegetació dunar és típicament coneguda per la seva zonificació paraŀlela a la línia de costa, que es veu influenciada per diversos factors abiòtics com el vent, l’onatge, les marees, la salinitat del sòl, la granulometria de les sorres, les dinàmiques costeres i la morfologia dunar (Acosta et al., 2007; Doody, 2008; Isermann, 2011).
La vegetació dels sistemes dunars a Catalunya ha estat estudiada florísticament de forma força completa, tal com es pot comprovar amb els nombrosos treballs publicats, (BraunBlanquet et al., 1936; Bolòs, 1950, 1962, 1967; Rivas Goday
19 Butlletí ICHN 85 (1), 2021
FLORA
FAUNA
GEA,
ET
GEA, FLORA ET FAUNA
Butlletí de la Institució Catalana d’Història Natural, 85 (1): 19-28. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition)
DOI: 10.2436/20.1502.01.68
GEA,
& Rivas Martínez, 1958; Bech & Hernández, 1976; Lapraz, 1976; Perdigó & Papió, 1985; Curcó, 1990; Seguí, 1994; Farràs & Velasco, 1994; Franquesa, 1995; Hoyo & González, 2001; Gesti, 2006; Seguí & Batriu, 2009; Vilar & Quintana, 2014). Gràcies a aquests treballs, es coneix amb prou detall la composició típica de les associacions més abundants a les dunes de Catalunya: Cypero mucronati-Agropyretum juncei Kühnholtz-Lordat ex Br.-Bl. 1933, Ammophiletum arundinaceae Br.-Bl. (1921) 1933 i Crucianelletum maritimae Br.-Bl. 1933. En canvi no es el cas per altres comunitats de rereduna, en part perquè estan pobrament representades al territori, a causa dels impactes sobretot antròpics al litoral.
Actualment els sistemes dunars es troben entre els ecosistemes més amenaçats tant a nivell europeu com global (Hesp & Martínez, 2007; Schlacher et al., 2007; Janssen et al., 2016; Marcenò et al., 2018). Els factors que més amenacen la seva biodiversitat són el desenvolupament urbanístic, seguit del sobreús recreacional d’aquests espais (Janssen et al., 2016). Durant les últimes dècades, les modificacions antròpiques del paisatge litoral (Martí, 2005; Malavasi et al., 2013), conjuntament amb el creixement de la indústria turística i totes les activitats relacionades (Romano & Zullo, 2014; Mir-Gual et al., 2015), han danyat severament els fràgils sistemes dunars (Garcia, 2019; Sperandii et al., 2019a). També s’han de considerar la proliferació de les espècies invasores (Janssen et al., 2016), que causen grans impactes sobre les comunitat vegetals (Marcantonio et al., 2014; Del Vecchio et al., 2015a; Sperandii et al., 2018). Per últim, no es pot ometre el context de canvi climàtic, que pot afectar a la vegetació dunar alterant el període vegetatiu, facilitant l’expansió d’espècies termòfiles, tant natives como exòtiques (Sobrino Vesperinas et al., 2001; UNEP-MAP-RAC/SPA, 2010; Del Vecchio et al., 2015b). Lamentablement, aquest context europeu es repeteix a Catalunya on les últimes dècades es calcula que el 60 % de sistemes dunars han desaparegut i que el 30 % s’han reduït de dimensió, quedant només el 10 % en un estat de conservació favorable (Garcia, 2019).
Al tractar-se d’hàbitats amb un grau alt d’amenaça, es dediquen molts esforços per a la seva conservació (Roig-Munar, 2016; Sperandii et al., 2019b). De fet, tots els hàbitats dunars de Catalunya són hàbitats d’interès comunitari a nivell europeu (HIC; CEE, 1992), amb les implicacions en gestió i conservació corresponents. En el cas concret que pertoca en aquest treball, el HIC 2240 Dunes amb prats d’anuals dels Thero-Brachypodietalia, és un hàbitat representat a Catalunya per una única associació Bartsia trixaginis-Scirpoidetum holoschoeni que només era coneguda a una sola localitat al delta del Llobregat. Aquesta associació va ser descrita per Bolòs (1962), a partir d’un sol inventari publicat d’aquesta comunitat (Bolòs, 1950), que inicialment s’havia identificat com una forma del Crucianelletum maritimae on la successió ja havia avançat amb plantes del Thero-Brachypodion i de les garrigues. La informació que es té sobre el Bartsia trixaginis-Scirpoidetum holoschoeni és molt limitada degut a la seva raresa i a què només es disposa de la interpretació fitosociològica de l’inventari en qüestió que va fer Bolòs (1962) i una breu descripció en el Manual dels hàbitats de Catalunya (Carreras et al., 2017).
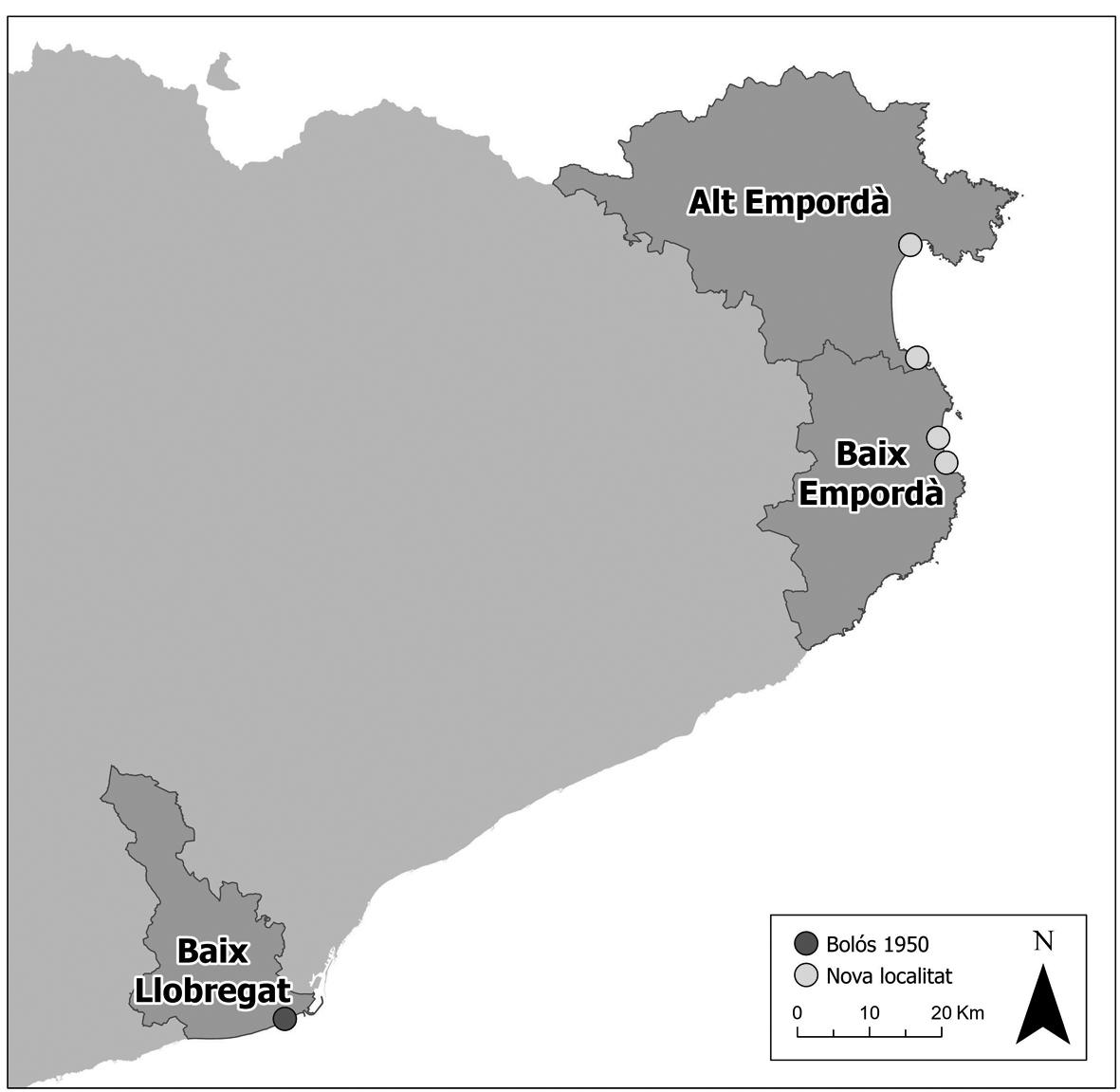
Figura 1. Àrea d’estudi. En vermell, la localitat on ja es tenia constància (Bolòs, 1950) del pradell de Bartsia trixago i, en groc, les noves localitats que s’aporten en aquest estudi.
En el decurs d’un estudi per aprofundir en el coneixement de les comunitats dunars a Catalunya, s’han realitzat tot un seguit d’inventaris als sistemes dunars amb els que s’ha pogut identificar per primer cop al litoral de l’Empordà la presència del Bartsia trixaginis-Scirpoidetum holoschoeni fora del delta del Llobregat (Fig. 1). Aquest estudi té un caire fitosociològic, però també tracte aspectes claus de les dinàmiques del paisatge dels sistemes dunars i aporta noves de dades sobre la singularitat i l’interès de conservació d’aquesta comunitat.
Materials i mètodes
El territori estudiat en el present treball comprèn el litoral, entre les comarques de l’Alt Empordà i del Baix Empordà. Les localitats estudiades es troben dins del Parc Natural dels Aiguamolls de l’Empordà i del Parc Natural del Montgrí, les Illes Medes i el Baix Ter; a excepció d’un inventari de la Rovina situat just a la perifèria de l’àmbit del Parc, però dins de la xarxa Natura 2000. Una descripció detallada de les característiques del territori es pot trobar a Gesti (2006), Vilar & Quintana (2014) i Bou & Jover (2016).
L’estudi i la caracterització de les comunitats s’ha realitzat mitjançant l’aixecament de 12 inventaris segons el mètode sigmatista de Braun-Blanquet (Braun-Blanquet, 1964), amb la determinació de totes les espècies i una anàlisi i catalogació dels inventaris. Pel que fa a la denominació dels tàxons, pel noms científics s’ha seguit la nomenclatura de la Flora Manual dels Països Catalans (Bolòs et al., 1993), excepte per les espècies que donen nom a l’associació. Alhora s’ha disposat d’un inventari inèdit cedit per Jordi Carreras i Albert Ferré d’un nou punt a la localitat de Gavà (Baix Llobregat).
20 Butlletí ICHN 85 (1), 2021
J. BOU MANOBENS & L. VILAR SAIS
FLORA ET FAUNA
Resultats i discussió
Descripció fitosociològica
Ass. Bartsia trixaginis-Scirpoidetum holoschoeni O. Bolòs 1962 nom. corr. (art. 44) [Bellardio-Holoschoenetum australis O. Bolòs 1962 nom. inept.]; pradell de Bartsia trixago dels sorrals fixats del litoral.
Prat d’anuals de sovint 30-70 cm d’alçària i amb un recobriment del sòl del 70-95 %, ric en teròfits alguns de les quals poden ser força abundants, com ara Lagurus ovatus o Rumex bucephalophorus (Fig. 2). També s’hi fan alguns geòfits primaverals entre els quals algunes orquídies. Els hemicriptòfits són freqüents, tot i que tenen un rol secundari, com Dactylis glomerata subsp. hispanica, mentre que la presència de camèfits no és molt rica, però més o menys constant com en el cas d’Helianthemum apenninum. Finalment, els faneròfits no hi són molt presents, però conjuntament amb algun camèfit (Ex: Thymus vulgaris) donen un aspecte diferencial respecte la resta de vegetació dunar.
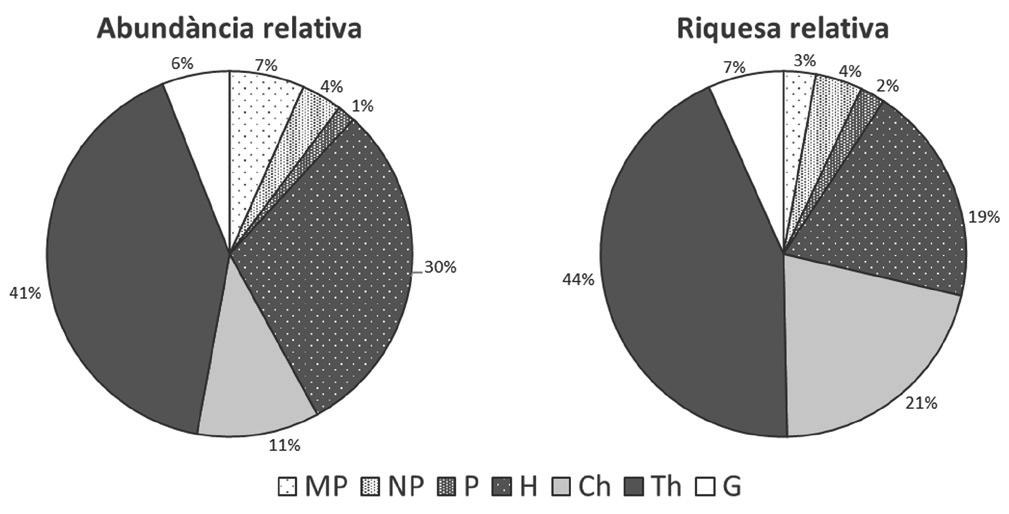
Figura 2. Espectres de formes vitals de la comunitat en abundància relativa i riquesa relativa. Abreviacions: MP, macrofaneròfits; NP, nanofaneròfits; P, faneròfits; H, hemicriptòfits; Ch, camèfits; Th, teròfits; G, geòfits.
L’associació Bartsia trixaginis-Scirpoidetum holoschoeni es diferencia per la presència de l’espècie característica Bartsia trixago, i per l’abundància dels tàxons propis de la Class. Thero-Brachypodietea, entre els que cal destacar Scabiosa atroporpurea, Trifolium scabrum, Catapodium rigidum i Silene nocturna. Aquest conjunt de plantes sol anar acompanyat d’espècies psammòfiles (Teucrium polium subsp. dunense o Medicago littoralis) que diferencien l’associació respecte els altres prats secs de l’aliança Thero-Brachypodion. També hi ha plantes silicícoles de l’Ord. Helianthemetalia guttati, com és el cas d’Helianthemum guttatum o Aira caryophyllea.
Cal destacar que a diferència dels inventaris del delta del Llobregat de Bolòs (1950), cap dels inventaris actuals no té presència de Scirpoides holoschoenus, essent només present un tàxon característic de la Class. Molinio-Arrhenatheretea (Linum angustifolium subsp. usitatissimum). Però si que s’observa un clar caràcter de prat sec de l’All. Thero-Brachypodion que el separa de les comunitats de plantes arenícoles que es troben en els sistemes dunars.
Pel que fa a les formes corològiques, les més freqüents són les plantes mediterrànies i les pluriregionals (Fig. 3). Tot i que la freqüència d’espècies exòtiques és baixa, a la localitat hi hem observat Carpobrotus edulis, que es podria estendre fàcilment per l’hàbitat.
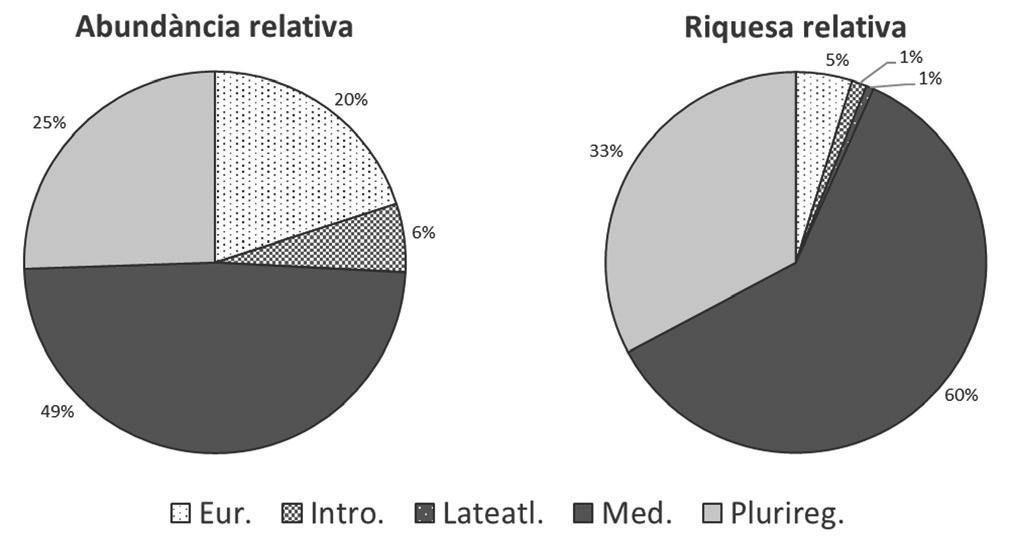
i riquesa relativa. Abreviacions:
Els pradells de Bartsia trixaginis-Scirpoidetum holoschoeni de mitjana tenen una riquesa florística de 22 tàxons per inventari, sense gaires espècies singulars. Cal destacar la presència d’Astragalus tragacantha, espècie protegida dins del Pla d’Espais d’Interès Natural (Annex 3 del Decret 328/1992, de 14 de desembre, per el que s’aprova el Pla d’Espais d’Interès Natural. Espècies de la flora estrictament protegides) com una planta singular dins dels espais naturals protegits del Cap de Creus, Illes Medes i Baix Ter (Generalitat de Catalunya, 2008).
Holotypus
Bolòs (1950: 50) [sub Crucianelletum maritimae Br.-Bl. (1921) 1933 etapa avanzada de sucesión hacia el Trero-Brachypodion]; designat a Bolòs (1962: Taula 22) [sub BellardioHoloschoenetum australis] (art. 18).
Sintaxonomia
Associació descrita per Bolòs (1962) a partir d’un inventari del delta del Llobregat, amb el nom Bellardio-Holoschoenetum australis, que seguint l’article 44 del Codi Internacional de Nomenclatura Fitosociològica (Theurillat et al., 2021), s’ha d’anomenar Bartsia trixaginis-Scirpoidetum holoschoeni O. Bolòs 1962 nom. corr. El nom emprat a Bolòs (1962) provenia de l’espècie característica de l’associació Bellardia trixago, que actualment s’inclou dins del gènere Bartsia (Benedí, 2009), així com d’anomenar el jonc boval Holoschoenus vulgaris Link. i de considerar la varietat australis, però actualment el tàxon s’anomena Scirpoides holoschoenus (L.) Soják. i no hi ha un consens sobre el valor taxonòmic d’aquesta raça, per la qual cosa s’opta per anomenar el sintàxon com a Bartsia trixaginis-Scirpoidetum holoschoeni
Els nous inventaris concorden amb la presència de plantes de la Class. Thero-Brachypodietea observades fins ara, i
21 Butlletí ICHN 85 (1), 2021 GEA, FLORA ET FAUNA L'ASSOCIACIÓ BARTSIA TRIXAGINIS-SCIRPOIDETUM HOLOSCHOENI A CATALUNYA
Figura 3. Espectres de les corologies de la comunitat en abundància relativa
Eur., eurosiberiana; Intro., introduïda; Lateatl., late-atlàntica; Med., mediterrània; Plurireg., pluriregional.
GEA, FLORA ET FAUNA
n’afegeixen de noves per aquesta associació. La diferència més notable és l’absència de Scirpoides holoschoenus en el nous inventaris, ja que a l’holotypus hi era força abundant. Tanmateix, la composició florística global, l’ecologia i la corologia coincideixen i permeten adscriure els nous inventaris al Bartsia trixaginis-Scirpoidetum holoschoeni
Aquesta associació va ser descrita per Bolòs (1962) dins l’All. Thero-Brachypodion, ja que es tractava de prats amb una important riquesa d’espècies d’aquesta aliança. Posteriorment Rivas-Martínez et al. (2001) proposen de situar aquesta associació dins de l’All. Molinio-Holoschoenion vulgaris Br.-Bl. ex Tchou 1948, de l’Ord. Holoschoenetalia vulgaris Br.-Bl. ex Tchou 1948, de la Class. Molinio-Arrhenatheretea Tüxen 1937. Aquesta proposta no seria adequada si s’analitzen el conjunt d’inventaris disponibles actualment, ja que hi ha poca presència d’espècies de prats humits. Per aquest motiu, seguim el criteri original proposat per Bolòs (1962).
Dins de l’associació hi distingim tres variants ecològiques:
Var. Catapodium rigidum. Variant típica i la més propera als Thero-Brachypodion. Les espècies diferencials són: Catapodium rigidum, Arenaria serpyllifolia i Petrorhagia prolifera. Inv.: 1-9.
Var. Thymelaea hirsuta. Variant menys madura i més propera a l’Ammophilion arundinaceae. Les espècies diferencials són: Thymelaea hirsuta i Medicago marina. Inv.: 12 i 13.
Var. Cistus albidus. Variant més madura de l’associació, amb abundància d’espècies llenyoses. Les espècies diferencials són: Cistus albidus i Pinus halepensis. Inv.: 10 i 11.
Existeix també un inventari a Bolòs (1950: 66), que l’autor descriu com una fase de transformació del Crucianalletum cap a l’associació rica en geòfits. Després d’analitzar l’inventari s’ha de concloure que es tracta també del prat de Bartsia trixago, tot i que aquest tàxon és absent a l’inventari. De fet al ser una fase en transformació, no es pot acabar d’identificar com a cap de les varietats ecològiques trobades.
Distribució
Les úniques localitats on s’havia observat aquesta associació fins el moment eren al delta del Llobregat (Baix Llobregat), al costat de l’estany de l’Illa, a uns 8-10 m s.n.m. i a Gavà, a l’estany de la Murtra (Bolòs, 1950). Actualment la primera d’aquestes localitats ha desaparegut, mentre que a la segona s’han observat pradells de poca superfície (Jordi Carreras, com. pers.). Ara s’hi afegeixen les noves localitats de les comarques de Girona, ampliant així l’àrea de distribució de l’associació a l’Empordà. Les localitats de l’Alt Empordà es troben aïllades entre si, la de més al nord a la Rovina a 0,8 m s.n.m. (Fig. 4), entre un ambient de maresmes i el sistema dunar actiu, i la del sud al Bol Roig, en unes sorres consolidades sobre d’un penya-segat a 10 m s.n.m. En el cas del Baix Empordà s’ha observat l’associació a dues localitats, però també es troben aïllades per diversos elements. La localitat de la Fonollera a 1 m s.n.m., se situa en una rereduna desconnectada del cordó dunar, i a Ràdio Liberty hi ha diversos rodals al llarg de l’espai (3-6 m s.n.m. De moment, només es disposen d’inventaris de Ràdio Liberty. Si bé sempre es trac-

ta de pradells de dimensions reduïdes, en el cas de la Rovina n’hi ha dos una mica més extensos de més de 2000 m2 cada un (Bou et al., 2020).
També existeix una cita d’aquesta associació a Mallorca de Martinez Taberner (1983), però de la qual no aporta cap inventari.
Ecologia i dinàmica vegetal
Aquesta comunitat creix en els sorrals fixats del litoral, en reredunes estables, on les dinàmiques de les sorres són poc actives. La seva distribució potencial semblaria estar relacionada a la distribució de Bartsia trixago i els prats secs dels TheroBrachypodietea, de manera que també podria créixer en altres punts del litoral català, i en d’altres regions de clima mediterrani òptims per aquesta tipologia de tàxons. Actualment té una distribució i extensió molt limitada, segurament degut a la destrucció de l’hàbitat i a la pèrdua de les dinàmiques naturals que afavoreixen l’aparició d’aquests ambients, motiu pel qual fins ara només s’havia localitzat en un sol punt de Catalunya.
Aquest alt grau d’impactes sobre la comunitat fa difícil l’estudi de la seva ecologia, però a partir dels inventaris efectuats i a l’observació de la seva variabilitat, es pot deduir aproximadament la seva dinàmica vegetal. Al tractar-se d’una comunitat de rereduna, l’associació semblaria presentar una successió similar a la del Crucianelletum maritimae típic, que com més estable és l’ambient, més plantes mediterrànies acaben colonitzant la comunitat. En alguns casos hi ha dunes residuals colonitzades per brolles o garrigues al litoral, però l’existència d’aquests prats seria una sèrie successional alternativa. En aquests cas una sèrie més lenta en el procés d’establiment de faneròfits, possiblement lligada a sòls més pobres en nutrients que no afavoreix l’establiment de les plantes llenyoses. En aquests casos la successió porta a l’associació Bartsia trixaginis-Scirpoidetum holoschoeni, que, com demostra la variant ecològica de Thymelaea hirsuta, té certa afinitat amb el Crucianelletum maritimae. En aquests inventaris s’observen amb certa freqüència alguns taxons més propis del Crucianelletum maritimae (Thymelea hirsuta i Medicago marina), els quals, a mesura que madura la comunitat, van desapareixent a favor de plantes característiques dels TheroBrachypodietea, i acaben formant el Bartsia trixaginis-Scirpoi-
22 Butlletí ICHN 85 (1), 2021
J. BOU MANOBENS & L. VILAR SAIS
Figura 4. Pradell de Bartsia trixaginis-Scirpoidetum holoschoeni a la Rovina (Alt Empordà), envoltat de vegetació halòfila.

detum holoschoeni var. Catapodium rigidum. Aquest pradell d’anuals va modificant el sòl, afavorint a la llarga l’entrada de faneròfits, com els pins i les estepes (Fig. 5), fenomen que acaba transformant la comunitat cap al Bartsia trixaginis-Scirpoidetum holoschoeni var. Cistus albidus. Aquesta última varietat ecològica seria més afí a les brolles, i faria pensar que la potencialitat de l’associació estaria relacionada amb una brolla mediterrània i amb pinedes secundàries.
Es tractaria, doncs, de comunitats efímeres de transició entre les unitats dunars de l’Ammophilion arundinaceae i la vegetació mediterrània. El nínxol ecològic compartit per les plantes característiques de l’associació i l’aliança tindrien un espai en el temps curt, ja que només creix en alguns sorrals fixats amb condicions determinades. Tanmateix, és necessari aprofundir en aquestes condicions òptimes per tal de poder detallar més l’ecologia de la comunitat, alhora que s’hauria d’aprofundir més en la dinàmica vegetal d’aquest sintàxon tan singular del sistema dunar de Catalunya.
Interès de conservació
El pradell de Bartsia trixago dels sorrals fixats del litoral només és present a sis localitats, i en la majoria de casos es tracte de superfícies exigües, a excepció de la localitat de la Rovina, on hi ha algun pradell més gran. La seva extensió territorial i forma d’implantació territorial fan d’aquests prats una comunitat de raresa notable. La composició florística més típica d’aquesta comunitat (Bartsia trixaginis-Scirpoidetum holoschoeni var. Catapodium rigidum) presenta una riquesa d’entre 22 i 29 tàxons per inventari, entre els quals es pot trobar Astragalus tragacantha, espècie singular que dóna un elevat interès naturalístic a l’hàbitat. Alhora, el grau de maduresa és elevat, ja que és una comunitat de dunes fixades que segueix a les associacions dunars corresponents. És per aquest motiu que l’hàbitat CORINE “16.229 Llistonars (prats de Brachypodium retusum) i altres prats secs amb teròfits, colonitzadors d’arenys marítims”, a la que s’atribueix aquesta associació té un Interès de Conservació (IC) de 16 sobre 24 (Carreras & Ferré, 2013). Tot i que tenint en compte els nous inventaris el seu IC hauria de ser de 17, ja que la riquesa sol ser superior a 20, de manera que el valor de “Ri-
quesa florística (biodiversitat)” (IC1) hauria de ser de 3, i no pas de 2 com fins ara era considerat.
Les característiques de la comunitat també la fan molt vulnerable a pertorbacions i impactes. La seva raresa al territori és una de les seves principals fragilitats, ja que qualsevol tipus de situació pot comprometre fàcilment gran part de les localitats. A més, el seu caràcter efímer de pradell ric en teròfits fa que sigui una comunitat de transició molt sensible, on la mateixa successió natural tendeix a transformar-la fàcilment. L’entrada d’espècies llenyoses és clarament un element clau a tenir en compte en l’ecologia i la seva conservació. Aquestes característiques són especialment problemàtiques per una comunitat del litoral marítim català, on hi ha hagut una important destrucció d’hàbitats durant el segle XX i a on la pressió turística és molt elevada, ja que la combinació d’aquests dos contextos representa una vulnerabilitat important. Per aquest motiu, el grau d’amenaça considerat per l’hàbitat 16.229 es de 3 (Carreras & Ferré, 2013). Tot i que observant els nous inventaris el seu grau hauria de ser de 4, ja que el nombre de localitats es menor de 20.
Durant la realització dels inventaris s’ha pogut comprovar com aquests pradells estan situats a prop de platges d’elevada freqüentació estival, amb paisatges profundament transformats. En aquest sentit, la localitat de Ràdio Liberty és la que té més minimitzada aquests impactes, ja que l’espai té l’accés tancat al públic. La localitat de l’estany de l’Illa és el cas contrari, on l’ampliació de la tercera pista de l’Aeroport, entre altres equipament, han transformat la vegetació de l’entorn (González et al., 2016), alterant de forma important la vegetació dunar (Jordi Carreras, com. pers.). Pel que fa als usos actuals, també cal tenir en compte que la sobrepastura existent a la Rovina desafavoreix la conservació d’aquest pradell, ja que empobreix la comunitat i afavoreix la presència de Parentucellia viscosa que acaba fent poblaments molt importants (Bou et al., 2020). Com que en tots els casos es tracta d’espais dins de la Xarxa Natura 2000, hauria de ser viable el control d’aquests tipus d’impactes sobre la comunitat. De la mateixa manera que en bona part del litoral, les espècies invasores són una amenaça per aquests pradells, ja que en ocupar poca superfície, fàcilment poden quedar totalment coberts per especies com ara Carpobrotus edulis, que a part d’aparèixer en un inventari de Radio Liberty, també va ser observat a prop d’algun inventari de la Rovina. En darrer lloc, cal considerar la pròpia dinàmica natural d’aquesta comunitat, que tendeix a tancar-se, desapareixent els prats actuals vers brolles i pinedes secundàries. En el paisatge primitiu s’anirien formant nous ambients on creixeria aquesta comunitat a la rereduna, però actualment les dinàmiques dunars estan molt alterades, i només es produiria el primer fenomen. Això implica la necessitat de més estudis per poder planificar una gestió activa especifica, segons els diferents casos, i, si pot ser, la seva restauració.
Considerant l’elevat interès de conservació i el grau alt d’amenaça, juntament amb les observacions fetes en aquest estudi, es posa de manifest la necessitat de l’elaboració d’unes bases estratègiques per a la preservació d’aquests pradells singulars de reredunes. Per una gestió adequada de la vegetació dunar de Catalunya és necessari, doncs, aprofundir en el coneixement de l’ecologia d’aquests pradells de Bartsia trixago, ja que són un element únic d’aquests ecosistemes.
23 Butlletí ICHN 85 (1), 2021 GEA, FLORA ET FAUNA
L'ASSOCIACIÓ BARTSIA TRIXAGINIS-SCIRPOIDETUM HOLOSCHOENI A CATALUNYA
Figura 5. Pradell de Bartsia trixaginis-Scirpoidetum holoschoeni a la Rovina (Alt Empordà), on en primer terme s’observa Pinus halepensis
GEA, FLORA ET FAUNA
Esquema sintaxonomic
Class. Thero-Brachypodietea Br.-Bl. 1947
Ord. Thero-Brachypodietalia (Br.-Bl.) R. Mol. 1934
All. Thero-Brachypodion Br.-Bl. 1925
Suball. Parentucellenion latifoliae Bharucha 1933 em. nom. O.
Bolòs 1981
Ass. Bartsia trixaginis-Scirpoidetum holoschoeni O. Bolòs 1962
Taula 1. Pradell de Bartsia trixaginis-Scirpoidetum holoschoeni.
Seguim aquí l’esquema sintaxonòmic clàssic per aquest sintàxon, si bé sembla que caldria incloure’l a la classe Lygeo sparti-Stipetea tenacissimae (Rivas-Martínez et al., 2001; Mucina et al., 2016).
24 Butlletí ICHN 85 (1), 2021
Número d’inventari 1 2 3 4 5 6 7 8 9 10 11 12 13 14 Àrea estudiada (m2) 25 25 25 25 25 25 25 - - 25 25 25 25Alçària de l’estrat arbori (m) 0 0 0 0 0 0 0 - - 12 8 0 0 12,5 Recobriment de l’estrat arbori (%) 0 0 0 0 0 0 0 - - 70 95 0 0 50 Alçària de l’estrat arbustiu (cm) 0 0 0 0 5 50 80 - - 200 50 0 100Recobriment de l’estrat arbustiu (%) 0 0 0 0 0 10 10 - - 10 40 0 10Alçària de l’estrat herbaci (cm) 70 70 70 70 70 30 50 - - 70 30 70 70Recobriment de l’estrat herbaci (%) 90 90 90 90 95 80 90 - 70 50 20 60 60 60 Recobriment total de la vegetació (%) 90 90 90 90 95 90 90 - 70 100 100 60 60 60 Nombre d’espècies 29 27 32 22 22 22 23 24 21 17 17 15 14 28 Característiques de l’associació Bartsia trixago 1 1 1 + + + + 2 + + + + Característiques de l’aliança, l’ordre i la classe Scabiosa atropurpurea + + + + + 2 2 + Catapodium rigidum subsp. hemipoa + + + + 1 + + Trifolium scabrum + + + + + + Arenaria serpyllifolia + + + + 1 Silene nocturna + + + + + Sedum album + + + + Petrorhagia prolifera + + + + Urospermum dalechampii + + + + Cerastium pumilum + + + + Linum strictum + + 2 1 Plantago albicans + 2 + + Paronychia argentea + + (+) Psoralea bituminosa 2 + + Hedypnois rhagadioloides + + + Vulpia myuros + + + Helianthemum ledifolium + + Brachypodium distachyon + + Centaurea aspera 1 1 Asterolinon linum-stellatum 1 + Hippocrepis multisiliquosa + (+) Argyrolobium zanonii + Filago pyramidata + Hyparrhenia hirta 2 Leontodon taraxacoides + Leontodon tuberosus + Medicago minima + Ophrys tenthredinifera 1 Reichardia picroides subsp. picroides + Ruta chalepensis + Espècies psammòfiles Teucrium polium subsp. dunense + + + + + + (+) + 1 + Medicago littoralis + + + + + Pancratium maritimum + + + + Thymelaea hirsuta + + 1 Elymus farctus subsp. farctus + + Medicago marina + + Echinophora spinosa + + Ephedra distachya 1 + Ononis natrix subsp. ramosissima + 2 Aetheorhiza bulbosa 1 1 Euphorbia terracina + +
BOU MANOBENS & L. VILAR SAIS
J.
Entre parèntesis quan el tàxon era present fora de l’inventari.
Companyes presents només en un inventari
1 Allium sp., Serapias lingua, Juncus bufonius, Sherardia arvensis
4 Inula viscosa, Lathyrus ochrus, Plantago lanceolata, Trifolium angustifolium
5 Asparagus officinalis, Olea europaea, Schoenus nigricans, Torilis arvensis
6 Limonium virgatum 1.
7 Echinops ritro, Romulea ramiflora, Rosmarinus officinalis.
8 Bromus cf. diandrus, Oryzopsis miliacea
10 Euphorbia segetalis, Rhamnus alaternus, Rubia peregrina, Yucca aloifolia
12 Lavandula stoechas Linum trigynum, Silene nutans.
13 Scrophularia canina.
25 Butlletí ICHN 85 (1), 2021 GEA, FLORA ET FAUNA Número d’inventari 1 2 3 4 5 6 7 8 9 10 11 12 13 14 Crucianella maritima + Cutandia maritima + Cyperus capitatus + Euphorbia paralias + Rumex roseus + Silene cf. niceensis + Silene sclerocarpa + Sporobolus pungens + Vulpia membranacea subsp. fasciculata 2 Companyes Lagurus ovatus + 2 + 1 1 3 3 + 1 + 2 1 Avena barbata + + + + + + + + Helichrysum stoechas + + + + + 1 1 + Dactylis glomerata subsp. hispanica + + + + + + + Daucus carota subsp. maritimus + + + + + 1 Alyssum maritimum subsp. maritimum + + + + + + Helianthemum apenninum subsp. apenninum + + + + + + Plantago lagopus 1 + + + + Sedum sediforme + + + 2 + Cistus salviifolius + + 2 + 1 Linum angustifolium subsp. usitatissimum + + + + Anagallis arvensis + + + + Trifolium campestre + + + + Blackstonia perfoliata + 1 + + Rumex bucephalophorus subsp. bucephalophorus 1 1 1 + Aira caryophyllea + + + Plantago crassifolia 2 + 1 Koeleria phleoides + 1 + Thymus vulgaris + + + Orobanche minor + 1 + Dipcadi serotinum + + 1 Pinus halepensis (+) 1 + Pistacia lentiscus + (+) + Astragalus tragacantha 1 1 + Scirpoides holoschoenus 3 3 Limonium narbonense + + Artemisia campestris + + Festuca glauca 1 + Gladiolus sp. + + Lathyrus cf. clymenum + + Parentucellia viscosa + 1 Plantago coronopus + + Polycarpon tetraphyllum + + Psilurus incurvus + + Cynodon dactylon + + Sonchus tenerrimus + + Eryngium campestre 1 + Asparagus acutifolius 1 + Pinus pinea + 3 Cistus albidus + + Carpobrotus edulis (+) + Corynephorus canescens + + Helianthemum
+ +
guttatum
TRIXAGINIS-SCIRPOIDETUM HOLOSCHOENI
CATALUNYA
14 Asphodelus fistulosus, Clematis flammula, Muscari neglectum, Odontides lutea, Ophrys fusca, Sanguisorba sp., Vicia sp. L'ASSOCIACIÓ BARTSIA
A
GEA, FLORA ET FAUNA
Procedència dels inventaris
1 Castelló d’Empúries, sorrals de la Rovina, Alt Empordà EG1178 (11/05/2020)
2 Castelló d’Empúries, sorrals de la Rovina, Alt Empordà EG1178 (11/05/2020)
3 Castelló d’Empúries, sorrals de la Rovina, Alt Empordà EG1178 (11/05/2020)
4 Castelló d’Empúries, sorrals de la Rovina, Alt Empordà EG1178 (11/05/2020)
5 Castelló d’Empúries, sorrals de la Rovina, Alt Empordà EG1178 (11/05/2020)
6 l’Escala, Bol Roig, Alt Empordà EG1262 (25/05/2020)
7 l’Escala, Bol Roig, Alt Empordà EG1262 (25/05/2020)
8 Inventari inèdit de Jordi Carreras i Albert Ferré 2018 (Gavà, camí de la Pineda, Baix Llobregat DF1969)
9 Inventari publicat a Bolòs (1950) i Bolòs (1962) (Estany de l’Illa, Baix Llobregat)
10 Pals, Ràdio Liberty, Baix Empordà EG1648 (8/06/2020)
11 Pals, Ràdio Liberty, Baix Empordà EG1648 (8/06/2020)
12 Pals, Ràdio Liberty, Baix Empordà EG1648 (19/06/2020)
13 Pals, Ràdio Liberty, Baix Empordà EG1648 (19/06/2020)
14 Inventari publicat a Bolòs 1950 (Estany de la Murtra, Baix Llobregat)
Agraïments
Volem agrair a Jordi Carreras i Albert Ferré la cessió d’un inventari de vegetació i els comentaris aportats. Aquest estudi s’ha realitzat dins de diferents projectes, però principalment sota l’empara dels projecte ECOPLAYA del Ministerio de Ciencia, Innovación y Universidades (RTI2018-095677B-I00).
Bibliografia
Acosta, A., Ercole, S., Stanisci, A., Pillar, V. D. P. & Blasi, C. 2007. Coastal vegetation zonation and dune morphology in some Mediterranean ecosystems. Journal of Coastal Research 236: 1518-1524.
Bech, J. & Hernández, A. M. 1976. Estudios sobre suelos y vegetación del delta del Llobregat. Collectanea Botanica (Barcelona) 10: 33-105.
Benedí, J. 2009. Bartsia L. P. 501-505. In: Benedí, C., Rico, E. & Güemes, J. (eds). Flora iberica 13. Real Jardi Botánico. CSIC. Madrid.
Bolòs, A. 1950. La vegetación de las comarcas barcelonesas. Instituto Español de Estudios Mediterráneos. Barcelona. 579 p.
Bolòs, O. 1967. Comunidades vegetales de las comarca próximas al litoral situadas entre los los rios Llobregat y Segura. Memorias la Real Academia de Ciencias y Artes de Barcelona, 38 (1): XX.
Bolòs, O. 1962. El paisaje vegetal barcelonés. Cátedra ciudad de Barcelona. Barcelona. 192 p.
Bolòs, O., Vigo, J., Masalles, R. M. & Ninot, J. M. 1993. Flora manual dels Països Catalans. Editorial Pòrtic, Barcelona. 1247 p.
Bou, J. & Jover, M. 2016. Cartografia digital dels hàbitats CORINE i dels Hàbitats d’Interès Comunitari del Parc Natural del Montgrí, les Illes Medes i el Baix Ter, escala 1:10.000. Disponible en: http://atzavara.bio.ub.edu/geoveg/docs/Memoria_ cartografia_habitats_Montgri2016.pdf. [Data de consulta: 11 gener 2021]
Bou, J., Vilar, L. & Quintana, X. 2020. Cartografía digital de los hábitats de la Rovina, escala 1:5.000. Universitat de Girona. Informe técnico para Tragsatec. 55 p.
Braun-Blanquet, J. 1964. Pflanzensoziologie Grundzüge der Vegetationskunde. Springer Vienna. Vienna. 631 p.
Braun-Blanquet, J., Font Quer, P., Braun-Blanquet, G., Frey, E., Jansenm, P. & Moor, M. 1936. L’excursion de la SIGMA en Catalogne (Pâques 1934). Communication SIGMA num. 38. 37 p.
Carreras, J. & Ferré, A. 2013. Avaluació del grau d’amenaça i de l’interès de conservació dels hàbitats de Catalunya. Universitat de Barcelona. Barcelona. 130 p.
Carreras, J., Ferré, A. & Vigo, J. (eds). 2017. Manual dels hàbitats de Catalunya: catàleg dels hàbitats naturals reconeguts en el territori català d’acord amb els criteris establerts pel CORINE biotopes manual de la Unió Europea. 8 volums. Edició revisada. Departament de Territori i Sostenibilitat.
CEE. 1992. Directiva 92/43/CEE del Consejo de 21 de mayo de 1992 relativa a la conservación de los hábitats naturales y de la fauna y flora silvestres. Bruselas.
Curcó, A. 1990. La vegetació del Delta de l’Ebre (I): les comunitats dunars (Classe Ammophiletea Br.-Bl. et Tx. 1943). Butlletí del Parc Natural del Delta l’Ebre 5: 9-18.
Defeo, O., Mclachlan, A., Schoeman, D. S., Schlacher, T. A., Dugan, J., Jones, A., Lastra, M. & Scapini, F. 2009. Threats to sandy beach ecosystems: A review. Estuarine, Coastal and Shelf Science 81 (1): 1-12.
Del Vecchio, S., Pizzo, L. & Buffa, G. 2015a. The response of plant community diversity to alien invasion: evidence from a sand dune time series. Biodiversity and Conservation. 24: 371-392.
Del Vecchio, S., Prisco, I., Acosta, A. T. R. & Stanisci, A. 2015b. Changes in plant species composition of coastal dune habitats over a 20-year period. AoB Plants 7: 1-10.
Doody, J. P. (ed). 2008. Sand dune inventory of Europe, 2nd ed. National Coastal Consultants and EUCC – The Coastal Union, IGU Coastal Commission. Peterborough. 685 p.
Everard, M., Jones, L. & Watts, B. 2010. Have we neglected the societal importance of sand dunes? An ecosystem services perspective. Aquatic Conservation: Marine and Freshwater Ecosystems 20: 476-487.
Farràs, A. & Velasco, E. 1994. Les comunitats vegetals de les zones humides de l’Alt Empordà. In: Gosálbez, J., Serra, J. & Velasco, E. (eds.) Els sistemes naturals dels aiguamolls de l’Empordà. Institució Catalana d’Història Natural (Treballs de la Institució Catalana d’Història Natural 13). Barcelona. 228 p.
Franquesa, T. 1995. El paisatge vegetal de la península del cap de Creus. Institut d’Estudis Catalans. Barcelona. 628 p.
Garcia, C. 2019. Els sistemes dunars de la costa catalana. Evolució històrica, estat actual i potencial de restauració. Tesi doctoral. Universitat de Girona. Girona. 270 p.
Generalitat de Catalunya. 2008. Decret 172/2008, de 26 d’agost, de creació del Catàleg de flora amenaçada de Catalunya.
Gesti, J. 2006. El poblament vegetal dels Aiguamolls de l’Empordà Institut d’Estudis Catalans, Barcelona. 862 p.
González, V., Hoyo, R., Seguí, J.M. & Valverde, A. 2016. Flora
26 Butlletí ICHN 85 (1), 2021
J. BOU MANOBENS & L. VILAR SAIS
vascular del delta del Llobregat. Institució Catalana d’Història Natural. 514p.
Hesp, P.A. & Martínez, M.L. 2007. Disturbance processes and dynamics in coastal dunes. In: Johnson, E.A. & Miyanishi, K. (ed.). Plant Disturbance Ecology: The Process and the Response. Academic Press, Elsevier. 720p.
Hoyo, R. & González, V. 2001. Anàlisi de l’estat de la vegetació i catàleg florístic del paratge de la pineda de Cal Francès i la seva zona litoral (Viladecans, Delta del Llobregat). Spartina 4: 79–106.
Isermann, M. 2011. Patterns in species diversity during succession of coastal dunes. Journal of Coastal Research 27: 661-671.
Janssen, J.A.M., Rodwell, J.S., García Criado, M., Gubbay, S., Haynes, T., Nieto, A., Sanders, N., Landucci, F., Loidi, J., Ssymank, A., Tahvanainen, T., Valderrabano, M., Acosta, A., Aronsson, M., Arts, G., Attorre, F., Bergmeier, E., Bijlsma, R.J., Bioret, F., Biţă-Nicolae, C., Biurrun, I., Calix, M., Capelo, J., Čarni, A., Chytrý, M., Dengler, J., Dimopoulos, P., Essl, F., Gardfjell, H., Gigante, D., Giusso Del Galdo, G., Hájek, M., Jansen, F., Jansen, J., Kapfer, J., Mickolajczak, A., Molina, J.A., Molnár, Z., Paternoster, D., Piernik, A., Poulin, B., Renaux, B., Schaminée, J.H.J., Šumberová, K., Toivonen, H., Tonteri, T., Tsiripidis, I., Tzonev, R. & Valachovič, M. 2016. European Red List of Habitats. Part 2. Terrestrial and Freshwater Habitats.
European Union. Luxembourg. 44 p.
Judge, E.K., Overton, M.F. & Fisher, J.S. 2003. Vulnerability Indicators for Coastal Dunes. Journal of Waterway, Port, Coastal, and Ocean Engineering 129: 270–278.
Lapraz, G. 1976. Recherches phytosociologiques en Catalogne. Collectanea Botanica X: 205–279.
Malavasi, M., Santoro, R., Cutini, M., Acosta, A.T.R. & Carranza, M.L. 2013. What has happened to coastal dunes in the last half century? A multitemporal coastal landscape analysis in Central Italy. Landscape and Urban Planning 119: 54–63.
Marcantonio, M., Rocchini, D. & Ottaviani, G. 2014. Impact of alien species on dune systems: a multifaceted approach. Biodiversity and Conservation 23: 2645–2668.
Marcenò, C., Guarino, R., Loidi, J., Herrera, M., Isermann, M., Knollová, I., Tichý, L., Tzonev R.T., Acosta, A.T.R., Fitzpatrick, U., Iakushenko, D., Janssen, J.A.M., Jiménez-Alfaro, B., Kącki, Z., Keizer-Sedláková, I., Kolomiychuk, V., Rodwell, J.S., Schaminée, J.H.J., Šilc, U. & Chytrý, M. 2018. Classification of European and Mediterranean coastal dune vegetation. Applied Vegetation Science 21: 533–559.
Martí, C. 2005. La transformació del paisatge litoral de la Costa Brava: Anàlisi de l’evolució (1956-2003), diagnosi de l’estat actual i prognosi de futur. Tesi doctoral. Universitat de Girona. Girona. 631p.
Martinez Taberner, A. 1983. La franja dunar de la badía d’Alcúdia (Mallorca) I. Estat actual de la maquia de Juniperus oxycedrus L. subsp. macrocarpa (Sibth. et S.) Ball. Bolletí de la Societat d’Història Natural de les Balears 27: 7–22.
Mclachlan, A. & Brown, A.C. (eds). 2006. The ecology of sandy shores. Elsevier. Burlington. 392p.
Mir-Gual, M., Pons, G.X., Martín-Prieto, J.A. & Rodríguez-Perea, A. 2015. A critical view of the Blue Flag beaches in Spain using environmental variables. Ocean and Coastal Management 105: 106–115.
Mucina, L., Bültmann, H., Dierßen, K., Theurillat, J., Raus, T., Čarni, A., Šumberová, K., Willner, W., Dengler, J., García, R.G., Chytrý, M., Hájek, M., Di Pietro, R., Iakushenko, D., Pallas, J., Daniëls, F., Bergmeier, E., Santos Guerra, A., Ermakov, N., Valachovič, M. Schaminée, J., Lysenko, T., Didukh, Y., Pignatti, S., Rodwell,
J., Capelo, J., Weber, H., Solomeshch, A., Dimopoulos, P., Aguiar, C., Hennekens, S. & Tichý, L. Vegetation of Europe: hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Applied Vegetation Science 19 (S1): 3-264.
Perdigó, M.T. & Papió, C. 1985. La vegetación litoral de Torredembarra (sud de Catalunya). Collectanea Botanica 16: 215–226.
Pries, A.J., Miller, D.L. & Branch, L.C. 2008. Identification of Structural and Spatial Features that Influence Storm-Related Dune Erosion along a Barrier-Island Ecosystem in the Gulf of Mexico. Journal of Coastal Research 4: 168–175.
Rivas-Martínez, S., Fernández-González, F., Loidi, J., Lousã, M. & Penas, A. 2001. Syntaxonomical checklist of vascular plant communities of Spain and Portugal to association level. Itinera Geobotanica 14: 5–341.
Rivas Goday, S. & Rivas Martínez, S. 1958. Acerca de la Ammophiletea del Este y Sur de España. Anales del Instituto Botánico A. J. Cavanilles. XVI: 549–564.
Rodríguez-Revelo, N., Espejel, I., Arredondo García, C., OjedaRevah, L. & Sánchez-Vázquez, M.A. 2018. Environmental Services of Beaches and Coastal Sand Dunes as a Tool for Their Conservation. In: Botero, C., Cervantes, O. & Finkl, C.W. (eds) Beach Management Tools - Concepts, Methodologies and Case Studies. Springer International Publishing AG.
Roig-Munar, F.X. (ed). 2016. Restauració i gestió de sistemes dunars. Esudi de casos. Càtedra de Geografia i Pensament Territorial. Girona. 222p
Romano, B. & Zullo, F. 2014. The urban transformation of Italy’s Adriatic coastal strip: Fifty years of unsustainability. Land use policy 38: 26–36.
Schlacher, T.A., Dugan, J., Schoeman, D.S., Lastra, M., Jones, A., Scapini, F., Mclachlan, A. & Defeo, O. 2007. Sandy beaches at the brink. Diversity and Distributions 13: 556–560.
Seguí, J.M. 1994. Alguns aspectes de la vegetació del Delta del Llobregat. Direcció General del Medi Natural. DARP. Generalitat de Catalunya.
Seguí, J.M. & Batriu, E. 2009. Línies d’estudi de la biodiversitat al delta del Llobregat. Anàlisi, estructura i composició de les comunitats vegetals psammòfiles. Any 2009. Consorci per la Protecció i Gestió dels Espais Naturals del Delta del Llobregat.
Sobrino Vesperinas, E., González Moreno, A., Sanz Elorza, M., Dana Sánchez, E., Sánchez Mata, D. & Gavilán, R. 2001. The expansion of thermophilic plants in the Iberian Peninsula as a sign of climatic change. In: Walther, G.R., Burga, C.A. & Edwards, P.J. (eds) “Finger- prints” of cliamte change. Kluwer Academic/Plenum Publisher. New York. 184p
Sperandii, M.G., Bazzichetto, M., Acosta, A.T.R., Barták, V. & Malavasi, M. 2019a. Multiple drivers of plant diversity in coastal dunes: A Mediterranean experience. Science of The Total Environment 652: 1435–1444.
Sperandii, M.G., Bazzichetto, M., Gatti, F. & Acosta, A.T.R. 2019b. Back into the past: Resurveying random plots to track community changes in Italian coastal dunes. Ecological Indicators 96: 572–578.
Sperandii, M.G., Prisco, I. & Acosta, A.T.R. 2018. Hard times for Italian coastal dunes: insights from a diachronic analysis based on random plots. Biodiversity and Conservation 27: 633–646. Springer Netherlands.
Taylor, E.B., Gibeaut, J.C., Yoskowitz, D.W. & Starek, M.J. 2015. Assessment and Monetary Valuation of the Storm Protection Function of Beaches and Foredunes on the Texas Coast. Journal of Coastal Research 315: 1205–1216.
27 Butlletí ICHN 85 (1), 2021 GEA, FLORA ET FAUNA
L'ASSOCIACIÓ BARTSIA TRIXAGINIS-SCIRPOIDETUM HOLOSCHOENI A CATALUNYA
GEA, FLORA ET FAUNA
Theurillat, J.,Willner, W., Fernández‐González, F., Bültmann, H., Čarni, A., Gigante, D., Mucina, L. & Weber, H. 2021. International Code of Phytosociological Nomenclature. 4th Edition. Applied Vegetation Science 24 (1): 1-62.
UNEP-MAP-RAC/SPA. 2010. Impact of climate change on marine and coastal biodiversity in the Mediterranean Sea: current state of knowledge. RAC/SPA Edit. Tunis. 28 p.
Vilar, L. & Quintana X. (eds.) 2014. El poblament vegetal i els hàbitats del massís del Montgrí, les illes Medes i la plana del Baix Ter. Càtedra d’Ecosistemes Litorals Mediterranis. Girona. 252 p. J.
Valls, J.F., Rucabado, J., Sardá, R. & Parera, A. 2017. The beach as a strategic element of governance for Spanish coastal towns. Journal of Sustainable Tourism 25: 1338-1352.
28 Butlletí ICHN 85 (1), 2021
BOU MANOBENS & L. VILAR SAIS
Dos noves espècies del gènere Parvospeonomus Bellés & Escolà, 1977 (Coleoptera: Leiodidae: Leptodirini)
Jordi Comas*
* Museu de Ciències Naturals de Barcelona. Laboratori de Natura. Passeig Picasso, s/n. 08003 Barcelona. A/e: comasj2@gmail.com
Rebut: 18.11.2020; Acceptat: 08.03.2021; Publicat: 31.03.2021
Resum
En el present treball, es descriuen dues noves espècies del gènere Parvospeonomus Bellés & Escolà, 1977, que han restat confoses amb Parvospeonomus urgellesi (Español, 1965): Parvospeonomus garrigai n. sp. i Parvospeonomus cruillensis n. sp., per la similitud de la seva morfologia dels habitus. Són mostrades imatges, de les diferències morfològiques entra les tres espècies estudiades.
Paraules clau: Coleoptera, Leiodidae, Leptodirini, Parvospeonomus garrigai n. sp., Parvospeonomus cruillensis n. sp., Catalunya.
Abstract
Two new species of the genus Parvospeonomus Bellés & Escolà, 1977 (Coleoptera: Leiodidae: Leptodirini)
In the present work, two new species of the genus Parvospeonomus Bellés & Escolà, 1977 are described, which have been confused with Parvospeonomus urgellesi (Español, 1965): Parvospeonomus garrigai n. sp. and Parvospeonomus cruillensis n. sp., by the similarity of their morphology of habits. Images are shown, of the morphological differences between the three species studied.
Key words: Coleoptera, Leiodidae, Leptodirini, Parvospeonomus garrigai n. sp., Parvospeonomus cruillensis n. sp., Catalonia.
urn:lsid:zoobank.org:pub:CFBBE504-266F-4924-807F-E00CC772B789
Introducció
El gènere Parvospeonomus Bellés & Escolà, (1977), inicialment descrit com a subgènere de Speonomus Jeannel, (1908), per presentar l’àpex dels paràmers fortament eixamplats de forma ovalada i còncava; posteriorment, Fresneda, (1998), en la revisió efectuada dels Leptodirini (Leptodirinae en el seu moment), treball en què s’estudia l’estructura de sac intern de l’edeagus, canvià l’estatus d’aquest, i l’elevà a la condició actual de gènere; en l’actualitat, aquest gènere compren quatre espècies: P. delarouzeei (Fairmaire, 1860), P. vilarubiasi (Zariquiey,1940), P. urgellesi (Español, 1965) i P. canyellesi (Lagar, 1974), totes ben diferenciades entre elles per la seva morfologia externa.
L’aïllament de la localitat típica de P. urgellesi, la Cova de les Aranyes, feia suposar que la resta de cites, no correspongueren a la mateixa espècie, i, gràcies al material recentment recoŀlectat en la Cova de les Aranyes, ha permès poder comparar tant les morfologies externes, com internes dels exemplars de les diferents poblacions fins al moment conegudes; aquesta revisió, ens ha confirmat l’especiació aŀlopàtrica d’aquestes dues noves espècies diferenciades de P. urgellesi, tal com són presentades en el present treball.
S’ha optat per prendre com a exemplars tipus les femelles, donat que aquestes són les que donen millors caràcters diferenciadors entre les espècies, sent en els mascles escassament apreciables.
Material i mètodes
El material estudiat es troba muntat en cartolina entomològica; les preparacions microscòpiques extretes, han estat durant 24 hores submergides en una solució del 50 % de Lacto-fenol i àcid Acètic, i posteriorment, muntades en immersió de Dimetil-Hidantoïna-Formaldehid (DMHF) sobre una cartolina d’acetat transparent adjuntada a la mateixa agulla de l’exemplar del qual procedeix.
Els acrònims utilitzats són els següents: CXB (Xavier Bellés coŀlecció, Barcelona, Espanya), JCC (Jordi Comas coŀlecció, Barcelona, Espanya), JFC (Javier Fresneda coŀlecció, Llesp, Espanya), MCNB (Museu Ciències Naturals Barcelona coŀlecció, Barcelona, Espanya), ICGC (Institut Cartogràfic i Geològic de Catalunya); i les abreviacions: Cv. (cova) i Av. (avenc).
També s’ha de fer constar, que les mesures preses de l’amplada dels artells de les antenes, són les màximes, donat que els tres darrers que conformen la maça són una mica comprimits.
La cartografia utilitzada per indicar la ubicació de les espècies han estat preses del ICGC.
Resultats
Parvospeonomus urgellesi (Español, 1965)
Speonomus (s. str.) urgellesi Español, 1965
29 Butlletí ICHN 85 (1), 2021 GEA, FLORA ET FAUNA GEA, FLORA
ET FAUNA
Butlletí de la Institució Catalana d’Història Natural, 85 (1): 29-34. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition)
10.2436/20.1502.01.69
DOI:
GEA,
Speonomus (Parvospeonomus) urgellesi (Español): Bellés & Escolà, 1977
Parvospeonomus urgellesi (Español): Fresneda, 1998
Material estudiat.
3 ♂ i 5 ♀, etiquetats: «Av. de les Aranyes / Tordera / (el Maresme) / 25-VI-2020 / J. Pallisé leg. »; «ETRS89 Fus: 31N, 470114, 4611973, 310 m», coordenades en la part posterior (etiqueta blanca). Les genitàlies es troben muntades en immersió de DMHF sobre una cartolina d’acetat transparent, junt l’exemplar del qual han sigut extretes. En la coŀlecció JCC.
Descripció de la ♀
Habitus figura 1a. Longitud 2,32 mm, mesures preses des de la part anterior del pronot fins a l’àpex dels èlitres. Cos de forma oval, fortament convex. Potes curtes.
Pronot de costats regularment arquejat des de la base fins a la part anterior, sent en la meitat anterior menys arquejada que la resta, més estret que els èlitres, presentant la màxima amplada en la seva base (1,2 mm d’amplada màxima, per 0,57 mm de longitud).
Antenes gràcils, arribant a la meitat dels èlitres, longitud 1,23 mm, mesura dels artells segons taula 1a.
Èlitres amb la màxima amplada en el terç anterior, retrocedint progressivament vers l’àpex; la longitud és una sisena part més llarg que ample (1,27 mm d’amplada del conjunt dels èlitres, per 1,75 mm de llargada); estria sutural clarament impresa.
Potes relativament curtes.
VIII segment abdominal, presentant una apòfisi en la zona mitjana ampla, curta i bífida (Fig. 2a).
Espermateca prima i allargada, escassament arquejada, amb la part anterior formant un lòbul engroguit, part basal recte i no engruixida en la inserció del conducta espermàtic i de la glàndula annexa (Fig. 3a).

Descripció del ♂
El ♂, presenta una morfologia externa similar a la de la ♀, i tan sols és diferència pels caràcters sexuals secundaris, presentant les mateixes proporcions tant en allò que respecte a les potes, com a les antenes.
L’edeagus presenta el terç basal un marcat bombament en la part ventral, i la part dorsal en corba continuada (Fig. 4a).
Distribució
Única cavitat coneguda en la que viu aquesta espècie. Catalunya: El Maresme.
Parvospeonomus cruillensis n. sp.
urn:lsid:zoobank.org:act:8F3B4A96-165D-4751-82054174234CB5C0
Material tipus
HOLOTIPUS: ♀, etiquetat: «Cv. Cruïlles / Aiguafreda (Vallès / Oriental) 27-VIII-2020 / Meseguer - Auroux - / Comas - Pallisé leg.»; «ETRS89 Fus: 31N / 438339, 4625636 / 678 m», coordenades en la part posterior (etiqueta blanca); «Parvospeonomus / cruillensis n. sp. / J. Comas det. 2020» (etiqueta vermella). La genitàlia es troba muntada en immersió de DMHF sobre una cartolina d’acetat transparent, junt l’exemplar. Dipositat en la coŀlecció JCC. PARATIPUS: 10 ♂ i 19 ♀, amb les mateixes dades que l’holotipus (etiqueta blanca) «Paratipus / Parvospeonomus / cruillensis n. sp. / J. Comas det. 2020» (etiqueta vermella); 1 ♂ i 2 ♀ «Cv. Cruïlles / Aiguafreda (Vallès / Oriental) 25-X-1964 / Auroux leg.» (etiqueta blanca) «Paratipus / Parvospeonomus / cruillensis n. sp. / J. Comas det. 2020» (etiqueta vermella); 6 ♂ i 5 ♀ «Cv. de l’Infern / Aiguafreda (Vallès / Oriental) / 27-VIII2020 / J. Pallisé leg.» (etiqueta blanca) «Paratipus / Parvospeonomus / cruillensis n. sp. / J. Comas det. 2020» (etiqueta


Figura 1, Habitus de: a) Parvospeonomus urgellesi (Español); b) Parvospeonomus cruillensis n. sp.; c) Parvospeonomus garrigai n. sp. (Escala: 1 mm) (Fotos A. Meseguer).
30 Butlletí ICHN 85 (1), 2021
J. COMAS
FLORA ET FAUNA
a b c

vermella); 1 ♂ i 1 ♀ «Cv. Trencada / Aiguafreda / (Vallès / Oriental) / 31-I-1970 / J. Comas leg.» (etiqueta blanca) «Paratipus / Parvospeonomus / cruillensis n. sp. / J. Comas det. 2020» (etiqueta vermella); 6 ♀ «Av. Enfonsat “A” / Centelles / (Osona) / 10-IV-1971 / J. Comas leg.» (etiqueta blanca) «Paratipus / Parvospeonomus / cruillensis n. sp. / J. Comas det. 2020» (etiqueta vermella); 1 ♂ i 3 ♀ «Av. Enfonsat “A” / Centelles / (Osona) / 25-IV-1971 / J. Comas leg.» (etiqueta blanca) «Paratipus / Parvospeonomus / cruillensis n. sp. / J. Comas det. 2020» (etiqueta vermella). Els paratipus queden repartits en les coŀleccions: JCC, CXB, JFC i MCNB.
Material addicional.
A més, s’ha de fer constar unes restes recollides el 29-IX2020 en la Cova de l’Ermita, Aiguafreda, (Vallès Oriental) per A. Meseguer, les quals han de ser atribuïdes a aquesta nova espècie.
Diagnosi
Parvospeonomus cruillensis n. sp., difereix de P. urgellesi en la morfologia externa, per la forma dels èlitres, sent en P. cruillensis menys acuminats i més amples; per la diferent configuració del VIII segment abdominal per ser l’apòfisi
31 Butlletí ICHN 85 (1), 2021 GEA, FLORA ET FAUNA DOS NOVES ESPÈCIES DEL GÈNERE PARVOSPEONOMUS BELLÉS & ESCOLÀ, 1977
Figura 2. VIII segment abdominal de: a) Parvospeonomus urgellesi (Español); b) Parvospeonomus cruillensis n. sp.; c) Parvospeonomus garrigai n. sp.
a b c a b c
Figura 3. Espermateques de: a) Parvospeonomus urgellesi (Español); b) Parvospeonomus cruillensis n. sp.; c) Parvospeonomus garrigai n. sp.
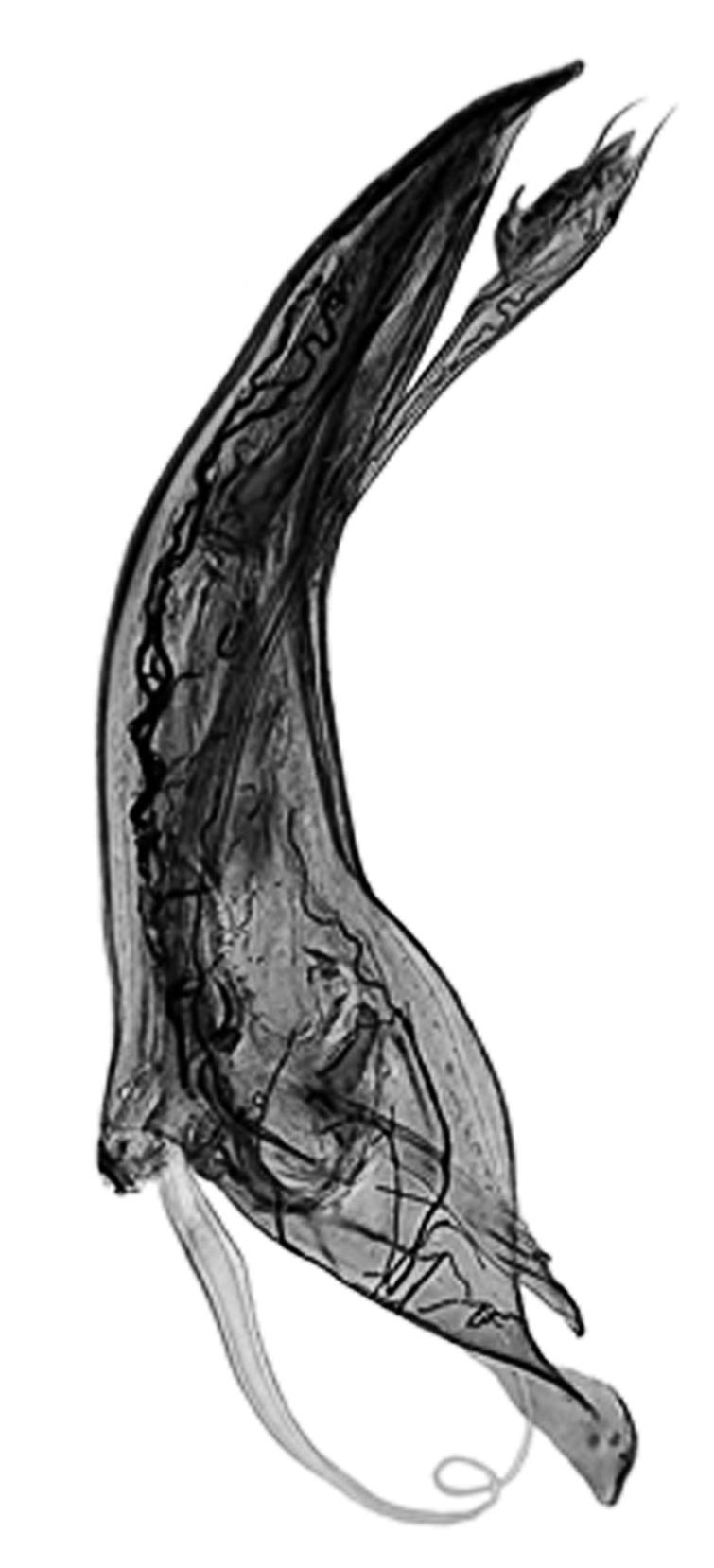
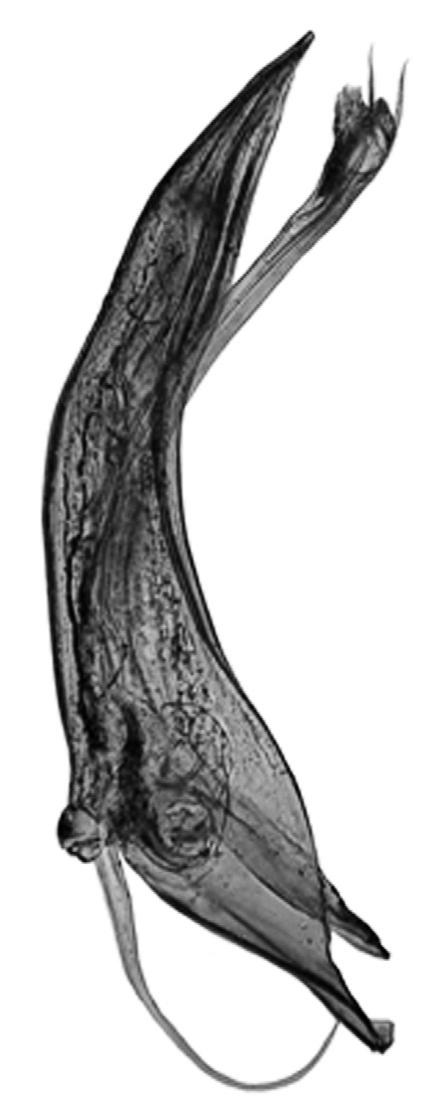
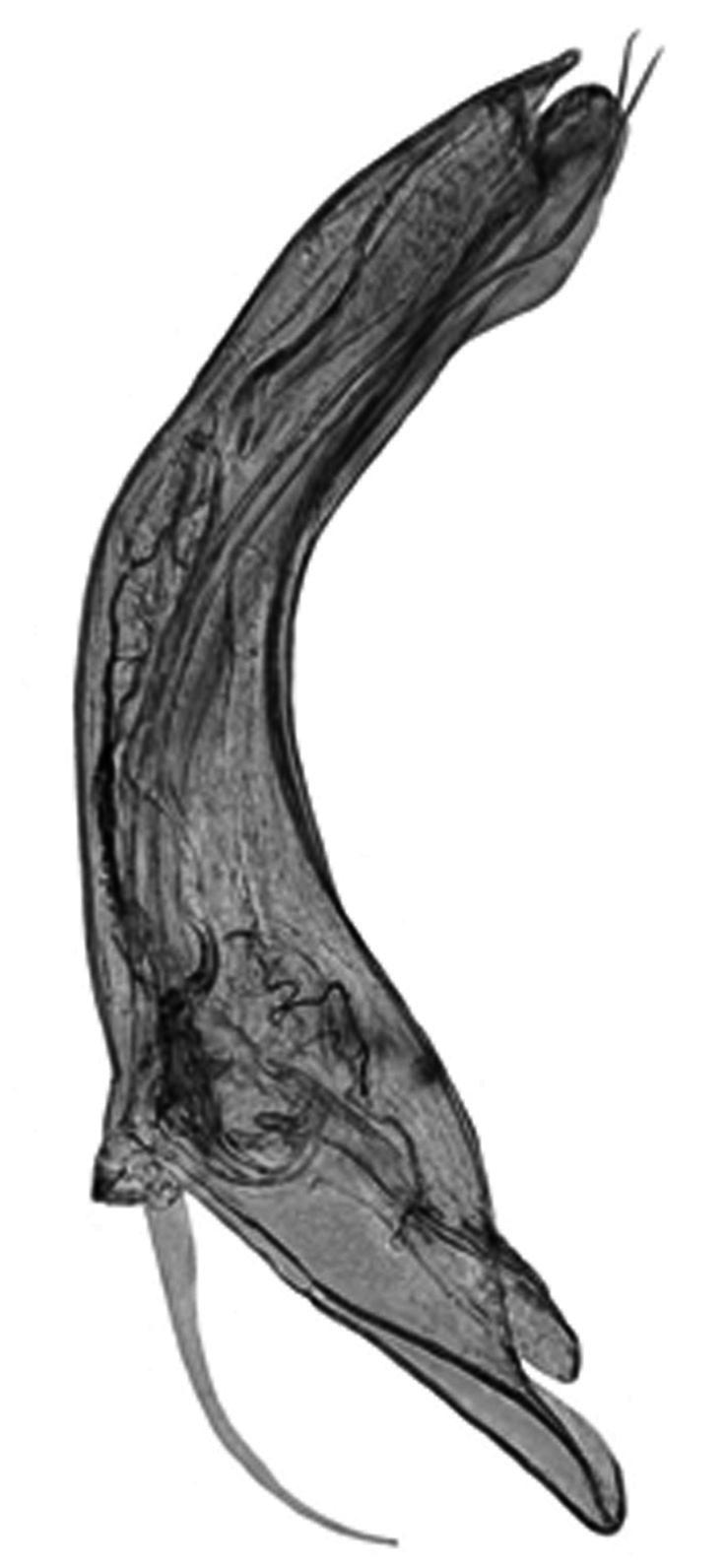
mitjana prima i allargada, i per l’espermateca que és colzada en la part basal i en la part apical.
Descripció de la ♀
Habitus figura 1b. Longitud 2,27 mm, mesures preses des de la part anterior del pronot fins a l’àpex dels èlitres. Cos de forma oval, fortament convex. Potes curtes.
Pronot regularment arquejat, des de la part anterior fins a la posterior, presentant la seva màxima amplada en la base; més estret que els èlitres (1,25 mm d’amplada màxima, per 0,65 mm de longitud).
Antenes gràcils, arribant escassament a la meitat dels èlitres, longitud 1,25 mm mesura dels artells segons taula 1b.
Èlitres amb la màxima amplada en el terç anterior, retrocedint progressivament vers l’àpex; la longitud és una sisena part més llarg que ample (1,37 mm d’amplada del conjunt dels èlitres, per 1,62 mm de llargada); estria sutural present.
Potes relativament curtes.
VIII segment abdominal presentant l’apòfisi de la zona mitjana estreta i allargada (Fig, 2b).
Espermateca prima i allargada, recte, amb l’àpex format un lòbul engroguit, arrodonit i doblegat cap a la seva part ventral; formant la part basal curta i colzada, amb un petit engruiximent en la inserció del conducte espermàtic i de la glàndula annexa (Fig. 3b).
Descripció del ♂
La diferència morfològica entre els mascles i femelles, és nuŀla en excepció dels caràcters sexuals secundaris externs tal com són el diferent nombre d’artells dels protarsos que en els mascles són de cinc, i en les femelles quatre; l’edeagus és poc arquejat presentant en el terç basal de la part ventral poc bombada (Fig. 4b),
Etimologia
La denominació de P. cruillensis, fa referència a la cova de Cruïlles, propera al castell de Cruïlles.
Distribució.
Catalunya: Vallès Oriental i Osona.
Parvospeonomus garrigai n. sp.
urn:lsid:zoobank.org:act:65DF62DA-8200-4531-AC2C404CE14DB6CC
Material tipus
HOLOTIPUS: ♀, etiquetat: «Cv. dels Ermitans / Caldes de Montbui / (Vallès Oriental) / 23-VII-2020 / J. Pallisé leg.»; «ETRS89 Fus: 31N / 428588, 4612604 / 520 m, en la part posterior de l’etiqueta» (etiqueta blanca); «Holotipus / Parvospeonomus / garrigai n. sp. / J. Comas det. 2020» (etiqueta vermella). La genitàlia es troba muntada en immersió de DMHF sobre una cartolina d’acetat transparent, junt l’exemplar. Dipositat en la coŀlecció JCC. PARATIPUS: 5 ♂ i 20 ♀, amb les mateixes dades que l’holotipus, (etiqueta blanca) «Paratipus / Parvospeonomus / garrigai n. sp. / J. Comas det. 2020»; 2 ♂ i 6 ♀ «Cv. de Solanes / Caldes de Montbui / (Vallès Oriental) / 6-20-VII-1986 / Auroux-Filbà leg.» (etiqueta blanca) «Paratipus / Parvospeonomus / garrigai n. sp. / J. Comas det. 2020» (etiqueta vermella); 1 ♂ i 5 ♀ «Av. del Bac / El Figarò / (Vallès Oriental) / 8-II-1970 / J. Comas leg.» (etiqueta blanca) «Paratipus / Parvospeonomus / garrigai n. sp. / J. Comas det. 2020» (etiqueta vermella); 1 ♂ «Av.
32 Butlletí ICHN 85 (1), 2021 GEA, FLORA ET FAUNA
J. COMAS a b c
Figura 4. Edeagus de: a) Parvospeonomus urgellesi (Español); b) Parvospeonomus cruillensis n. sp.; c) Parvospeonomus garrigai n. sp.
DOS NOVES ESPÈCIES DEL GÈNERE PARVOSPEONOMUS BELLÉS & ESCOLÀ, 1977
del Bac / El Figarò (Vallès / Oriental) 19-IX-1971 / J. Comas leg.» (etiqueta blanca) «Paratipus / Parvospeonomus / garrigai n. sp. / J. Comas det. 2020» (etiqueta vermella); 1 ♂ i 1 ♀ «Av. del Toni / El Figarò / (Vallès Oriental) / 9-V-1976 / J. Comas leg.» (etiqueta blanca) «Paratipus / Parvospeonomus / garrigai n. sp. / J. Comas det. 2020» (etiqueta vermella); 5 ♀ «Cv. dels Ermitans / Caldes de Montbui / (Vallès Oriental) / 22-XI-1974 / J. Comas leg.» (etiqueta blanca); «Paratipus / Parvospeonomus / garrigai n. sp. / J. Comas det. 2020» (etiqueta vermella); 4 ♂ i 13 ♀ «Cv. de Solanes / Caldes de Montbui / (Vallès Oriental) / 23-VII-2020 / J. Pallisé leg.» (etiqueta blanca) «Paratipus / Parvospeonomus / garrigai n. sp. / J. Comas det. 2020» (etiqueta vermella). Paratipus repartits en les coŀleccions: JCC, CXB, JFC i MCNB.
Diagnosi
Parvospeonomus garrigai n. sp., difereix de P. urgellesi i P. cruillensis, en la seva morfologia externa per la forma dels èlitres, sent en P. garrigai més llargs i acuminats, mostrant una silueta més gràcil que les altres dues espècies; també separada d’aquestes dues espècies, per la morfologia diferent del VIII segment abdominal presenta l’apòfisi mitjana llarga
i ample, i la de l’espermateca fortament colzada en la base i tan sols engrossida en l’àpex.
Descripció de la ♀
Habitus figura 1c. Longitud 2,25 mm. mesures preses des de la part anterior del pronot fins a l’àpex dels èlitres. Cos de forma oval, fortament convex. Potes curtes.
Pronot arquejat des de la part anterior fins base, sent en la meitat anterior menys arquejada; més estret que els èlitres, presentant la màxima amplada en la seva base (1,25 mm d’amplada màxima, per 0,62 mm de longitud)
Antenes gràcils, arribant escassament a la meitat dels èlitres, longitud 1,27 mm mesura dels artells segons taula 1c.
Èlitres amb la màxima amplada en el terç anterior, retrocedint progressivament vers l’àpex; la longitud, es d’un terç més llarg que ample (1,35 mm d’amplada del conjunt dels èlitres, per 1,63 mm de llargada); estria sutural visiblement impresa.
Potes relativament curtes.
VIII segment abdominal presentant una prolongació o apòfisi en la zona mitjana ampla i allargada (Fig. 2c) Espermateca prima i allargada, escassament arquejada,
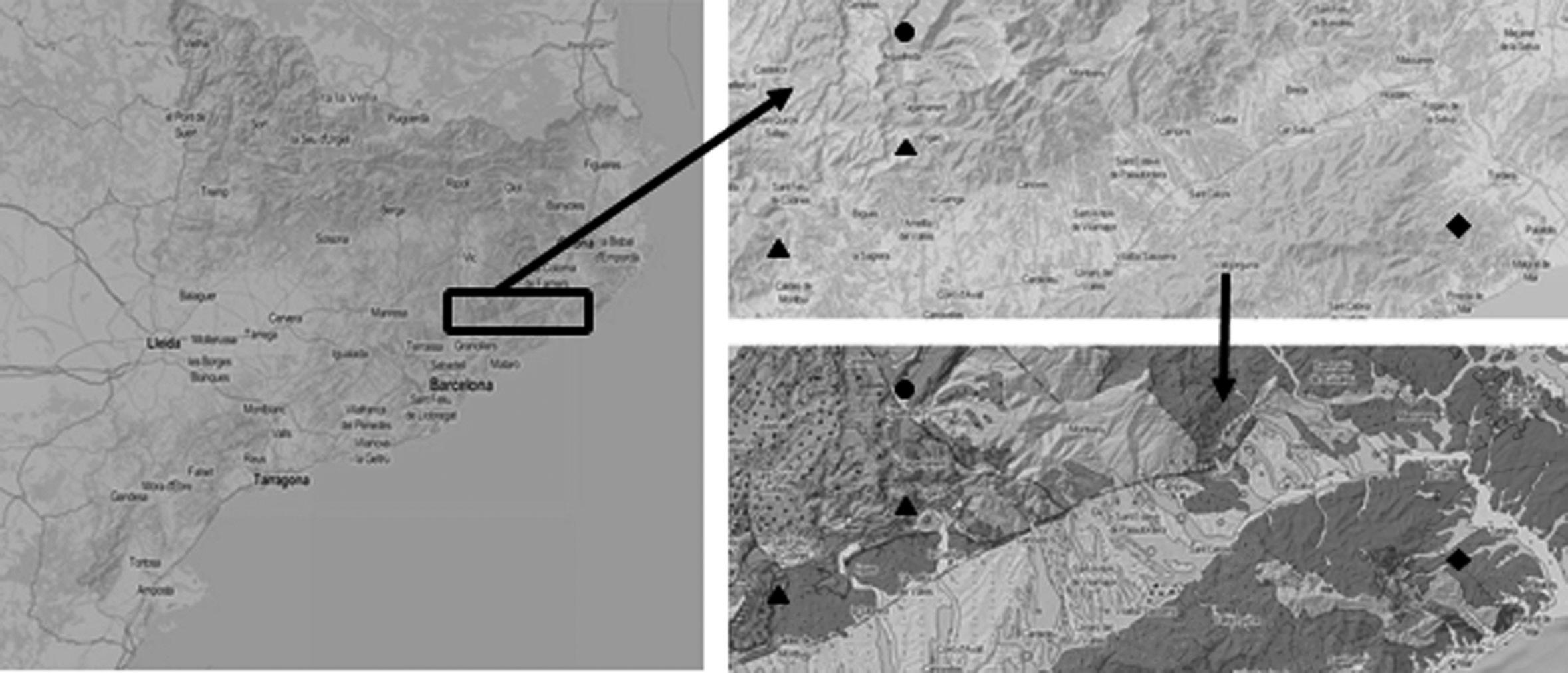
33 Butlletí ICHN 85 (1), 2021 GEA, FLORA ET FAUNA
a I II III IV V VI VII VIII IX X XI Longitud 0,12 0,16 0,12 0,09 0,10 0,11 0,11 0,07 0,10 0,10 0,15 Amplada 0,05 0,05 0,04 0,04 0,04 0,04 0,07 0,05 0,08 0,08 0,08 b I II III IV V VI VII VIII IX X XI Longitud 0,13 0,15 0,12 0,09 0,11 0,10 0,12 0,07 0,10 0,10 0,16 Amplada 0,05 0,05 0,04 0,04 0,04 0,04 0,06 0,05 0,08 0,09 0,09 c I II III IV V VI VII VIII IX X XI Longitud 0,13 0,17 0,13 0,09 0,11 0,11 0,13 0,07 0,10 0,09 0,14 Amplada 0,06 0,05 0,04 0,04 0,04 0,04 0,06 0,04 0,07 0,08 0,08
Taula 1. Formula antenal de: a) Parvospeonomus urgellesi (Español), longitud total: 1,23 mm; b) P. cruillensis n. sp., longitud total: 1,25 mm; c) P. garrigai n. sp. longitud total: 1,27 mm.
Figura 5. Mapa de situació de les espècies: Parvospeonomus urgellesi (Español) (rombe); P. cruillensis n. sp. (triangle); P. garrigai n.sp. (punt). (Extret del ICGC).
GEA, FLORA ET FAUNA
formant un lòbul engroguit en el àpex, la part basal fortament colzada, amb un engruiximent en la inserció del conducta espermàtic i de la glàndula annexa (Fig. 3c).
Descripció del ♂
Igual que l’anterior espècie, la diferència morfològica és nul·la entre els dos sexes, en excepció dels caràcters sexuals secundaris externs; l’edeagus es presenta més arquejat, amb el terç basal de la part ventral poc bombada (Fig. 4c).
Etimologia.
Espècie dedicada al Sr. Martí Garriga i Argullós (Sant Feliu de Codines: 1925-2012), estudiós de les coves i jaciments arqueològics de la comarca del Vallès Oriental i fundador del Museu Arqueològic de Sant Feliu de Codines.
Distribució.
Catalunya: Vallès Oriental.
Comentaris
La distribució aŀlopàtrica de les espècies es pot veure en les seves ubicacions, tal com queden representades en el mapa (Fig. 5), ja que P. urgellesi, es troba aïllada de les altres dues per la vall del riu de la Tordera i pel massís del Montseny, i, P. cruillensis de P. garrigai, pel riu Congost.
Clau d’especies del gènere Parvospeonomus
1 Protarsos dels mascles lleugerament dilatats. Mida molt petita (1,7-1,9 mm) P. canyellesi (Lagar)
– Protarsos dels mascles no dilatats. Individus més grans (1.8-3.0 mm) ...................................................................2
2 Antenes més llargues respecte a la longitud del cos, amb el VIII artell cilíndric, dues vegades més llarg que ample. Mida relativament més gran (2,5-3mm) P. vilarrubiasi (Zariquiey)
– Antenes curtes respecte a la longitud del cos, amb el VIII
artell de forma globulosa, com a màxim una vegada i mitja més llarg que ample. Mida relativament més petita (1,8-2,8mm) ....................................................................3
3 Artell VIII de les antenes netament globulós. Metatíbies curtes. Àpex dels paràmers de l’edeagus poc eixamplats P. delarouzeei (Fairmaire)
– Artell VIII de les antenes vegada i mitja més llarg que ample. Metatíbies llargues, 1/3 de la longitud del cos. Àpex dels paràmers de l’edeagus més eixamplats (Fig. 4)
........................................................................................4
4 Èlitres poc amples amb relació al pronot, escassament arrodonits en l’àpex (Fig. 1c). VIII abdominal de les femelles amb l’apòfisi mitjana ampla i allargada (Fig. 2c)
P. garrigai n. sp.
– Èlitres més amples amb relació al pronot (Figs. 1a, b). VIII abdominal de les femelles amb l’apòfisi mitjana d’aspecte diferent (Fig. 2a, b) .........................................5
5 Èlitres convexos respecte al pronot, arrodonits en l’àpex (Fig. 1b). VIII segment abdominal de les femelles amb l’apòfisi mitjana prima i allargada (Fig. 2b) .....................
............................................................ P. cruillensis n. sp. – Èlitres menys respecte el pronot convexos, menys arrodonits en l’àpex (Fig. 1a). VIII segment abdominal de les femelles amb l’apòfisi mitjana curta i ampla (Fig. 2a) P. urgellesi (Español)
Bibliografia
Español, F. 1964. Una interesante localización de Speonomus en calizas paleozoicas de la cordillera litoral catalana. Speleon, 15 (1-4): 49-59.
Bellés, X. & Escolà, O. 1977. Nuevos datos sobre los Speonomus del grupo delarouzeei (Fairmaire, 1860): Parvospeonomus subgen. nov. (Col. Bathysciinae). Speleon, 23: 33-37.
Fresneda, J., 1998. Revisión de los generos de Leptodirinae de la sección Speonomus del Sur de los Pirineos: géneros Antrocharidius Jeannel 1910, Perriniella Jeannel 1910, Speonomus Jeannel 1908, Troglocharinus Reitter 1908 y Troglophyes Abeille 1894. (Coleoptera, Cholevidae). Mémoires de Biospéologie, 25: 53-86.
34 Butlletí ICHN 85 (1), 2021
J. COMAS
Butlletí de la Institució Catalana d’Història Natural, 85 (1): 35-36. 2021
NOTA BREU
ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition)
NOTA BREU
Actualització de la informació sobre la població de Saponaria glutinosa (Caryophyllaceae) a l’Alt Urgell (Catalunya)
New information about Saponaria glutinosa (Caryophyllaceae) in Alt Urgell (Catalonia)
Albert Tarragó*
* Centre de Ciència i Tecnologia Forestal de Catalunya (CTFC). Carretera de Sant Llorenç de Morunys, km 2. 25280 Solsona. Lleida. A/e: albert.tarrago@ gmail.com
Rebut: 03.02.2021. Acceptat: 11.03.2021. Publicat: 31.03.2021
Saponaria glutinosa M. Bieb.
Alt Urgell: Organyà, Congost de Tresponts 556 m, 31TCG67, tartera estabilitzada de roques calcàries, en un marge de la carretera, 28/06/2020, A. Tarragó (Fig. 1, 2 i 3).
Saponaria glutinosa és una planta herbàcia de fins a 50 cm d’alçada, que floreix entre els mesos de maig a juny. Es distribueix pel nord d’Àfrica i el sud d’Europa, i es troba molt dispersa a la península Ibèrica. L’espècie té preferència per zones pedregoses obertes i de forta insolació. L’actual estatus relatiu a l’estat de conservació de l’espècie a Catalunya és EN -En perill d’extinció- (Sáez et al. 2010), categoria assignada sobre la base dels criteris estàndards de la IUCN. L’espècie es troba catalogada com a «en perill d’extinció» en el Catàleg de flora amenaçada de Catalunya: Decret 172/2008, Resolució AAM/732/2015.
La localitat documentada en temps més recents correspon a la Vall de Ca n’Aguilera (Piera, Anoia), en tres nuclis separats. En aquest indret l’espècie creix sobre esquist primaris, a una altitud entre els 500 i 680 m snm en orientació sud (Qua-

drada, 1997). Per altra banda, la presència de S. glutinosa a la Serra del Montsec seria deguda a una confusió amb Saponaria caespitosa DC. (Sáez et al. 2010).


35 Butlletí ICHN 85 (1), 2021
DOI: 10.2436/20.1502.01.70
Figura 1. Detall de les flors de Saponaria glutinosa
Figura 2. Individu en roseta basal en el període vegetatiu previ a la floració.
Figura 3. Zona de la carretera C-14 on ha estat eliminada la població de S. glutinosa per efectes de l’ampliació de la calçada.
L’altra localitat catalana coneguda de S. glutinosa es situa prop d’Organyà, on va ser trobada inicialment per Soulié el 1908, que la situava en pedruscalls calcaris de Tres Ponts (Vogin, 2018). Posteriorment, l’any 1974, P. Montserrat la va situar sobre unes tarteres de les gorgues del riu Segre prop d’Organyà (Erviti, 1978).
La població redescoberta de S. glutinosa a Organyà està situada a l’entrada sud del congost de Tresponts. Les plantes d’aquesta localitat creixen en una tartera calcària estabilitzada a peu de carretera. Inicialment (28/06/2020) es van comptabilitzar 7 individus, 4 en floració i 3 vegetatius. Es van prospectar les zones properes però no es van trobar més individus. Aquesta població es trobava en una situació de màxima vulnerabilitat per la proximitat a la carretera C-14. Cal afegir, que el congost de Trespont es troba en obres per la construcció de diversos túnels amb l’objectiu de millorar el traçat de la via actual. Degut als moviments de terra efectuats, no era descartable que una part de la població pogués ser eliminada. Un dels exemplars en floració havia estat escapçat pels treballs de neteja del voral i estava generant una nova tija floral des dels brots basals. En la mateixa zona es va poder observar un petit ramat de cabres que arribaven fins a les mateixes plantes. Tot i que no es van observar indicis d’herbivorisme per part dels ungulats, la seua presència en la zona pot posar en risc la germinació de plantes per efecte del trepig. Per altra banda, la presència dels ungulats en la zona podria ser un possible vector de dispersió de l’espècie que caldria estudiar amb més profunditat.
En una visita posterior (19/02/2021) la zona, s’ha constatat que les plantes observades al juny de 2020 ha estat completament destruïdes per les obres d’ampliació de la calçada
de la carretera C-14 (Fig. 3). Algunes de les plantes han estat eliminades pel raspat de la maquinària al talús de la carretera i les altres han estat cobertes de terra. En aquest nova visita es van prospectar novament les tarteres properes i en una d’elles es van trobar tres peus de S. glutinosa, en una zona propera més al sud del nucli destruït. Aquests individus es troben sobre una tartera calcària en un dels seus laterals amb materials de menor grandària, major acumulació de sòl i vegetació. No es descarta que puguin aparèixer més peus propers a la nova zona corresponent a noves germinacions.
La destrucció del nucli proper a la carretera C-14 per efecte de les obres de millora de la via comprometen la viabilitat de la població de S. glutinosa d’Organyà. A causa de la raresa de l’espècie a Catalunya, el nucli proper a la carretera destruït, el baix nombre d’individus existents i els actuals i futurs factors de risc derivats de la carretera; es considera que s’haurien de prendre mesures immediates per a la conservació d’aquesta població.
Bibliografia
Erviti, J. 1978. Notas de flora navarra, Saponaria glutinosa Bieb. y otras especies interesantes. Munibe (Ciencias Naturales), 4: 249-256. Quadrada, R. V. 1997. Saponaria glutinosa Bieb. i Opuntia vulgaris Mill. a l’Anoia (Barcelona). Butlletí de la Instutució Catalana d’Història Natural, 65: 44.
Sáez, L., Aymerich, P., & Blanché, C. 2010. Llibre vermell de les plantes vasculars endèmiques i amenaçades de Catalunya Argania editio. Barcelona. 811 p.
Vogin, P. 2018. Les pérégrinations botaniques de l’abbé Soulié (1868-1930). Universidade Estadual Paulista «Júlio de Mesquita Filho». 436 p.
36 Butlletí ICHN 85 (1), 2021
TARRAGÓ
NOTA BREU A.
Butlletí de la Institució Catalana d’Història
NOTA BREU
Sclerodermus versus Scleroderma (Hymenoptera: Bethylidae)
Sclerodermus vs. Scleroderma (Hymenoptera: Bethylidae)
Juli Pujade-Villar*, Amador Viñolas**, Josep Muñoz-Batet**, Denis J. Brothers*** & Celso O. Azevedo****
* Universitat de Barcelona. Facultat de Biologia. Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals. Avda. Diagonal, 645. 08028 Barcelona, Catalonia.
** Museu de Ciències Naturals de Barcelona. Laboratori de Natura. Coŀlecció d’artròpodes. Passeig Picasso, s/n. 08003 Barcelona, Catalonia.
*** School of Life Sciences. University of KwaZulu-Natal, Pietermaritzburg. Private Bag X01. Scottsville. 3209 South Africa.
**** Universidade Federal do Espírito Santo. Departamento de Ciências Biológicas. Av. Fernando Ferrari, 514, Goiabeiras, 29.075 910, Vitória, ES, Brazil.
Corresponding author: Juli Pujade-Villar: A/e: jpujade@ub.edu
Rebut: 04.08.2020; Acceptat: 20.03.2021; Publicat: 31.03.2021 urn:lsid:zoobank.org:pub:84B0DB4B-62D2-4DBB-B6F8-8EB8D20BE41D
Bethylidae comprise 96 valid genera (Azevedo et al., 2018). One of these genera has been written with two different spellings, Sclerodermus and Scleroderma. The former is ambiguously attributed to Latreille (1809) or Klug (1809) and the latter to Westwood (1839). Since the description of the genus, several authors have referred to it as Scleroderma (e.g., Berland, 1928; Maneval, 1932; García del Cid, 1941; Bernard & Jacquemin, 1948; Lahourcade, 1963; Evans, 1964; Dauphin & Castro, 1991; Jeremias et al., 2005; Lembo et al., 2008; Fuentes et al., 2009, and see below), while others have referred to it as Sclerodermus (e.g., Gordh & Móczár, 1990; Pérez-Irezábal, 2003; Papini, 2014; Jirón et al., 2016; Karimi et al., 2017; Skvarla, 2018; Verdugo & Tussac, 2001; Azevedo et al., 2018; Rond, 2019, and see below). However, this ambiguous usage is wrong and one spelling must be rejected.
The main goal of this contribution is to check if both names are available and, if so, which one has priority under the scope of the International Code of Zoological Nomenclature (ICZN, 1999).
The history begins with Volume 4 of Latreille’s (1809: 118) «Genera Crustaceorum et Insectorum…», in which he considered that specimens (named by Klug as «Sclerodermus») belonged to the genus Methoca Latreille, 1805 (correctly Methocha Latreille, 1804, according to Agnoli (2005)). The name Sclerodermus was undoubtedly given by Klug, which Latreille (1809) mentioned («Sclerodermus Klug»), but Latreille did not describe it as such since he considered it to be a junior synonym of Methoca, and therefore not a valid name. For this reason (see ICZN, 1999: Article 11.5), the name Sclerodermus is not available from that publication. Latreille (1809: 119) included and differentiated two species in the genus Methoca, M. ichneumonides Latreille (defined as: «Truncus nodosus, articulatus») and «Sclerodermus domesticus, Klug» (defined as: «Truncus elongato-cubicus, supra planus»). These short descriptions provide characters
to differentiate the species. So, the specific name domesticus is accompanied by a description, and is therefore available from Latreille (1809), but as authored by Latreille only (not Klug as Latreille wrote), since Klug had not provided the description or published the name (see ICZN, 1999: Articles 50.1 and 50.1.1). (In this context, we also have to mention Guérin (1839: 630) who was the first author to state that Klug never described «Sclerodermus».) The fact that the generic name was unavailable has no effect on the availability of the species name (ICZN, 1999: Article 11.9.3.1).
In his subsequent one-volume exposition on his rationales for developing a natural system of the crustaceans, arachnids and insects, and a review of their genera («Considérations Générales sur l’Ordre Naturel des Animaux…»), Latreille (1810: 314) used the name Sclerodermus as a valid genus and described it for the first time. He also specified its type species as «Sclerodermus domesticus, Klug» (Latreille, 1810: 437), but did not indicate that the genus was one which he had proposed himself (it was not asterisked), presumably because he considered Klug the first to have done so, although Klug had not actually published the name. Surprisingly, this study has apparently gone unnoticed by all authors to date and nobody has used the correct Latreille work to establish the availability of Sclerodermus. The reason it has been overlooked remains a mystery, but perhaps people just erroneously assumed that the work was merely a summarized version of the major «Genera» work. Further confusion resulted from the fact that in the first edition of Cuvier’s «Le Règne Animal…» Latreille (1817: 491) used only the colloquial French version («sclérodermes»), but in the second edition (Latreille, 1829: 316) he used the spelling «Scleroderma» as the formal name and attributed it to Klüg [sic], and in the third edition the spelling was again «Scleroderma» attributed to Klüg (Cuvier, 1836: 191). The first author to use the spelling «Scleroderma» (without any author attribution) was apparently Oken (1817: «1178») in his comparison of his own and Cu-
37 Butlletí ICHN 85 (1), 2021
NOTA BREU
Natural, 85 (1): 37-40. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition)
10.2436/20.1502.01.71
DOI:
vier’s classifications. The change in spelling was presumably to agree with the proper transliteration of the Greek δέρμα (= derma, meaning «skin»), a neuter noun, instead of the irregular masculine latinization, but this change is an incorrect subsequent spelling (ICZN, 1999: Article 33.3) and is not available, although it was subsequently often adopted.
Most significantly, Westwood (1839) used the spelling Scleroderma in his monograph of the genus, although he quoted Latreille (1809: 119) as mentioning «Sclerodermus domesticus» and Latreille (1829) as including «Scleroderma» between Myrmecodes and Methoca. He also stated: «As far as I am aware, no further description either generic or specific has ever been published», apparently being unaware of Latreille’s (1810) treatment. Westwood (1839: 164) gave both spellings as a heading («Scleroderma, Klug. MSS. Sclerodermus, Latreille olim.») and therefore demonstrably intended the change from the original spelling. Scleroderma Westwood, 1839 is, therefore, an «unjustified emendation» (ICZN, 1999: Article 33.2.3) and is a separately available name and an objective junior synonym of Sclerodermus Latreille, 1810.
Lepeletier de Saint Fargeau (1845: 573–574) under «Methoca» considered «Sclerodermus domesticus Klug» to be a synonym of «Methoca domestica Latr.», but, inexplicably, also treated the genus «Schroderma Klug» separately, giving the characters stated by Latreille (1829) for Scleroderma as distinctive. He also commented that, although neither he nor Latreille had seen any species of the genus, it should meanwhile be linked with Methoca. The changed spelling was most likely merely the result of a transcription error (the le having been read as h).
Giraud (1856: 180-181), on the basis of two mating pairs, identified «Methoca domestica» of Latreille ([1809]) (= «Sclerodermus domesticus (Klug)» of Latreille (1810)) as being the female of «Bethylus depressus» of Fabricius ([1804]), and to be the female of the type species of the genus Pristocera Klug, [1808], but did not comment on the spelling of Sclerodermus
Even in the mid-20th century, Lahourcade (1963: 849) commented that the description of the genus «Scleroderma» was merely outlined by Latreille at the beginning of the previous century, and was not explicitly detailed until Westwood’s (1839) work. He did not mention the date of Latreille’s description and did not use the original spelling.
In conclusion, Sclerodermus Latreille, 1810 (not 1809) is the oldest available name and its type species is Sclerodermus domesticus Latreille, 1809 (not Klug). Scleroderma Westwood, 1839 is therefore identified as a junior objective synonym of Sclerodermus Latreille, 1910. The names and authorships for the genus published over time by taxonomists are summarized as follows, including only references considered to be of particular significance:
Sclerodermus Latreille, 1809: 118 [nomen nudum, considered erroneously as an available name by different authors]: Bridwell (1919: 26), Gordh & Moczar (1990: 156), Skvarla (2018: 752), Azevedo et al. (2018: 245).
Sclerodermus Latreille, 1810: 314 [σκληρός (= sklēros) hard (adj.) + δέρμα] (= derma, but dermus after irregular latinization) skin (masculine noun); available name] .
Scleroderma Klug, 1809 [incorrect subsequent spelling (unavailable name) and erroneous authorship]: Latreille (1829: 316), Westwood (1839: 164), Guérin (1839: 630), Blanchard (1840: 374), Lucas (1849: 322), Foerster (1856: 96), Walker (1871: 59), Saunders (1881: 109), Westwood (1881: 117), Ashmead (1902: 272), Kieffer (1914: 253), Berland (1928: 124), Bernard & Jacquemin (1948: 160), Nicoli & Erlande-Branderburg (1957: 551).
Schroderma Klug, 1809 [incorrect subsequent spelling (unavailable name) and erroneous authorship]: Lepeletier de Saint Fargeau (1845: 646).
Scleroderma Latreille, 1809 [incorrect subsequent spelling (unavailable name) and erroneous year of descriptor]: Oken, (1817: 1178), Cuvier (1836: 191), Ashmead (1893: 40), Ashmead (1902: 270), Kieffer (1904: 373), Kieffer (1905: 103), Kieffer (1908: 41), Muesebeck & Walkley (1951: 728), Ogloblin (1954: 101), Lahourcade (1963: 849), Evans (1964-65: 173), Yang et al. (2012: 619).
Sclerodermus Klug, 1810 [erroneous authorship]: Blanchard (1840: 375).
Scleroderma Westwood, 1839: 164 [unjustified emendation (neuter noun), available name] objective synonymy.
Sclerochroa Foerster, 1850: 502-503 [unnecessary new name for Scleroderma because of homonymy with Scleroderma Person, 1801, a genus of fungi (available name), synonymy automatic and by Ashmead, 1893: 40].
Ateleopterus Foerster, 1856: 95-96 [Synonymy by Azevedo et al., 2018: 246].
Schleroderma Costa 1864: 131 [incorrect subsequent spelling (unavailable name)].
Neoscleroderma Kieffer, 1905: 106 [Synonymy by Evans, 1964: 174].
Lepidosternopsis Ogloblin, 1954 [«1953»]: 101-102 [Synonymy by Lanes & Azevedo, 2008: 83].
Sclerodermus includes, according to Azevedo et al. (2018), a total of 81 extant species and a single fossil. In 2011 and 2018 Celso Azevedo visited the Museum für Naturkunde in Berlin, where several holotypes of Sclerodermus species are deposited, and studied a specimen with some labels bearing data as given in the original publication for the type of Scleroderma abdominalis Westwood, 1839. The specimen is a male of Pseudisobrachium Kieffer, 1904 (Pristocerinae) according to the morphology of the eyes (Azevedo et al., 2018: 25) a character which differentiates tribes; consequently, the species was transferred to Pseudisobrachium by Azevedo et al. (2018). This is the only specimen in that collection with the labels of S. abdominalis. The original description by Westwood (1839) is specified as of a female and this is confirmed by the features described; the mentioned specimen thus can-
38 Butlletí ICHN 85 (1), 2021
J. PUJADE.VILLAR, A. VIÑOLAS, J. MUÑOZ-BATET, D. J. BROTHERS & C. O. AZEVEDO
NOTA BREU
not possibly be the holotype of this species. Probably, the labels were changed by mistake, or the females are lost in this Museum. For these reasons, this species is transferred back to Sclerodermus, while hoping to find the female (or females) used to describe this species. Pseudisobrachium abdominalis (Westwood, 1839) (correctly abdominale) is hereby transferred back to Sclerodermus as Sclerodermus abdominalis Westwood, 1839, comb. rev., so that the current number of species included in Sclerodermus is 82.
Bibliography
Ashmead, W. H., 1893. A monograph of the North American Proctotrypidae. Bulletin of the United States National Museum, 45: 1-472.
Ashmead, W. H., 1902. Classification of the fossorial, predaceous and parasitic wasps, or the Superfamily Vespoidea. Family XXXII. – Bethylidae. The Canadian Entomologist, 34: 268-273.
Azevedo, C. O., Alencar, I. D. C. C., Ramos, M. S., Barbosa, D. N., Colombo, W. D., Vargas, J. M. R. & Lim, J. 2018. Global guide of the flat wasps (Hymenoptera, Bethylidae). Zootaxa, 4489: 1-294.
Agnoli, G. L. 2005. The genus Methocha in Europe: a discussion on taxonomy, distribution and likely origin of its known species and subspecies (Hymenoptera Tiphiidae Methochinae). Bulletin of Insectology, 58 (1): 35-47.
Berland, L. 1928. Faune de France 19. Hyménoptères vespiformes II. Office Central de Faunistique, Paris, 215 p.
Bernard, F. & Jacquemin, P. 1948. Effets des piqures de Scleroderma (Hyménoptères Bethylidae) et revision des espèces nord-africaines. Bulletin de la Société d’Histoire Naturelle de l’Afrique du Nord, 39: 160-167.
Blanchard, E.,1840. Hyménoptères. Histoire naturelle des Insectes Orthoptères, Névroptères, Hémiptères, Hyménoptères, Lépidoptères et Diptères, Tome Troisième. P. Duménil, Paris. P. 219-415 + 1-7 pls.
Bridwell, J. C. 1919. Some notes on Hawaiian and other Bethylidae (Hymenoptera) with descriptions of new species. Proceedings of the Hawaiian Entomological Society, 4: 21-38.
Costa, A. 1864. Géneri e specie d’Insetti della Fauna Italiana. Annuario del Museo Zoologico della R. Università di Napoli, 2: 128-138, 1 pl.
Cuvier, G. 1836. Le Règne Animal distribué d’après son organisation, pour servir de base à l’histoire naturelle des animaux et d’introduction à l’anatomie comparée, Troisième edition, Tome Troisième. Louis Hauman et Comp. Bruxelles. 524 p.
Dauphin, P. & Castro, J. 1991. Scleroderma abdominalis West, responsable de dermatoses dans le Sud-Ouest de la France (Hym. Bethylidae). L’Entomologiste, 47 (6): 327-328.
Evans, H. E. 1964. A synopsis of the American Bethylidae (Hymenoptera, Aculeata). Bulletin of the Museum of Comparative Zoology, 132: 1-222.
Fabricius, J. C. 1804. Systema Piezatorum secundum ordines, genera, species, adiectis synonymis, locis, observationibus, descriptionibus. Carolum Reichard. Brunsvigae (= Braunschweig). 439 p.
Foerster, A. 1850. Eine Centurie neuer Hymenopteren, Dritte Dekade. Verhandlungen des naturhistorischen Vereines der preussischen Rheinlande und Westphalens, 7: 501-518.
Foerster, A. 1856. Hymenopterologische Studien, II. Heft. Chalcidiae und Proctotrupii. Ernst ter Meer. Aachen. 152 p.
Fuentes, M. V., Sainz-Elipe, S., Sáez-Durán, S. & Galan-Puchades, M. T. 2009. Scleroderma domestica (Hymenoptera, Bethylidae),
woodworm parasitoid and accidental human pathogen. Report of three cases in the city of València (Spain) and its surroundings. Revista Ibero-Latinoamericana de Parasitología, 68 (2): 185-187. García del Cid, F. 1941. Lucha contra los insectos bibliófagos en las Bibliotecas de Cataluña. Anales de la Escuela de Peritos Agrícolas y Superior de Agricultura y de los Servicios Técnicos de Agricultura, 1 (4): 291-309.
Giraud, J. 1856. Observations sur quelques espèces d’Hyménoptères rares ou peu connues trouvées dans les environs de Vienne. Verhandlungen der Zoologisch-Botanischen Gesellschaft in Wien, Abhandlungen, 6: 179-188.
Guérin, F.-E. 1839. SCLÉRODERME, Sclerodermus (Ins.). In: Guérin, F.-E. (Ed.), Dictionnaire pittoresque d’histoire naturelle et des phénomènes de la nature. Tome Huitième. P. 630.
Gordh, G. & Móczár, L. 1990. A catalog of the world Bethylidae (Hymenoptera: Aculeata). Memoirs of the American Entomological Institute, 46: 1-364.
ICZN (International Commission on Zoological Nomenclature), 1999. International Code of Zoological Nomenclature, Fourth Edition. International Trust for Zoological Nomenclature. London. 335 p.
Jirón, L. F., Hanson, P. & Zeledón, R. 2016. Un brote de dermatitis producido por picaduras de Sclerodermus sp. (Hymenoptera, Bethylidae) en Costa Rica, América Central. Acta Médica Costarricense, 58 (3): 122-125.
Jeremias, J., Escolà, O. & Pérez De-Gregorio, J. J. 2005. Scleroderma domestica Latreille, 1809 (Hymenoptera, Bethylidae) un patógeno accidental no parásito para el hombre. Sessio Conjunta de Entomologia ICHN-SCL, 13 (2003): 61-66.
Karimi, J., Darsouei, R. & Sharifi, S. 2017. A bethylid wasp (Hymenoptera: Bethylidae) as a promising biocontrol agent of rosaceous long horn beetle Ospheranteria coerulescens (Coleoptera: Cerambycidae). Entomological News , 127 (2): 123-132.
Kieffer, J. J. 1904. Description de nouveaux Dryininae et Bethylinae du Musée Civique de Gênes. Annali del Museo Civico di Storia Naturale di Genova, 41: 351-412.
Kieffer, J. J. 1905. Description de nouveaux Proctotrypides exotiques avec une planche et une figure dans le texte. Annales de la Société Scientifique de Bruxelles, 29: 95-142.
Kieffer, J. J. 1908. Bethylidae. In: Wytsman, P. (Ed.), Genera Insectorum, 76: 1-50.
Kieffer, J. J. 1914. Bethylidae. Das Tierreich, 41: 228-595.
Klug, F. 1808. Über die Geschlechtsverschiedenheit der Piezaten, andere Hälfte der Fabriciusschen Gattungen. Der Gesellschaft naturforschender Freunde zu Berlin, Magazin für die neuesten Entdeckungen in der gesammten Naturkunde, 2: 48-63.
Lahourcade, M. 1963. Quelques précisions sur la morphologie et la biologie de Scleroderma domestica Latr. petit Hyménoptère Béthylide vulnérant. Annales de Parasitologie Humaine et Comparée, 37 (5-6): 848-860.
Lanes, G. O. & Azevedo, C. O. 2008. Phylogeny and Taxonomy of Sclerodermini (Hymenoptera, Bethylidae, Epyrinae). Insect Systematics and Evolution, 39: 55-86. https://doi.org/10.1163/ 187631208788784165
Latreille, P. A. 1804. Tableau méthodique des Insectes. Classe huitiéme. Insectes, Insecta. In: Nouveau Dictionnaire d’Histoire Naturelle, appliquée aux Arts, Principalement à l’Agriculture et à l’Economie rurale et domestique. Tome XXIV. Deterville. Paris. P. 129-200.
Latreille, P. A., 1805. Histoire Naturelle, Genérale et particuliere, des Crustacés et des Insectes. Tome treizième. F. Dufart. Paris. 432 p.
Latreille, P. A. 1809. Genera Crustaceorum et Insectorum secundum ordinem naturalem in familias desposita, iconibus exemplisque
39 Butlletí ICHN 85 (1), 2021 NOTA BREU SCLERODERMUS VERSUS SCLERODERMA (HYMENOPTERA: BETHYLIDAE)
J. PUJADE.VILLAR, A. VIÑOLAS, J. MUÑOZ-BATET, D. J. BROTHERS & C. O. AZEVEDO
plurimis explicata. Vol. 4. Armand Konig. Parisiis et Argentorati (Paris & Strasbourg). 399 p.
Latreille, P.A., 1810. Considérations générales sur l’ordre naturel des animaux composant les classes des crustacés, des arachnides, et des insectes; avec un tableau méthodique de leurs genres, disposés en familles. F. Schoell. Paris. 444 p.
Latreille, [P. A.] 1817. Les Crustacés, les Arachnides et les Insectes. Cuvier, [G.], (Ed.), Le Règne Animal, distribué d’apres son organisation, pour servir de base à l’histoire naturelle des animaux et d’introduction a l’anatomie comparée. Tome III, Deterville. Paris. 682 p.
Latreille, [P. A.] 1829. Suite et Fin des Insectes. Cuvier, [G.], (Ed.), Le Règne Animal, distribue d’après son organisation, pour servir de base a l’histoire naturelle des animaux et d’introduction a l’anatomie compare. Nouvelle Édition, revue et augmentée. Tome V. Deterville. Paris. 580 p + 20 plate.
Lembo, S., Panariello, L., d’Errico, F. P. & Lembo, G. 2008. Professional’s and non-professional’s papular urticaria caused by Scleroderma domesticum Contact Dermatitis, 58: 58-59.
Lepeletier de Saint Fargeau, A. 1845. Histoire naturelle des Insectes. Hyménoptères. Tome Troisième. Librairie Encyclopédique de Roret. Paris. 646 p.
Lucas, H. 1849. Histoire naturelle des animaux articulés. Troisième partie - Insectes (Suite). Exploration scientifique de l’Algérie pendant les années 1840, 1841, 1842. Sciences physiques. Zoologie. III. Arhis Bertrand. Paris. 532 p.
Maneval, H. 1932. Notes recueillies sur les Hyménoptères. Annales de la Société entomologique de France, 101: 85-110.
Muesebeck, C. F. W. & Walkley, L. M. 1951. Family Bethylidae. In: Muesebeck, C. F. W., Krombein, K. V. & Townes, H. K. (Eds.), Hymenoptera of America North of Mexico: Synoptic Catalog. United States Department of Agriculture Agriculture Monograph, 2: 726-734.
Nicoli, R. M. & Erlande-Brandenburg, G. 1957. L’envenimation par piqure de Scleroderma (Hymenoptera, Bethylidae). Annales de Parasitologie Humaine et Comparee, 32: 551-562.
Ogloblin, A. A. 1954 . Los insectos de las islas Juan Fernandez. 14. Bethylidae y Dryinidae. Revista Chilena de Entomologia, 3 (1953): 101-115.
Oken, L. 1817. Cuviers und Okens Zoologien neben einander gestellt. Isis oder Encyclopaedische Zeitungvon Oken, 1 (8): 1145-1185.
Papini, R. A. 2014. A case of stings in humans caused by Sclerodermus sp. in Italy. Journal of Venomous Animals and Toxins including Tropical Diseases, 20: 1-5.
Pérez-Irezábal, J., Gardeazabal, J. & Saloña, M. 2003. Urticaria recidivante de dos años de evolución. Enfermedades Infecciosas y Microbiología Clínica, 21 (7): 381-382.
Rond, J. de, 2019. First record of Bethylidae (Hymenoptera) from Malta. Bulletin of the Entomological Society of Malta , 10: 108-109.
Saunders, S. S. 1881. On the habits and affinities of the Hymenopterous genus Scleroderma , with descr£ptions of new species. Transactions of the Entomological Society of London , 1881: 119-116.
Skvarla, J. M. 2018. A Review of Sclerodermus Latreille, 1809 (Hymenoptera: Bethylidae) infestations and report of the first case in North America North of Mexico. Journal of Medical Entomology, XX (X), 2018, 1-5. doi: 10.1093/jme/tjx238
Verdugo, A. & Tussac, H. 2001. Sclerodermus abdominalis Westwood, 1839 (Hymenoptera, Bethylidae) primer estoparásito conocido de Sphenoptera pilosula gadetinica Verdugo y Coello, 2001 (Coleoptera, Buprestidae). Boletín de la Sociedad Entomologíca Aragonesa, 29: 65-67.
Walker, F. 1871. Notes on Chalcidiae. Part IV. Chalcididae, Leucospidae, Agaonidae, Eucharidae, Perilampidae, Ormyridae, Encyrtidae. E. W. Janson. London. P. 55-70.
Westwood, J. O. 1839. Monograph upon the hymenopterous genus Scleroderma Transactions of the Royal Entomological Society of London, 2: 164-172.
Westwood, J. O. 1881. Observations on the hymenopterous genus Scleroderma, Klug, and some allied groups. Transactions of the Entomological Society of London, 1881: 117-140, pls. IV-VII.
Yang, Z.-Q., Wang, X.-Y., Yao, Y-X., Gould, J. R. & Cao, L.-M. 2012. A new species of Sclerodermus (Hymenoptera: Bethylidae) parasitizing Agrilus planipennis (Coleoptera: Buprestidae) from China, with a key to Chinese species in the genus. Annals of the Entomological Society of America, 105 (5): 619-627.
40 Butlletí ICHN 85 (1), 2021
NOTA BREU
Notes florístiques de les conques altes dels rius Segre i Llobregat. V
Pere Aymerich Boixader*
* C. Barcelona, 29. 08600 Berga. A/e: pere_aymerich@yahoo.es
Rebut: 17.01.2021; Acceptat: 10.04.2021; Publicat: 30.06.2021
Resum
S’aporten noves dades per a la flora regional de les conques superiors dels rius Segre i Llobregat (Pirineus orientals, Catalunya). Destaquem el conjunt de localitats que precisen la distribució de l’endemisme d’àrea restringida i mal conegut Alchemilla cadinensis. Una part notable de les citacions correspon a plantes introduïdes, al·lòctones al conjunt de Catalunya o bé a la zona d’estudi, de les quals destaquen les primeres observacions en aquesta àrea de la invasora Impatiens glandulifera.
Paraules clau: flora, corologia, Pirineus, Catalunya, NE península Ibèrica.
Abstract
Floristic data of the upper Segre and Llobregat basins. V
This work provide new data about the regional flora of the upper Segre and Llobregat basins (Eastern Pyrenees, Catalonia). We highlight the localities that contribute to knowledge of the distribution of the narrow endemic species Alchemilla cadinensis. Many data belong to introduced plants, which are alien in the whole Catalonia or only in the study area, including the first reports in this area of the highly invasive Impatiens glandulifera.
Key words: flora, chorology, alien species, Pyrenees, Catalonia, north-eastern Iberian Peninsula. Butlletí
Introducció
Aquest article continua treballs previs, en particular quatre de títol homònim (Aymerich, 2003, 2014, 2015b, 2016b), que tenen com a objectiu millorar el coneixement de la flora de la zona que abasten les conques altes dels rius Segre i Llobregat, situada en l’àmbit pirinenc i subpirinenc del nord de Catalunya. Es tracta d’informacions diverses, que es refereixen a novetats florístiques regionals o a tàxons que són rars o mal coneguts tant a escala regional com catalana.
Material i mètodes
Com als treballs previs d’aquesta sèrie, l’àmbit geogràfic considerat és una zona heterogènia del nord de Catalunya amb una extensió d’uns 2000 km2, que comprèn les conques altes dels rius Segre (amb límit sud a l’embassament de Rialb) i Llobregat (amb límit sud al baix Berguedà i zones adjacents del Bages i el Solsonès).
Les citacions de cada tàxon indiquen, en aquest ordre: comarca, municipi, indret, quadrat UTM 1 × 1 (sempre dins el fus 31T i amb sistema de referència ETRS89), altitud, hàbitat i data d’observació (si s’han fet diverses observacions al lloc
en un mateix període, només indiquem el mes o l’any). Els topònims de referència utilitzats són els de la base cartogràfica de l’Institut Cartogràfic de Catalunya (http://www.icgc. cat), amb el nivell de detall considerat més adequat en cada cas. Les citacions són complementades amb comentaris diversos que tenen com a objecte contextualitzar la dada.
Les referències florístiques principals utilitzades per avaluar l’interès relatiu de les dades en el context de Catalunya han estat els diversos volums de l’atles florístic ORCA, les síntesis florístiques modernes (Bolòs & Vigo, 1984-2001; Bolòs et al., 2005) i dues bases de dades corològiques en línia (Base de Dades de Biodiversitat de Catalunya: http://biodiver.bio.ub.es/biocat/; Atles de la Flora dels Pirineus: www. florapyrenaea.com o www.atlasflorapyrenaea.eu/). Aquestes bases de dades es citen dins el text de forma abreviada: la Base de Dades de Biodiversitat de Catalunya com a BDBC i l’Atles de la Flora dels Pirineus com a AFP. Les consultes a BDBC i AFP han estat actualitzades a l’hivern de 2021.
Les citacions es presenten agrupades en dos blocs, segons l’origen de les plantes al territori considerat. El primer inclou tàxons que es consideren autòctons a escala regional. Al segon bloc s’exposen els tàxons aŀlòctons a Catalunya i també aquells que, tot i ser autòctons en alguna part de Catalunya, han arribat a l’àmbit d’estudi per via antròpica
41 Butlletí ICHN 85 (2), 2021 GEA, FLORA ET FAUNA GEA, FLORA ET FAUNA
de la
d’Història Natural, 85 (2): 41-53. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition) DOI: 10.2436/20.1502.01.72
Institució Catalana
GEA, FLORA ET FAUNA
Tàxons autòctons
Achillea ceretanica (Sennen) I. Soriano
Berguedà: Bagà, entre coll de Pal i Comafloriu, DG1084, 2180 m, pastura acidòfila, 17-VIII-2020.
Primera observació al Berguedà d’aquest endemisme escàs de la meitat est dels Pirineus, amb àrea centrada a la Cerdanya, i que també ha estat trobat localment a l’Alt Urgell i a Andorra (Soriano, 2019). Tot i l’ampliació administrativa, aquesta nova localitat no representa una novetat important en la seva àrea de distribució, perquè A. ceretanica ja era coneguda d’alguns punts de la vall de la Molina (Cerdanya) situats a pocs quilòmetres. A Bagà se’n va veure una sola taca clonal, cosa que fa pensar en una arribada recent, potser derivada del transport de llavors pel bestiar.
Aira caryophyllea L.
Berguedà: Saldes, camp de futbol, CG9575, 1175 m, sobre sorres dipositades artificialment en un camp de futbol poc utilitzat, 23-V-2019.
Aquesta espècie no havia estat citada en l’àmbit del Cadí i les serres prepirinenques pròximes (Vigo et al., 2003). A la localitat de Saldes hi havia una població abundant (centenars d’individus) limitada a un hàbitat artificial i en un entorn en el qual són absents els terrenys silicis en què habitualment es troba aquesta planta. Desconeixem si hi va arribar per via antròpica transportada amb la sorra o si va colonitzar espontàniament el lloc més endavant, però la segona opció sembla menys probable perquè les localitats conegudes més pròximes se situen a desenes de quilòmetres.
Aira cupaniana Guss.
Berguedà: Sagàs, bosc de la Tor, DG1150, 600 m, sòls sorrencs sobre gresos, en clarianes de carrascar, 12-V-2017.
Localitat interior i aparentment isolada d’una espècie que a Catalunya és relativament comuna a la franja litoral i prelitoral (BDBC). En aquesta mateixa localitat ja s’havia trobat A. elegantissima Schur, espècie d’ecologia similar però més rara a Catalunya (Aymerich, 2013a).
Alchemilla cadinensis Aymerich & L. Sáez
Berguedà: Bagà, sota el Roc Negre, Escletxa del Gel, DG0983, 1990 m, fons d’una gran esquerda en una zona calcària carstificada, 1-IX-2017; Saldes, Enforcadura del Pedraforca, costat del Pollegó Inferior, CG9376, 2360 m, base d’un cingle calcari orientat al nord, 23-VII-2018; Cerdanya: Alp, puig de la Font Molsa, sobre Coma Pregona, DG0986, 2290 m, peu de rocam calcari orientat al nord, 28-VII-2017; Alp, torrent de les Set Fonts, DG1086, 2090 m, base de cingle calcari orientat al nord, 16-VII-2019; Urús, Coma Oriola, costat oest sota els Rasos d’Urús, DG0886, 2330 m, vessant rocós amb vegetació quionòfila (Salix pyrenaica, Salix retusa, Antennaria carpatica,...), 22-VI-2017.
Ampliem les localitats conegudes d’aquest endemisme descrit fa pocs anys (Aymerich & Sáez, 2015a), que fins ara són vuit i es concentren al massís de la Tosa d’Alp. Es con-
firma la seva presència al Pedraforca, on ja s’havia considerat probable (Aymerich & Sáez, 2015a) arran d’una indicació poc creïble d’A. fissa Günther & Schummel (Vigo et al., 2003). La localitat de Coma Oriola és possible que sigui la mateixa ja citada en l’article de descripció de l’espècie, basada en un plec d’herbari i de la qual es desconeixien detalls sobre la seva localització i característiques.
Allium scaberrimum J. Serres [A. pardoi Loscos]
Berguedà: Montmajor, al costat de Can Sabata de Dalt, CG9551, 815 m, fenassar en un marge de camp, 4-VII-2015.
Tàxon mal conegut a Catalunya, tot i que segurament rar. En aquesta nota adoptem el criteri de Molina et al. (2018), que consideren A. scaberrimum una espècie mediterrània occidental (europea i africana) d’àrea fragmentada, que inclou A. scaberrimum en sentit estricte, tradicionalment considerat un endemisme provençal, i A. pardoi , durant anys considerat un endemisme de la depressió de l’Ebre. Dins l’àmbit d’aquest article havia estat citat només de prop de la Seu d’Urgell (Molero et al., 2006), mentre que a la resta de Catalunya se’n coneixen unes poques localitats més al Matarranya, les Garrigues, el Priorat, el Pallars Jussà i el Rosselló (Molero et al., 2006; Lewin, 2018).
Arabis auriculata Lam.
alt urgell: Alàs i Cerc, al sud de la Molina de Lletó, CG8184, 1350 m, clarianes de matollar, en terreny rocós silici, 24-V-2018; Alàs i Cerc, barranc de la Fou, CG7984, 1340 m, sòl rocós en un aflorament de roques volcàniques, 10-VI2018; Alàs i Cerc, les Colladetes, CG7684, 1320 m, sòl rocós en un aflorament de roques volcàniques, 10-VI-2018; Josa i Tuixén, a l’oest de Josa, CG8679, 1380 m, clarianes de matollar, en terreny rocós calcari, 21-V-2018; Cerdanya: Bellver de Cerdanya, vall de l’Ingla, pla de l’Ingla, CG9986, 1270 m, matollars en un marge de prat, 29-IV-2020; Bellver de Cerdanya, Santa Eugènia de Nerellà, a l’oest de Cal Quelet, CG9490, 1085 m, clariana de boixeda, en terreny rocós calcari, 29-IV-2020; Urús, Roca d’Urús, DG0687, 1450-1520 m, sòls prims sobre roca calcària, 3-VII-2020..
Espècie no citada fins ara de l’àmbit de la serra del Cadí (cf. Vigo et al., 2003), en el qual es situen totes les localitats indicades. Sí que es coneixia als solells de l’Alt Urgell i la Cerdanya situats al nord del Segre, i també a l’alt Solsonès (BDBC).
Arabis nova Vill.
alt urgell: Alàs i Cerc, la Molina de Lletó, CG8185, 1240 m, bardissa al costat d’un camí, 24-V-2018; Josa i Tuixén, a l’est de Josa, CG8679, 1360-1380 m, bardisses en llocs freqüentats pel bestiar, 21-V-2018; Berguedà: Castellar del Riu, serrat de la Figuerassa, DG0264, 1440 m, matollar de Buxus sempervirens cremat, 20-VI-2013; Cerdanya: Guils de Cerdanya, serra dels Fills, DG0899, 1315 m, matollar nitrificat, en substrat silici, 5-VI-2018; Montellà i Martinet, Estana, a l’oest del coll de Pallers, CG8985, 1470 m, bardissa al costat d’un camí, 1-VI-2017.
42 Butlletí ICHN 85 (2), 2021
P. AYMERICH BOIXADER
Localitats que precisen la distribució regional d’aquesta espècie, escassa i localitzada. Les citacions prèvies provenien en general del sector entre la serra del Cadí sud-oriental i el massís de la Tosa d’Alp (Vigo et al., 2003; BDBC, AFP).
Asphodelus cerasiferus J. Gay
alt urgell: el Pont de Bar, Roca Sombra, CG8492, 11001200 m, relleixos de cingles calcaris orientats al sud, 4-V2017.
Aportem una nova localitat d’aquesta espècie a l’alt Segre, després de la confirmació de la seva presència en un indret de la Cerdanya (Aymerich, 2014). Al Pont de Bar feia una població relativament nombrosa en un solell càlid, en què es troben també altres plantes típicament mediterrànies.
Cardamine heptaphylla (Vill.) O.E. Schulz
alt urgell: Alàs i Cerc, marges del riu de Vilanova prop de la Molina de Lletó, CG8184-8185, 1190-1270 m, verneda i avellanosa, 4-V-2020.
Planta nemoral molt rara a la conca alta del Segre, on només havia estat citada de les valls de l’Ingla i de Pi, al Cadí oriental (Vigo et al., 2003; BDBC). A l’Alt Urgell tan sols era coneguda de la vall de Santa Magdalena, que geogràficament es situa a la conca de la Noguera Pallaresa (Carreras, 1993).
Carex acutiformis Ehrh.
Cerdanya: Das, Sanavastre, riberal del Segre sota el poble, DG0493, 1045 m, vores de sèquies entre prats de dall, VIII i X-2020.
Espècie molt rara a Catalunya, aparentment amb menys de 20 localitats documentades (BDBC), que es concentren als aiguamolls de terra baixa. A la Cerdanya ja havia estat citada d’una localitat entre Bellver i Martinet (Vigo et al., 2003), que al BDBC es reflecteix per duplicat abastant dos quadrats UTM de 10 × 10 km (CG99 i DG09); l’assignació a DG09 és incorrecta, de manera que la nova citació de Sanavastre és la primera per aquest quadrat. També n’hi ha una citació antiga al Segre a la Seu d’Urgell (Montserrat, 1957).
Carex brevicollis DC.
Cerdanya: Alp, vessant nord de la Tosa d’Alp, torrent de les Set Fonts, DG1086, 2170-2190 m, en un vessant rocós calcari orientat al nord, 16-VII-2019.
Nova localitat d’aquesta espècie molt rara i protegida a Catalunya, que només es coneix amb seguretat al massís de la Tosa d’Alp (Sáez et al., 2010; Soriano & Aymerich, 2017).
Cerastium gracile Dufour
Berguedà: Vilada, riba est del pont de la Baells, DG0764, 640 m, terreny sorrenc en una zona d’aparcament, 3-V-2018.
A Catalunya, aquest teròfit viu bàsicament a l’oest i al sud (BDBC). En coneixem una sola citació prèvia a la conca del Llobregat, a Gironella (Aymerich, 2014), on apareixia molt escàs, mentre que a la Baells feia una població de centenars d’individus.
Colchicum autumnale L.
Cerdanya: Bellver de Cerdanya, vall de l’Ingla, al sud-est de Santa Magdalena, als dos costats del riu de l’Ingla, CG99879988, 1130-1170 m, antics prats de dall, arbredes i bardisses, 29-IV-2020.
Nova localitat d’una espècie que a Catalunya és molt rara -menys de quinze citacions històriques (BDBC, AFP)- i es troba en situació d’extrem meridional d’àrea. Aproximadament la meitat de les citacions correspon a les parts basals de la serra del Cadí a la Cerdanya, com aquesta nova. La localitat de l’Ingla és remarcable per la seva extensió (diversos nuclis poblacionals al llarg de més de 600 m) i població (aparentment uns quants milers d’individus).
Daphne alpina L.
Cerdanya: Urús, Roca d’Urús, DG0687, 1400 m, base d’un cingle calcari, 25-VI-2020; Urús, serrat de les Pedrusques, al nord-oest del Roc de l’Orri, DG0687, 1650-1830 m, rocams calcaris i tarteres semiestabilitzades, 2017-2018.
Planta molt rara a Catalunya (Sáez et al., 2020), de la qual els darrers anys s’han trobat aquestes dues noves poblacions (o subpoblacions) en l’àmbit ampli del serrat de les Pedrusques (massís de la Tosa d’Alp). S’afegeixen a les altres tres localitats conegudes en territori català, dues també a la Cerdanya, més cap a l’oest (Gruber, 1977; Aymerich, 2014) i una a la serra de Prada, Alt Urgell (Guardiola et al., 2016). La localitat pròxima al Roc de l’Orri va ser objecte d’una prospecció promoguda pel Parc Natural del Cadí-Moixeró l’any 2018, amb la qual es van localitzar 35 individus, mentre que a la de la Roca d’Urús se n’han observat una desena, però resta pendent fer-ne una avaluació detallada.
Delphinium montanum DC.
Berguedà: Saldes, Pedraforca, sota el Pollegó Inferior, CG9376, 2350 m, peu de cingle calcari orientat al nord, 23VII-2018; Gósol, Pedraforca, tartera de Gósol, CG9276, 2255-2260 m, tartera calcària orientada al sud, 23-VII-2018.
Es coneixia una sola observació d’aquesta espècie al Pedraforca (Busquets et al., 1983), però no hi havia estat retrobada més endavant i s’havia considerat probable una extinció local. Hi confirmem la seva persistència fins a l’actualitat, bé que en nombre molt escàs, ja que els anys 2019 i 2020 es van censar només 14 i 12 individus (6 i 9 reproductors) en quatre punts, separats fins a un màxim de 0,5 km.
Doronicum pardalianches L.
Cerdanya: Bolvir, ribera del Segre cap al pont del Soler, DG0994, 1080 m, bosc de ribera, 23-IV-2018; Ger, riba esquerra del Segre a l’alçada del molí de Ger, DG0694, 1060 m, bosc de ribera, 23-IV-2019; Isòvol, ribera del Segre cap a les Prades de Segre, DG0593, 1050 m, bosc de ribera, 23-IV2018; Llívia, ribera del Segre, DH1500-1601, 1180-1195 m, bosc de ribera, 26-IV-2018; Puigcerdà, ribera del Segre cap a Sant Marc, DG1296, 1115 m, bosc de ribera, 26-IV-2019; Puigcerdà, ribera del Segre cap a la depuradora, DG1297, 1125 m, bosc de ribera, 26-IV-2019.
43 Butlletí ICHN 85 (2), 2021 GEA, FLORA ET FAUNA NOTES FLORÍSTIQUES DE LES CONQUES ALTES DELS RIUS SEGRE I LLOBREGAT. V
GEA, FLORA ET FAUNA
Espècie molt rara a la Cerdanya, que fins ara havia estat citada de menys de cinc localitats de les muntanyes que envolten la plana del Segre (Vigo et al., 2003; Font & Vigo, 2008; BDBC; AFP). Als boscos de ribera del Segre és escassa i es presenta en forma de grups clonals molt localitzats.
Draba nemorosa L.
alt urgell: Alàs i Cerc, al sud de la Molina de Lletó, CG8184, 1360 m, taques de sòl pertorbat dins prats mesoxeròfils silicícoles, 24-V-2018; Cava, Ansovell, entorn del santuari del Boscalt, CG8385, 1430-1460 m, taques de sòl pertorbat dins prats mesoxeròfils silicícoles, 24-V-2018.
Localitats d’aquesta espècie, en general rara als Pirineus, al sector nord-oest de la serra del Cadí, on no havia estat citada (cf. Vigo et al., 2003). Als dos llocs es trobava de forma molt localitzada, però feia poblacions nombroses, de centenars d’individus.
Erysimum incanum (Kunze) subsp. aurigeranum (Jeanb, & Timb.-Lagr.) Nieto Fel.
alt urgell: Alàs i Cerc, a l’oest de la collada de Lletó, CG7985, 1500 m, vora de camí, 5-VI-2017; Alàs i Cerc, Lletó, CG8086, 1500 m, marges de camins, 5-VI-2017; la Vansa i Fórnols, cortal del Porta, CG7883, 1620 m, vora de camí, 10-VI-2018.
Aportem tres localitats d’aquest subendemisme català, més aviat escàs, al Cadí nord-occidental. No havia estat citat d’aquest sector, tot i que sí que es coneixia en altres punts de l’alt Segre i les muntanyes que l’envolten (Vigo et al., 2003).
Euphorbia hyberna L.
Cerdanya: Puigcerdà, vores del riu Querol entre Saneja i Sant Martí d’Aravó, DG1099, 1140m, bosc de ribera, 10-IV2017.
Les localitats més pròximes que coneixem d’aquesta espècie atlàntica es situen al Capcir i a l’Alta Cerdanya a l’entorn de Font-romeu (Bolòs et al., 1999, AFP). Al bosc de ribera del Querol hi vam observar un sol individu, probablement d’aparició accidental, però en desconeixem la procedència, ja que no sabem que E. hyberna hagi estat citada més amunt a la vall d’aquest riu.
Euphorbia sulcata Loisel.
Berguedà: Casserres, al sud del Soler de Sant Pau, DG0454, 560 m, codina de gresos carbonatats, 6-VI-2018.
Primera dada sobre aquest teròfit al Berguedà, que ja era conegut d’algun indret pròxim del Bages. Considerant que aquesta espècie no sembla excepcional a la plana de Vic (Casas, 1996; BDBC), suposem que al Berguedà està més estesa del que sembla i que no s’havia citat per confusió amb E. exigua L., amb la qual conviu.
Gagea lutea (L.) Ker Gawl.
Cerdanya: Llívia, ribera del Segre, DH1500-1601, 11801195 m, bosc de ribera, 26-IV-2018.
Hem observat poblacions d’aquesta espècie en dos punts de Llívia, en vernedes eutròfiques, el seu hàbitat típic a la Cerdanya (Aymerich, 2016b).
Gagea reverchonii Degen
Cerdanya: Llívia, entre Llívia i Gorguja, al marge dret del riu d’Er, DH1701, 1210 m, taca de vegetació nitròfila dins un prat de dall, 21-IV-2017.
Nova localitat d’aquesta espècie considerada amenaçada a Catalunya (Sáez et al., 2010) i amb protecció legal. L’observació es refereix a un sol individu reproductor. Gairebé totes les poblacions catalanes es troben a la Cerdanya, sobretot en llocs de muntanya del massís de la Tosa d’Alp (Aymerich et al., 2008), però en els darrers anys també ha estat observada en uns pocs punts de la plana, vora el Segre (Aymerich, 2013b).
Himantoglossum hircinum (L.) Spreng.
Berguedà: Berga, part alta del nucli urbà, al sector de les Estaselles, DG0461, 710 m, parterre d’un jardí particular, 17VI-2020.
Orquídia escassa a Catalunya, que es coneix sobretot del sector nord-oriental comprès entre el nord d’Osona (Vidranès), la Garrotxa i l’Alt Empordà, però que també ha estat indicada d’alguna localitat de la Terra Alta i, antigament, de l’Aran (BDBC). La nova citació al Berguedà és isolada de les anteriors i resulta sorprenent per l’hàbitat completament artificial, un jardí urbà envoltat de cases. Segons la propietat de la casa (Maria Faura & Dolors Pedrals, com. pers.), aquesta espècie hi va aparèixer espontàniament uns anys abans i no s’havien transportat terres des d’altres llocs. L’any 2020 s’hi van observar 5 individus reproductors i 4 no reproductors.
Juncus capitatus Weigel
alt urgell: Cava, el Querforadat, a l’oest del coll de Pallers, CG8985, 1480 m, mulladius temporals en substrat silici, 1-VI-2017; Alàs i Cerc, tossal de Lletó, CG7985, 1540 m, mulladius temporals en substrat silici, 5-VI-2017; Alàs i Cerc, Lletó, CG8086, 1480 m, mulladius temporals en substrat silici, 5-VI-2017.
Ampliem les dades en l’àrea d’estudi sobre aquesta espècie, que és rara als Pirineus. La seva presència en l’àmbit de la serra del Cadí nord-occidental va ser detectada per Guardiola et al. (2016).
Juncus pyrenaeus Jeanb. & Timb.-Lagr.
alt urgell: les Valls de Valira, Ras de Conques, CH6500, 1810 m, mulladiu en sòl carbonatat, juntament amb Carex paniculata i altres plantes higròfiles, 17-VI-2019.
Aquest endemisme de la meitat est dels Pirineus i del Sistema Ibèric és rar a Catalunya i hi està protegit legalment. En l’àmbit considerat, només es coneixien dues localitats prèvies, també a l’Alt Urgell: molleres de Pradell a l’extrem occidental de la serra del Cadí (Vigo et al., 2003) i mollera de Pratprimer poc al sud del port del Cantó (Aymerich, 2012a).
44 Butlletí ICHN 85 (2), 2021
P. AYMERICH BOIXADER
Juncus tenageia L. f.
alt urgell: Alàs i Cerc, sobre el coll de Vanses, CG7985, 1540 m, pradells terofítics humits sobre roques volcàniques, 5-VI-2017; Alàs i Cerc, coll de Vanses, CG8084, 1485 m, pradells terofítics humits sobre roques volcàniques, 20-VI-2017.
Aquestes localitats són, aparentment, les més occidentals d’aquest jonc a Catalunya, on es localitza sobretot a la zona marítima del nord-est. Ja era coneguda de la plana cerdana i el seu entorn (BDBC, dades pròpies), però no de la serra del Cadí.
Legousia scabra (Lowe) Gamisans
alt urgell: Alàs i Cerc, a l’oest de la collada de Lletó, CG7985, 1500 m, vora de camí amb herbes anuals, en substrat silici, 5-VI-2017.
Espècie molt rara en l’àmbit d’estudi i, en general, als Pirineus. Havia estat indicada de dues localitats, a Alp, Cerdanya (Vigo et al., 2003) i Sant Julià de Lòria, Andorra (BDBC, AFP).
Lythrum thymifolia L.
Berguedà: Sagàs, vora Ca l’Estevet, DG1454, 675 m, bassa de nivell fluctuant, 9-VI-2018.
Espècie en general escassa i localitzada a Catalunya, que ha estat citada sobretot de l’Alt Empordà i del sector entre el marge sud del territori sicòric i les muntanyes de Prades (BDBC). La localitat més pròxima coneguda a aquesta nova, també isolada, es troba al límit meridional de l’altiplà del Moianès (Mercadé, 2016).
Minuartia cymifera (Rouy & Foucaud) Graebn.
Berguedà: Guardiola de Berguedà, plans de Canells, sota coll de Pal, DG1082, 1900 m, prat calcícola en sòl pedregós, 12VIII-2018; Gósol, part baixa de la tartera occidental del Pedraforca, CG9177-9277, 1930-2000 m, pedrusques calcàries, 19-VIII-2019; Saldes, tartera oriental del Pedraforca, CG9376, 1950 m, sòl pedregós calcari, 23-VII-2018; Cerdanya: Alp, torrent de Set Fonts, DG1086, 1960-2050 m, pradells pedregosos i marges de camins, 15-IX-2020; Das, entre Coma Oriola i Coma Pregona, DG0887, 2020 m, sòl pedregós calcari, 21VII-2017; Das, vessant nord de la Tosa d’Alp, DG0986, 2440 m, marge d’una pista de servei de l’estació d’esquí, 17-VIII2020; Montellà i Martinet, pas del Grau, vora Prat d’Aguiló, CG9483, 1960 m, marge de pista forestal, 10-VIII-2020; Urús, sota el cim de la Tosa d’Alp, DG0885, 2520 m, sòls pedregosos remenats per obres, 17-VIII-2020.
Espècie que havia estat força inadvertida i és aparentment escassa als Pirineus orientals calcaris. Fins ara havia estat indicada d’uns pocs punts de les serres prepirinenques situades al sud de l’eix Cadí-Moxeró (Vigo et al., 2003; Aymerich et al., 2008; Aymerich, 2015b).
Myriophyllum spicatum L.
Cerdanya: Bellver de Cerdanya, ribera del Segre a Gallissà, CG9791, 995 m, bassa artificial al costat del riu, 5-XII-2019.
Espècie molt rara a l’àmbit considerat i que a la Cerdanya només era coneguda de la bassa de Sanavastre, Das (González & Nuet, 2008; Aymerich et al., 2014). La colonització d’aquesta bassa de Bellver –creada fa uns 20 anys- és molt recent, perquè l’hem visitada de forma anual i fins a 2019 no hi hem vist aquesta planta aquàtica; aquest any ja feia una població abundant que ocupava bona part de la superfície.
Narcissus poeticus L.
alt urgell: Alàs i Cerc, al sud de la collada de Lletó, CG7985, 1510 m, prat mesòfil, 24-V-2018; la Vansa i Fórnols, cortal del Porta, CG7883, 1610 m, antic prat de dall abandonat, 10-VI-2018; Cerdanya: Bellver de Cerdanya, vall de l’Ingla, al sud-est de Santa Magdalena, sobretot al costat est del riu de l’Ingla, CG9987, 1160 m, antic prat de dall abandonat, 29-IV-2020.
Espècie no citada fins ara en l’àmbit de la serra del Cadí (cf. Vigo et al., 2003). A la localitat de la vall de l’Ingla feia una població important, de centenars d’individus, mentre que als dos llocs de l’Alt Urgell era molt escassa (només quatre plantes a la primera localitat i una a la segona).
Onobrychis conferta (Desf.) Desv. subsp. hispanica (Sirj.)
Guitt. & Kerguelen [O. argentea Boiss. subsp. hispanica (Sirj.) P.W. Ball]
alt urgell: la Vansa i Fórnols, entre el coll de Bancs i Fórnols, CG7677, 980 m, talús de carretera orientat al sud, 5-VI2020; SolSonèS: Guixers, marges de la carretera de Sant Llorenç de Morunys a Berga, CG8665-8765-8864-8964, CG9064-9363, 820-930 m, prats secs i matollars clars en talussos de carretera orientats al sud, sobretot en llocs de sòl margós-guixenc i amb coberta arbòria clara de Pinus nigra, 17-VI-2018.
Novetat remarcable per als Prepirineus orientals, amb un conjunt de localitats que marquen l’extrem nord-est fins ara conegut d’aquesta espècie a la península Ibèrica. A Catalunya, és poc freqüent i ha estat citada sobretot de les serres prepirinenques centrals (BDBC, AFP). Les localitats més pròximes referenciades a la bibliografia i a les bases de dades es troben a l’Alt Urgell: vall de Cabó (Sáez et al., 2008) i Organyà (AFP: plec herbari Jaca, leg. P. Montserrat & L. Villar, alt. 500 m, any 1972). Podem afegir que també l’hem vista recentment a Organyà, en el mateix hàbitat que la dada de 1972 (margues erosionades) però a més altitud: Organyà, Cap del Terme, DG6074, 730 m, 4-VI-2019. Tot i que referim aquestes plantes a la subespècie hispanica, cal precisar que presenten caràcters que no són del tot coincidents amb els de la descripció de Valdés ( 2000) , en particular per tenir l’anvers dels folíols sovint més o menys pilós i no pas glabre.
Petasites paradoxus (Retz.) Baumg.
Cerdanya: Alp, la Molina, conca del torrent de les Set Fonts, DG1086, 1900 m, talús d’una pista de servei de l’estació d’esquí, 23-VI-2020.
Aquesta espècie fa poblacions extenses a la part central del vessant nord de la serra del Cadí (Vigo et al., 2003), on
45 Butlletí ICHN 85 (2), 2021 GEA, FLORA ET FAUNA
NOTES FLORÍSTIQUES DE LES CONQUES ALTES DELS RIUS SEGRE I LLOBREGAT. V
GEA, FLORA ET FAUNA
concentra la seva àrea catalana. En zones relativament pròximes de la serra del Cadí occidental i de la serra del Moixeró s’han citat un parell de petits nuclis associats a pistes forestals (Aymerich, 1998), en l’establiment dels quals podria haver intervingut la dispersió antròpica. Correspon a aquesta mateixa tipologia la nova localitat que aportem, situada al massís de la Tosa d’Alp i formada en la data d’observació per menys de 10 individus.
Physalis alkekengi L.
alt urgell: Organyà, cap a Caselles, CG6074, 695 m, marge de camp, 4-VI-2019.
Aquesta planta gairebé no ha estat indicada a la conca alta del riu Segre. Només en coneixem dues indicacions imprecises, una antiga a la Cerdanya i una altra aparentment moderna a la Seu d’Urgell (cf. BDBC, AFP). Aquí n’aportem una dada concreta i recent, referida a una petita població d’una desena d’individus.
Potamogeton natans L.
alt urgell: Organyà, Cap del Terne, CG6074, 720 m, bassa artificial alimentada amb aigua de la font Bordonera, 4-VI-2019.
Novetat per als Prepirineus centrals i l’Alt Urgell. Aquest hidròfit, en general rar a Catalunya, havia estat citat d’una localitat relativament pròxima del Solsonès (Sáez et al., 2004) -Cambrils, uns 10 km cap al sud-est- i es fa més freqüent al Solsonès oriental i al Berguedà, on està en expansió (Aymerich, 2012b).
Primula elatior (L.) L. subsp. elatior
Cerdanya: Bolvir, vores del riu Querol sota Talltorta, DG0994, 1085 m, bosc de ribera, 27-IV-2019; Ger, riba esquerra del Segre a l’alçada del molí de Ger, DG0694, 1055 m, bosc de ribera, 23-IV-2018; Puigcerdà, diversos punts de les vores del riu Querol des de Ventajola fins a Saneja, DG10951097-1099, 1095-1170 m, bosc de ribera, 8 i 10-IV-2017.
Novetat florística interessant per a la Cerdanya i aquest sector dels Pirineus, d’un tàxon que és molt rar a Catalunya (Bolòs et al., 2001; BDBC). Als Pirineus es troba bàsicament a les zones d’influència atlàntica del vessant nord i de l’extrem oest. A Catalunya és coneguda de l’Aran, on no és rara, i penetra cap al sud de forma molt local a la vall d’Àneu, alt Pallars (Aymerich, 2008). La seva localització a la vall del riu Querol representa, aparentment, una penetració al vessant sud-pirinenc similar a la del Pallars. La seva presència al vessant nord de la serra de l’Albera, a l’extrem est dels Pirineus, indicada per Font (2000) a partir de dades bibliogràfiques i recollida després per bases de dades (BDBC, AFP) és incerta i s’hauria de confirmar. Els nuclis locals de la Cerdanya viuen sempre en vernedes eutròfiques i són poc nombrosos, de menys d’una desena d’individus, amb l’excepció d’un de Puigcerdà a 1190 m on n’hi havia desenes.
Rhinanthus alectorolophus (Scop.) Pollich alt urgell: Josa i Tuixén, Josa de Cadí, al nord i est del poble, CG8579-8679, 1380-1390 m, prats de dall, conreus farratgers i pastures mesòfiles, 5-VI-2020; Berguedà: Gósol, zona agrària al sud i sud-est del nucli urbà, CG8976-
CG9076, 1350-1440 m, marges de prats de dall i de conreus farratgers, 5-VI-2020; Gósol, coll de Josa, CG8878, 1630 m, pastura mesoxeròfila en substrat calcari, 5-VI-2020; Saldes, sector de Xisquer, CG9475, 1270-1280 m, marges de prats de dall i de conreus farratgers, 5-VI-2020; Saldes, el Pla, sota el poble, CG9575, 1170-1180 m, marges de conreus farratgers i de camins, 5-VI-2020.
La presència als Pirineus d’aquesta espècie havia estat dubtosa durant dècades, però s’ha confirmat recentment, al Vallespir, Alta Cerdanya i Pallars Sobirà (Mercadal, 2019; Aymerich, 2020). Prèviament se n’havien fet diverses citacions, que en uns casos s’havien considerat poc fiables i en altres s’havia demostrat que eren errònies. L’única citació d’aquest tipus que en coneixem al territori administratiu de la Catalunya autònoma correspon a Gósol i va ser feta per Vives (1964), qui especificava que aquestes plantes tenien pèls llargs al calze, caràcter distintiu de R. alectorolophus. A la flora regional del Cadí (Vigo et al., 2003) s’esmentava aquesta dada bibliogràfica, però no se’n va poder aclarir la validesa. Per aquest motiu, poc després de descobrir la presència de l’espècie al Pallars, vam decidir buscar-la a la rodalia de Gósol. El resultat va ser positiu, i no va costar gens localitzar R. alectorolophus, ja que certament és freqüent als plans que envolten el poble. A més, vam poder trobar aquesta espècie en una àrea més extensa, des de Josa fins a Saldes.
Sabulina villarii (Balb.) Rchb. [Minuartia villarii (Balb.) Wilczek & Chenevard]
Berguedà: Gósol, Pedraforca, base del Pollegó superior a la tartera de Gósol, CG9277, 2325 m, base de cingle calcari orientat al sud, 19-VIII-2019; Saldes, Pedraforca, canal al sudoest del cim del Calderer, CG9377, 2380 m, roques calcàries orientades al nord-oest, 10-IX-2020..
Espècie de distribució alpina molt rara i protegida a Catalunya, que fins ara només es coneixia del massís de la Tosa d’Alp, Berguedà-Cerdanya (Aymerich et al., 2008; Sáez et al., 2010). El seu descobriment al Pedraforca representa una ampliació notable de l’àrea coneguda. Les poblacions observades al Pedraforca eren petites,de menys de 50 individus a Gósol i només un a Saldes.
Sclerochloa dura (L.) P. Beauv.
Cerdanya: Bellver de Cerdanya, vall de l’Ingla, pla de l’Ingla, CG9986, 1270 m, marge de camí forestal, 29-IV-2020; Bellver de Cerdanya, vall de Pi, prop de la font de l’Agre, CG9785, 1360 m, marge de camí forestal, 29-IV-2020.
Tot i que aquesta petita gramínia es coneix des de fa molt a la Cerdanya (Bolòs & Vigo, 2001), on té gairebé totes les seves poblacions catalanes, el nombre de localitzacions concretes fins ara recollides a les bases de dades és molt baix, només vuit (BDBC, AFP). N’aportem dues més, a les valls del vessant nord de la serra del Cadí, sector del qual no havia estat indicada.
Sideritis bubanii Font Quer
alt urgell: Valls de Valira, Sant Joan Fumat, al costat del poble, 990 m, CG7098, herbassars secs sobre carretera, 17-
46 Butlletí ICHN 85 (2), 2021
P. AYMERICH BOIXADER
NOTES
VI-2019; Valls de Valira, sobre Asnurri, CH700, 1260 m, herbassars secs sobre la carretera, 17-VI-2019.
Localitats que precisen el límit nord-oest d’aquesta espècie endèmica (cf. Sáez et al., 2010), que es situa a la vall de Sant Joan Fumat, en la qual fins ara només havia estat citada de la seva part més baixa (Aymerich, 2003).
Sisymbrium macroloma Pomel
Cerdanya: Urús, Roca d’Urús, DG0687, 1450 m, base de cingle, 3-VII-2020.
Nova localitat d’una planta molt rara a Catalunya, coneguda d’una desena de punts dels Pirineus i de les serres meridionals (BDBC). A la Cerdanya havia estat citada només d’un indret del serrat de les Esposes, més cap a l’oest (Aymerich, 2014), i també es coneixia de llocs relativament pròxims del vessant berguedà d’aquestes muntanyes (Soriano, 1992). Com és habitual, aquesta població local era petita, d’una desena d’individus reproductors.
Rorippa palustris (L.) Besser
Berguedà: Cercs, embassament de la Baells sota el poble, DG0666, 630 m, fangar a la franja d’inundació temporal de l’embassament, 3-XI-2017.
Indret colonitzat recentment per aquesta espècie, en expansió als embassaments i la xarxa fluvial de la zona d’estudi (Aymerich, 2016b). La tardor de 2017 hi feia un petit nucli poblacional de menys de deu plantes.
Scleranthus polycarpos L.
Berguedà: l’Espunyola, pla de Caus, CG9765, 825 m, sòls esquelètics sobre conglomerats, 9-V-2018.
Aquesta espècie és força comuna a les serres prepirinenques de la zona considerada, però no era coneguda al baix Berguedà. Sí que es coneixia, en canvi, en altituds similars a les serres prelitorals situades al nord del Llobregat (BDBC).
A la localitat de l’Espunyola convivia amb Muscari baeticum Blanca, Ruiz Rejón & Suár.-Sant., una altra espècie que habitualment està limitada a les zones de muntanya.
Sempervivum ×barbulatum Schott [S. arachnoideum L. × S. montanum L.]
Cerdanya: Urús, costat oest de Coma Oriola, DG0886, 2340 m, rocams calcaris, 22-VI-2017; Urús, entre la Tosa d’Alp i el serrat de les Pedrusques, DG0886, 2420 m, rocams calcaris, 22-VI-2017.
Amb l’excepció relativa de S. fauconnettii Reut. (Aymerich & Sáez, 2015b), els híbrids interespecífics de Sempervivum han estat poc documentats als Pirineus, tot i que hi són relativament freqüents i algunes vegades fan poblacions autònomes. Aportem dues localitats de l’híbrid entre S. arachnoideum i S. montanum, en les quals hi havia poblacions de mida petita (taques disperses) i sense presència pròxima de les dues espècies parentals.
Sempervivum ×schottii C.B. Lehm. & Schinttsp. [S. montanum L. × S. tectorum L.]
Cerdanya: Alp, vessant nord de la Tosa d’Alp, torrent de les Set Fonts, DG1086, 2160-2190 m, vessant rocós calcari orientat al nord, 16-VII-2019; Urús, costat oest de Coma Oriola, DG0886, 2330-2350 m, rocams i sòls esquelètics sobre calcària, 22-VI-2017.
Dues localitats d’un altre híbrid interespecífic, que en aquest cas feia poblacions grans (centenars de rosetes) en absència d’una o dues de les espècies parentals. Les dues poblacions es situen en terrenys calcaris, evitats per S. montanum.
Symphytum tuberosum L.
Cerdanya: Fontanals de Cerdanya, ribera del Segre a l’alçada del camp de golf, DG0794, 1065 m, bosc de ribera, 23IV-2019; Isòvol, ribera del Segre cap a les Prades de Segre, DG0593, 1050 m, bosc de ribera, 23-IV-2018; Isòvol, ribera del Segre prop de l’ermita de Quadres, DG0493, 1040 m, bosc de ribera, 25-IV-2019; Puigcerdà, ribera del Segre entre Mas Florença i Sant Marc, DG1296, 1115-1120 m, bosc de ribera, 26-IV-2019;.
Espècie no citada fins ara de la Cerdanya i molt rara a la conca alta del Segre, on només havia estat indicada de comptades localitats de l’Alt Urgell i d’una d’Andorra (Carreras, 1993; Carrillo et al., 2008; BDBC; AFP). A la ribera del Segre apareix en forma de taques clonals molt escasses i isolades.
Vicia parviflora Cav.
alt urgell: el Pla de Sant Tirs, la Parròquia d’Hortó, Cortal del Sants, CG6487, 1140 m, pradell de teròfits en substrat silici, 1-VI-2017.
Espècie rara a les valls dels Pirineus, que a l’Alt Urgell havia estat indicada només, sense localitat precisa, dels voltants de la Seu (Vigo et al., 2003). A l’alt Segre se’n coneix també una citació a Ur, Alta Cerdanya (AFP).
Viola odorata L.
Cerdanya: Llívia, ribera del Segre, DH1500, 1175 m, bosc de ribera, 26-IV-2018.
Confirmem la presència d’aquesta planta a l’alt Segre, on deu ser molt rara i localitzada. En coneixem una sola indicació prèvia dins l’àmbit geogràfic d’aquest article, corresponent al quadrat UTM CG98 (Bolòs et al., 1998), sobre la qual no s’aporten detalls i que caldria verificar, ja que aquesta espècie no va ser acceptada en la síntesi de la flora regional (Vigo et al., 2003).
Tàxons aŀlòctons i dispersats per via antròpica
En aquest apartat es citen els tàxons que es consideren introduïts a l’àmbit territorial considerat. Es tracta de plantes de dos grups: 1) Tàxons aŀlòctons al conjunt de Catalunya; 2) Tàxons autòctons o probablement autòctons en altres zones de Catalunya, però que a l’àrea d’estudi només es coneixen com a dispersats per via antròpica, de forma intencionada o accidental. Aquest segon grup és un element creixent de les
47 Butlletí ICHN 85 (2), 2021 GEA, FLORA ET FAUNA
LLOBREGAT. V
FLORÍSTIQUES DE LES CONQUES ALTES DELS RIUS SEGRE I
GEA, FLORA ET FAUNA
flores locals, derivat de l’increment en les darreres dècades de la mobilitat i dels transports. El seu tractament és complex, perquè sovint aquestes plantes es naturalitzen i poden plantejar dificultats d’interpretació o ser difícils de segregar de la flora nativa a escala regional. Tot i la dificultat que pot representar, considerem interessant documentar en la mesura possible aquestes situacions, per tal de millorar la interpretació de les dinàmiques florístiques a llarg termini. En general, els tàxons autòctons d’altres parts de Catalunya que aquí es citen com a introduïts a escala local, a hores d’ara resten limitats a hàbitats netament antròpics, sobretot marges de carreteres i ambients ruderals.
Achillea roseoalba Ehrend.
Cerdanya: Bellver de Cerdanya, riba esquerra del Segre, cap a Gallissà, CG9891, 1000 m, prat de dall, 8-VI-2020.
Espècie del complex d’A. millefolium procedent dels Alps i aparentment introduïda a Catalunya. S’ha trobat sobretot en prats de dall de l’Alt Urgell (Aymerich & Soriano, 2016), bé que puntualment també ha estat observada al Pallars Sobirà (Aymerich, 2017). Amb aquesta nota ampliem la seva distribució a la Cerdanya. A Bellver era molt abundant en un prat de dall, però absent a les parceŀles del costat, que tenien el mateix tipus d’hàbitat.
Allium porrum L.
alt urgell: la Seu d’Urgell, prop de l’aiguabarreig del Segre i el torrent de les Torres, CG7590, 690 m, herbassar en un marge de camí, 14-VI-2019; Berguedà: Berga, sota el castell de Sant Ferran, DG0462, 760-770 m, talussos solells amb herbassars nitròfils, 8-VI-2019; Gironella, perifèria nord del poble, DG0754, 460 m, vegetació nitròfila en relleixos d’un rocam, 17-II-2020; Puig-reig, Colònia Vidal, DG0744, 390 m, talús amb vegetació nitròfila, 17-II-2020; Aquesta planta introduïda, escapada dels horts, no havia estat citada de forma clara en l’àmbit d’aquest treball. Les poques dades prèvies es refereixen a A. ampeloprasum en sentit ampli, que també podria incloure altres tàxons. A Berga en coneixem una població persistent ben establerta, que l’any 2019 va augmentar de forma sobtada fins a uns quants centenars de peus, com a resultat d’unes cremes realitzades al vessant en què viu.
Asphodelus fistulosus L.
alt urgell: Bassella, Ogern, perifèria oest del poble, CG6253, 465 m, marge de carretera, 3-V-2019; Coll de Nargó, perifèria oest del poble, a l’inici de la carretera cap a Isona, CG6174, 580 m, marge de carretera, 3-V-2019; Oliana, perifèria nord del poble, CG5959, 455 m, marge de carretera, 3-V-2019; Organyà perifèria nord del nucli urbà, CG6275, 545 m, marge de carretera, 3-V-2019; la Seu d’Urgell, Castellciutat, CG7290, 720 m, marge de carretera, 3-V-2019. Berguedà: Bagà, inici de la carretera de coll de Pal, DG0579, 860 m, marge de carretera, 1-VI-2019; Berga, entrada sud des de l’eix viari del Llobregat, DG0561, 635 m, marge de carretera, 7-V-2014; Cercs, prop de la boca sud del túnel de Cercs, DG0665, 720 m, marge
de carretera, 7-V-2014; Cercs, al costat de l’accés a la presa de la Baells, DG0664, 630 m, 11-V-2014; Olvan, pla de Fullà, DG0755, 480 m, marge de carretera, 12-V-2019; Puig-reig, inici de la carretera a Santa Maria de Merlès, DG0747, 420 m, marge de carretera, 25-IV-2014. OSONA: Olost, carretera de Vic a Prats de Lluçanès a l’alçada del poble, DG2548, 560 m, marge de carretera, 12-V-2019.
Fins a temps recents, aquesta espècie tenia caràcter accidental al Berguedà, on només se’n coneixien un parell d’observacions esporàdiques al nord de la comarca (Soriano, 1984). Coneixem menys la situació prèvia a la vall del Segre a l’Alt Urgell, però sembla molt similar, ja que no n’hem trobat cap dada bibliogràfica segura; l’única recollida a les bases de dades (Devis et al., 2001) prové d’un inventari i, pel lloc i hàbitat, és probable que sigui una confusió amb A. cerasiferus. Pel que fa a l’altiplà del Lluçanès, no sabem que hi hagués estat documentada fins ara. En la darrera dècada, però, A. fistulosus s’ha expandit de forma molt clara al territori, establint noves colònies (desenes o centenars d’individus) en diverses localitats. Aquesta expansió s’ha fet aprofitant la xarxa viària principal i els hàbitats antròpics dels marges de carreteres. És un procés que es va començar a detectar en la segona meitat de la dècada del 2000 i s’ha anat accelerant progressivament. Aportem, sense voluntat exhaustiva, diverses localitats amb presència coneguda de l’espècie, per tal de deixar constància d’aquesta dinàmica colonitzadora recent.
Bothriochloa barbinodis (Lag.) Herter
Berguedà: Olvan, Colònia Rosal, DG0658, 485 m, marge de carretera, 1-X-2020.
Gramínia americana que està en expansió seguint la xarxa viària. Les poques dades prèvies a Catalunya, on hauria arribat des del sud de França, corresponen a la franja marítima entre Barcelona i l’Empordà (Pyke, 2010; Verloove & Aymerich, 2020). La nova citació seria la primera en una zona de l’interior de la Catalunya autònoma. S’hi van veure 6 individus, amb espigues en la data d’observació, però un mes més tard es va constatar que havien estat tallats a causa de treballs d’arranjament viari. Recentment (Lewin et al., 2020) ha estat indicada també d’un punt de l’Alta Cerdanya (Catalunya Nord) molt pròxim a la frontera.
Broussonetia papyrifera (L.) Vent.
BageS: Navàs, solell de can Vall, DG0639, 390 m, talús sobre carretera, 2013-2019; Berguedà: Avià, Graugés, al costat de cal Torrentbò, DG0457, 600 m, marges de camp i bardisses, 4-X-2019.
Aquesta espècie introduïda, originària d’Àsia oriental, és freqüent a les comarques marítimes de Catalunya, però molt rara a l’interior (BDBC). Encara no havia estat citada al Berguedà, on s’ha localitzat una població molt nombrosa, de desenes de peus. Al Bages ja havia estat indicada de la meitat sud (Casasayas, 1989).
Bunias orientalis L.
Cerdanya: Prats i Sansor, pla de Tartera, DG0490, 1115-1125 m, camps i marges de camins, 31-V-2019.
48 Butlletí ICHN 85 (2), 2021
P. AYMERICH BOIXADER
Nova dada d’aquesta planta aŀlòctona a Catalunya, que fins ara només havia estat observada en un parell de localitats, també a la Cerdanya (Aymerich, 2014, 2017). Aquesta població estava formada per un centenar d’individus, escampats per una superfície aproximada d’1 ha.
Coronilla valentina (L.) subsp glauca (L.) Blatt. Berguedà: Berga, perifèria urbana sud-est, DG0461, 650 m, erm en una zona semiurbanitzada, 22-X-2018.
Planta aŀlòctona, escapada de jardineria, que és molt freqüent a la franja marítima de Catalunya però gairebé no ha estat citada a les comarques interiors (BDBC). A la localitat berguedana hi vam observar una desena d’individus.
Cytisus scoparius (L.) Link subsp. scoparius
alt urgell: Montferrer i Castellbò, serrat de Carlà a la cruïlla de la carretera de la Seu d’Urgell a Sort i l’accés al poble de Guils del Cantó, CG5989, 1570 m, marge d’una pineda de Pinus sylvestris, 2-IX-2020; Berguedà: Saldes, entorn del camp de futbol, CG9575, 1175 m, herbassars i bosc clar, 23V-2019.
Aquest tàxon és absent de forma natural al territori considerat en aquest article. Aportem dues dades referides a processos de naturalització local més o menys avançats. La seva presència a Saldes és deguda a la plantació en enjardinaments fets al voltant del camp de futbol, i entre els individus plantats n’hem vist 5-8 nascuts espontàniament, sempre a distàncies curtes, però una veritable naturalització sembla improbable perquè es tracta d’una zona calcària. Sobre Guils del Cantó, en canvi, sí que hi ha una petita població naturalitzada (50-100 individus) , però molt concentrada en l’espai. En aquesta segona localitat, en què conviu amb abundants Cytisus oromediterraneus, no hi ha cap indici que s’hi hagin fet plantacions i es desconeix l’origen de les plantes, que podria ser un transport accidental de llavors per vehicles, ja que es troben al costat de la carretera.
Eruca vesicaria (L.) Cav.
Berguedà: Berga, sota el castell de Sant Ferran, DG0462, 770 m, talús en un marge de carretera, 29-III-2017; l’Espunyola, al sud dels Sants Metges, CG9756, 830 m, camp de cereals, 6-V-2018.
Aquesta espècie no havia estat citada fins ara del Berguedà i és rara, en general, a la meitat nord de Catalunya (BDBC). Els dos petits nucli poblacionals observats en aquestes localitats és probable que siguin el resultat d’introduccions antròpiques recents.
Fallopia japonica (Houtt.) Ronse Decr.
Cerdanya: Puigcerdà, ribera del Segre a la Pedragosa, DG1298, 1130 m, talús sobre el riu, 24-IV-2018; Puigcerdà, ribera del Segre entre Mas Florença i Sant Marc, DG1296, 1115 m, bosc de ribera, 22-IV-2016.
Dades sobre nuclis d’aquesta espècie aŀlòctona en ambients de ribera, el primer una petita població i el segon una sola taca clonal. Ja havia estat indicada de Puigcerdà, però
només en ambients suburbans (Casasayas, 1989; Aymerich, 2014), no en hàbitats fluvials en els quals potencialment podria desenvolupar un comportament invasor.
Glebionis coronaria (L.) Spach
Berguedà: Puig-reig, la Prada, a la perifèria sud del nucli urbà, DG0746, 435 m, herbassar ruderal, 8-II-2020.
Espècie molt poc citada a l’interior de Catalunya, on no és autòctona. Les bases de dades (BDBC) en recullen comptades observacions a Osona i al límit Segrià-Baix Cinca, i també a la Llitera, ja dins àmbit administratiu aragonès. Les plantes observades a Puig-reig (una trentena) tenien el seu origen en escapaments des de jardins situats a pocs metres.
Impatiens glandulifera Royle
Cerdanya: Das, Sanavastre, ribera del Segre, DG0493, 1040 m, illeta fluvial amb herbassars higròfils, 24-IX-2020; Puigcerdà, riba dreta del riu Reür, prop de la confluència amb el Segre, DG1298, 1130 m. bosc de ribera, 24-IX-2020.
Primeres citacions d’aquesta espècie al territori considerat en aquest article. Prèviament, a la Catalunya autònoma havia estat indicada de la conca del Ter (Rifà, 2015; Aymerich, 2016a) i del baix Aran (Aymerich, 2019). Tot i que el nombre d’individus observats a la Cerdanya és baix (menys de vint), considerant la separació entre les dues localitats i el gran potencial invasor d’aquesta planta, és previsible una expansió forta per l’alt Segre en els propers anys.
Isatis tinctoria L.
Berguedà: Berga, al sud-oest de la casa de Garreta, sobre el polígon industrial, DG0161, 865 m, marges de camp, 5-V2020.
Primera dada a la Catalunya central d’una població establerta d’aquesta espècie, formada per uns 50 individus. Prèviament, l’any 2008 s’havien observat dos individus isolats en marges viaris als voltants de Berga (Aymerich, 2013), dades reflectides al BDBC i que sobredimensionen la presència d’aquesta planta a la zona. Aquestes observacions no es van repetir en anys posteriors, però una es va fer a menys d’1 km de la població detectada l’any 2020, motiu pel qual no es pot excloure que tinguin alguna relació.
Kalanchoe ×hougthonii D.B. Ward
Berguedà: Berga, nucli urbà, DG0461, 690 m, fissura de paviment al peu d’una paret, 2-XII-2020.
Espècie hibridògena generada en jardineria, actualment freqüent a la franja marítima de Catalunya (Mesquida et al., 2017; Aymerich & Sáez, 2019). La dada que aportem és, aparentment, la primera a l’interior del territori. Es tracta d’una presència anecdòtica,d’un sol individu de viabilitat dubtosa a mitjà termini, però que reflecteix la capacitat d’aquesta planta de viure en zones d’altitud mitjana i relativament fredes, si més no en hàbitats urbans. Val a dir que, en les modelitzacions de nínxol ecològic de Mesquida et al. (2017), la Catalunya central no es considera favorable per a aquest tàxon en l’actualitat, però sí que ha estat indicada com a zona adequa-
49 Butlletí ICHN 85 (2), 2021 GEA, FLORA ET FAUNA
FLORÍSTIQUES DE LES CONQUES ALTES DELS RIUS SEGRE I LLOBREGAT. V
NOTES
GEA, FLORA ET FAUNA
da per a la seva expansió a finals de segle en els escenaris de més increment de la temperatura.
Lotus corniculatus L. var. sativus Hyl. Berguedà: Berga, enllaç Berga-sud des de l’autovia C-16, DG0560, 630 m, talussos viaris i herbassars adjacents, 24VI-2019; Olvan, enllaç de l’autovia C-16 amb la carretera de Prats de Lluçanès, DG0755, 480 m, talussos viaris i herbassars adjacents, 24-VI-2019.
Els darrers temps, en la restauració d’obres d’infraestructures viàries, ha estat freqüent –en general a Catalunya, no només en l’àmbit considerat- la sembra d’un Lotus corniculatus especialment robust i erecto-ascendent. El tractament taxonòmic més acceptat per a aquestes plantes és el de varietat, però existeixen propostes d’elevar-les a rang específic com a L. sativus (Hyl.) Buscher & G. H. Loos (Loos, 2010), que han tingut una acceptació limitada en cercles botànics del centre i nord d’Europa (ex. Buttler & Hand, 2011; NBIC, 2019). Amb aquesta nota ens limitem a constatar la presència al territori considerat d’aquestes plantes aŀlòctones, sense valorar quin és el seu tractament taxonòmic més adequat. Indiquem com a exemple dues localitats del Berguedà en les quals hem observat que ha establert poblacions persistents durant un mínim d’una desena d’anys, però també hem vist aquesta planta en força altres punts en els quals no hem seguit la seva dinàmica.
Lotus tenuis Mill.
Berguedà: Berga, autovia C-16 (eix del Llobregat), DG0562, 695 m, marge de carretera, 1-IX-2018.
La presència d’aquesta espècie en l’àmbit geogràfic considerat no havia estat confirmada. Les úniques citacions corresponen a l’entorn de Bagà (Lapraz, 1958) i han estat considerades errònies (Soriano, 1992; Vigo et al., 2003; BDBC). La població trobada a Berga, de desenes d’individus, sembla atribuïble a una colonització recent i s’hi comporta com a planta viària. Les localitats conegudes més pròximes es troben a la part central del Bages.
Malva arborea (L.) Webb & Berthel. [Lavatera arborea L.]
B
ageS: Cardona, perifèria sud de la zona urbana, al costat d ela carretera al Miracle, CG9040, 540 m, vegetació ruderal, 7-II-2015; Súria, vora Can Sivila, CG9532, 300 m, vegetació ruderal, 7-II-2015.
Aparentment, aquesta espècie no havia estat citada de la Catalunya interior, més cap a l’oest de les serres prelitorals (BDBC). La flora en línia d’Aragó (http://floragon.ipe.csic.es/) sí que n’indica alguna localitat lluny del litoral mediterrani, una de les quals és a la Franja (Purroi, Baixa Ribagorça). Els dos petits nuclis del Bages provenien de l’escapament de jardins pròxims, en què es cultivava com a planta ornamental.
Mesembryanthemum ×vascosilvae (Gideon F. Sm., E. Laguna, F. Verloove & P.P. Ferrer) L. Sáez & P. Aymerich [Aptenia ×vascosilvae Gideon F. Sm., E. Laguna, F. Verloove & P.P. Ferrer]
Berguedà: Berga, nucli urbà en dos punts diferents, DG0461, 660 i 680 m, fissures de paviment al peu d’una paret, 8-X2020 i 15-II-2021..
Cas similar al de Kalanchoe ×houghtonii indicat abans, d’una espècie crassa hibridògena i artificial que es naturalitza amb rapidesa a la regió mediterrània. Documentem també la seva presència casual (dos individus isolats i de persistència incerta) en hàbitats urbans de l’interior de Catalunya, on només en coneixem una observació prèvia (publicada com a Aptenia cordifolia) en una urbanització del Bages (Aymerich, 2015a).
Moricandia arvensis (L.) DC.
Berguedà: Berga, solell de la serra de Queralt, al voltant de la casa de Fumanya, DG0362, 850-860 m, 2018-2020.
Espècie mediterrània que els darrers temps s’ha estès molt a Catalunya fora de la seva àrea tradicional de presència, com a planta viària, ja sigui per transport de terres amb llavors o per sembra intencionada. Documentem la seva presència puntual al Berguedà, en una localitat en què va aparèixer l’any 2018 associada a les obres d’arranjament d’un camí, i on ha mantingut una població relativament nombrosa fins a l’actualitat. En l’àmbit d’aquest treball, només en coneixem una citació prèvia a l’alt Cardener (BDBC).
Oxalis latifolia Kunth
Berguedà: Berga, nucli urbà, DG0461, 680 m, vegetació ruderal en un jardí abandonat, 8-I-2018.
Espècie que, en sentit estricte (plantes de fulles molt anguloses i flors roses), no havia estat citada en l’àmbit d’aquest article. Sí que s’hi coneixia O. vallicola (Rose) Kunth, planta de flors blanques i fulles arrodonides que sovint s’ha considerat com a una forma d’O. latifolia.
Panicum capillare L. subsp. capillare
Berguedà: Olvan, polígon industrial, DG0856, 525 m, marges de camins, 8-X-2020; Vilada, embassament de la Baells, DG0764, 640 m, marge de carretera, 17-IX-2019.
Tàxon aŀlòcton que no havia estat indicat al Berguedà, tot i ser-hi previsible. En l’àmbit considerat, només havia estat citat a la vall del Segre a l’Alt Urgell, mentre que a la del Llobregat les dades més pròximes es situen a l’alçada de Manresa (BDBC).
Sagina micropetala Rauschert [S. apetala subsp. erecta (Hornem.) Herm.).]
Berguedà: Berga, nucli urbà, DG0461-0561, 670-690 m, fissures de paviment, 2017-2020; Vilada, riba est del pont de la Baells, DG0764, 640 m, terreny sorrenc en una zona d’aparcament, 3-V-2018; SolSonèS: Guixers, vora la cruïlla de les carreteres a Berga i a Solsona, CG8465, 825 m, fissures de paviment, 24-VI-2019.
Tàxon del qual, a la zona considerada, només coneixem una citació a Alp, Cerdanya (BDBC). Les tres localitats que aportem corresponen a hàbitats antròpics, cosa que permet
50 Butlletí ICHN 85 (2), 2021
P. AYMERICH BOIXADER
suposar que aquesta planta hi ha arribat gràcies a una dispersió recent per mitjans humans.
Sedum caespitosum (Cav.) DC.
Berguedà: la Nou de Berguedà, carretera Barcelona-Andorra al nord del Far, DG0672, 680 m, marge de carretera, 16-V2017.
Teròfit no gaire rar a la Catalunya mediterrània, però absent dels Pirineus, amb l’excepció del seu extrem oriental marítim. Considerem que la presència en aquesta localitat és accidental i que segurament deriva del transport de graves o terres per a treballs de manteniment de la carretera. L’any 2017 presentava una població relativament nombrosa, d’unes quantes desenes d’individus. Es mantenia l’any 2018, però la quantitat d’individus semblava que havia disminuït. Les localitats més pròximes conegudes se situen al Moianès (Mercadé, 2016).
Silene noctiflora L.
Berguedà: Avià, Graugés, DG0457, 595 m, marge de camí, 20-VII-2019.
Espècie detectada a Catalunya fa pocs anys a la Cerdanya (Aymerich, 2013b), on està ben establerta, i que més endavant es va trobar també a l’Alt Urgell (Aymerich, 2016b). La nova observació del Berguedà, d’una quinzena d’individus, és la primera que es fa fora de la conca superior del riu Segre. Ha de correspondre a una colonització molt recent, perquè hem visitat aquest lloc de forma anual en els darrers temps i fins a 2019 no hi havíem vist mai S. noctiflora.
Sinapis alba L. subsp. mairei (H. Lindb.) Maire
Berguedà: Berga, al nord de la Colònia Rosal, DG0658, 505 m, talús amb vegetació ruderal, 18-IV-2018.
Tàxon que a Catalunya es concentra a la franja marítima (BDBC) i que no havia estat citat fins ara de la conca mitjana i alta del riu Llobregat. La població observada estava formada per desenes d’individus i és probable que sigui el resultat d’una colonització recent.
Vicia villosa Roth.
Berguedà: Olvan, al sud de la Colònia Rosal, DG0657, 475 m, camp semiabandonat, 21-X-2018.
Espècie aŀlòctona que ha estat molt poc citada a Catalunya (BDBC) i de la qual no coneixem indicacions al Berguedà. La seva presència en aquesta localitat –on era abundant- s’ha d’atribuir, com és habitual, a la persistència després de sembres realitzades per a fer-la servir com a farratge.
Vinca difformis Pourr.
Berguedà: Berga, al nord-est de l’hospital comarcal, DG0562, 740 m, herbassar en un talús entre antics camps abandonats, 1-II-2020; Santa Maria de Merlès, al sud del molí de Vilalta, DG1652, 545 m, herbassar al talús d’un camp, 23-VII-2020.
En l’àmbit territorial considerat, gairebé no s’ha citat aquesta espècie i les informacions existents són poc clares.
El BDBC en recull només dues dades, una de les quals, al quadrat UTM CG95 (Berguedà), deriva de l’Atles ORCA (Bolòs et al., 2001) i no se’n coneixen ni la localitat ni la font original. L’altra sí que està ben documentada i correspon a la Coma (Solsonès), però originalment va ser citada de forma insegura, com a Vinca cf. difformis (Vives, 1964). Les dues observacions noves corresponen a taques clonals de 5-10 m2 , i tindrien el seu origen en una resta de cultiu. A la zona considerada, la Vinca més cultivada com a ornamental és V. major, seguida de V. minor, molt més rara; en canvi, fins ara mai no hi havíem observat V. difformis.
Referències
Aymerich, P. 1998. Aportació al coneixement florístic del nord de Catalunya. Butlletí de la Institució Catalana d’Història Natural, 66: 41-57.
Aymerich, P. 2003. Notes florístiques de les conques altes dels rius Segre i Llobregat. Acta Botanica Barcinonensia, 48: 15-28.
Aymerich, P. 2008. Notes florístiques de l’alt Pallars. Acta Botanica Barcinonensia, 51: 17-33.
Aymerich, P. 2012a. Juncus pyrenaeus, P. 13. In: Fernández, J. (coord.) Anuari Naturalista del Parc Natural de l’Alt Pirineu 2012. 58 p.
Aymerich, P. 2012b. Potamogeton i Zannichellia a la conca mitjana del riu Llobregat (Catalunya, NE de la península Ibèrica). Orsis, 26: 57-89.
Aymerich, P. 2013a. Contribució al coneixement florístic del territori ausosegàrric (NE de la península Ibèrica). Orsis, 27: 209-259.
Aymerich, P. 2013b. Notes sobre algunes plantes, rares o amenaçades als Pirineus catalans. Butlletí de la Institució Catalana d’Història Natural, 77: 5-26.
Aymerich, P. 2013c. Sobre algunes espècies aŀlòctones a l’alt Segre (NE de la península Ibèrica), noves o molt rares per a la flora catalana. Orsis, 27: 195-207.
Aymerich, P. 2014. Notes florístiques de les conques altes dels rius Segre i Llobregat (II). Orsis, 28: 7-47.
Aymerich, P. 2015a. Nuevos datos sobre plantas suculentas alóctonas en Cataluña. Bouteloua, 22: 99-116.
Aymerich, P. 2015b. Notes florístiques de les conques altes dels rius Segre i Llobregat (III). Orsis, 29: 1-28.
Aymerich, P. 2016a. Contribució al coneixement de la flora aŀlòctona del nord i centre de Catalunya. Orsis, 30: 11-40.
Aymerich, P. 2016b. Notes florístiques de les conques altes dels rius
Segre i Llobregat (IV). Orsis, 30: 133-165.
Aymerich, P. 2017. Notes sobre flora aŀlòctona a Catalunya. Butlletí de la Institució Catalana d’Història Natural, 81: 97-116.
Aymerich, P. 2019. Notes sobre flora aŀlòctona a Catalunya. II. Butlletí de la Institució Catalana d’Història Natural, 83: 3-21.
Aymerich, P. 2020. Rhinanthus alectorolophus (Orobanchaceae) a la península Ibèrica. Butlletí de la Institució Catalana d’Història Natural, 84: 125-126.
Aymerich, P., Soriano, I. & Llistosella, J. 2008. Addicions a la flora vascular del Parc Natural del Cadí-Moixeró i de les serres veïnes (Prepirineus orientals ibèrics). Acta Botanica Barcinonensia, 51: 35-47.
Aymerich, P., Guadiola, M., Petit, A., Ballesteros, E. & Chappuis, E. 2014. Distribució, ecologia i estat de conservació de Potamogeton lucens i P. schweinfurthii a Catalunya. Butlletí de la Institució Catalana d’Història Natural, 78: 25-37.
51 Butlletí ICHN 85 (2), 2021 GEA, FLORA ET FAUNA
NOTES FLORÍSTIQUES DE LES CONQUES ALTES DELS RIUS SEGRE I LLOBREGAT. V
GEA, FLORA ET FAUNA
Aymerich, P. & Sáez, L. 2015a. Alchemilla cadinensis (Rosaceae), a new species from the Pyrenees (SW Europe). Willdenowia, 45: 435-442.
Aymerich, P. & Sáez, L. 2015b. Comentaris i precisions previs a la Checklist de la flora de Catalunya (nord-est de la península Ibèrica). Orsis, 29: 1-68.
Aymerich, P. & Sáez, L. 2019. Checklist of the vascular alien flora of Catalonia (northeastern Iberian Peninsula, Spain). Mediterranean Botany, 40 (2): 215-242.
Aymerich, P. & Soriano, I. 2016. Presència d’Achillea roseoalba (Asteraceae) Pirineus. Butlletí de la Institució Catalana d’Història Natural, 80: 93-112.
Bolòs, O., Font, X., Pons, X. & Vigo, J. (eds.) 1998. ORCA. Atlas corològic de la flora vascular dels Països Catalans, 8. Institut d’Estudis Catalans. Barcelona.
Bolòs, O., Font, X., & Vigo, J. (eds.) 1999. ORCA. Atlas corològic de la flora vascular dels Països Catalans, 9. Institut d’Estudis Catalans. Barcelona.
Bolòs, O., Font, X., & Vigo, J. (eds.) 2001. ORCA. Atlas corològic de la flora vascular dels Països Catalans, 11. Institut d’Estudis Catalans. Barcelona.
Bolòs, O. & Vigo, J. 1984. Flora dels Països Catalans Vol. 1. Ed. Barcino. Barcelona. 736 p.
Bolòs, O. & Vigo, J. 1990. Flora dels Països Catalans Vol. 2. Ed. Barcino. Barcelona. 921 p.
Bolòs, O. & Vigo, J. 1996. Flora dels Països Catalans. Vol. 3. Ed. Barcino. Barcelona. 1230 p.
Bolòs, O. & Vigo, J. 2001. Flora dels Països Catalans Vol. 4. Ed. Barcino. Barcelona. 750 p.
Bolòs, O., Vigo, J., Masalles, R. M. & Ninot, J. M. 2005. Flora manual dels Països Catalans. Ed. Pòrtic. Barcelona. 1310 p.
Busquets, J. M., Gorchs, M. Macarro, J., Oriola, J. & Viladrich, L. 1983. Recull d’observacions biològiques i geològiques efectuades a les muntanyes del Berguedà. L’Erol, 7: 13-17.
Buttler, K. P. & Hand, R. 2011. Beiträge zur Fortschreibung der Florenliste Deutschlands (Pteridophyta, Spermatophyta) – Vierte Fole. Kochia, 5: 83-91.
Carreras, J. 1993. Flora i vegetació de Sant Joan de l’Erm i de la vall de Santa Magdalena. Coŀlecció Estudis, 3. Institut d’Estudis Ilerdencs. Lleida. 321 p.
Carrillo, E., Mercadé, A., Ninot, J. M., Carreras, J., Ferré, A. & Font, X. 2008. Check-list i Llista vermella de la flora d’Andorra. CENMA-IEA-Ministeri d’Ordenament Territorial, Urbanisme i Medi Ambient. Andorra. 488 p.
Casas, C. & Ninot, J. M. 1996. Estudi fitocenològic de les pastures de la Plana de Vic. II: comunitats terofítiques (TheroBrachypodietea) i síntesi. Butlletí de la Institució Catalana d’Història Natural, 63: 27-50.
Casasayas, T. 1989. La flora aŀlòctona de Catalunya. Tesi doctoral. Facultat de Biologia, Universitat de Barcelona. 880 p.
Devis, J., Ferré, A. & Soriano, I. 2001. Datos sobre vegetación de las sierras de Turp y Odèn. Matorrales xeroacánticos y sabinares. P. 395-407. In: Gómez, F.; Mota, F.J. (eds.) Vegetación y cambios climáticos. Universidad de Almería. 500 p.
Font, J. 2000. Estudis botànics de la serra de l’Albera. Catàleg florístic general i poblament vegetal de les Basses de l’Albera. Tesi doctoral. Universitat de Girona. 692 p.
Font, X., & Vigo, J. (eds.) 2008. ORCA. Atlas corològic de la flora vascular dels Països Catalans, 15. Institut d’Estudis Catalans. Barcelona.
González, V. & Nuet, J. 2008. Flora i vegetació de la bassa de Sanavastre. Cerdanya. Lulu Enterprises. Morrisville. 138 p.
Gruber, M. 1977. Contribution à la flore des Pyrénées ariègeoises et catalanes: 2e note. Bulletin Societé Botanique de France, 124: 93-104.
Guardiola M., Petit, A., Molero, J. & Sáez, L. 2016. Aportacions al coneixement de la flora vascular del massís de Boumort i serres veïnes. Orsis, 30: 67-100.
Lapraz, G. 1958. Notes sur les landes et pélouses des environs des environs de Bagà (Pyrénées catalanes). Collectanea Botanica, 5: 867-872.
Lewin, J. M. 2018. Compléments à la connaissance de la flore des Pyrénées-Orientales (suite 2). Mycologie et Botanique (Bulletin de la Societé mycologique et botanique de Catalogne Nord), 33: 46-52.
Lewin, J. M., Bouteloup, R, Schwab, P. & Nicolas, S. 2020. Nouvelles contributions à la flore des Pyrénées-Orientales. Observations 2019-2020. Mycologie et Botanique (Bulletin de la Societé mycologique et botanique de Catalogne Nord), 35: 3952.
Loos, G. H. 2010. Taxonomische Neukombinationen zur Flora Mittel – und Osteuropas, insbesondere Nordrhein-Westfalens. OnlineVeröffentlichungen Bochumer Botanische Verein, 2: 1-20.
Mercadal, G. 2019. Els prats de dall de la terra baixa catalana. Caracterització geobotànica, valoració agroambiental i estudi de les relacions fitosociològiques entre els prats dalladors de l’Europa occidental. Universitat de Girona. 2306 p. https://www. tdx.cat/handle/10803/669044#page=1
Mercadé, A. 2016. Estudis de vegetació i flora del Moianès i àrees properes. Tesi doctoral. Universitat de Barcelona. 619 p.
Mesquida, V., Gómez-Bellver, C., Guillot, D., Herrando, S., Nualart, N., Sáez, L. & López-Pujol, J. 2017. El gènere Kalanchoe (Crassulaceae) a Catalunya: situació i distribució potencial del tàxon invasor Kalanchoe x houghtonii Orsis, 31: 37-64.
Molero, J., Sáez, L., Vallverdú, J. & Valle, L. G. 2006. Noves aportacions al coneixement de la flora vascular de les comarques meridionals de Catalunya. III. Orsis, 21: 19-39.
Molina, J.. Michaud, H., Tison, J. M., Fernández, R. & Véla, E. 2018. Allium scaberrimum. The IUCN Red List of Threatened Species: e.T110805790A87775132.
Montserrat, P. 1957. Contribución al estudio de los prados próximos a Seo de Urgel. Publicaciones del Instituto de Biología Aplicada, 25: 49-112.
NBIC-Norwegian Biodiversity Information Centre. 2019. Lotus sativus. Disponible en: http://beta.biodiversity.no/ScientificName/ Lotus/101967 [Data de consulta: agost de 2020]
Pyke, S. 2010. Three recently-introduced alien grasses in the Iberian Peninsula. Collectanea Botanica, 29: 91-93.
Rifà, P. 2015. Impatiens glandulifera naturalitzada al riu Ter. Butlletí de la Institució Catalana d’Història Natural, 79: 111-112.
Sáez, L., Devis, J. & Soriano, I. 2004. Flora vascular de la vall d’Alinyà. P. 237-300. In: Germain, J. (ed.) Els sistemes naturals de la vall d’Alinyà. Treballs de la Institució Catalana d’Història Natural, 14. 608 p.
Sáez, L., Molero, J., Carrillo, E., Ninot, J. M., Guardiola, M., Guàrdia Valle, L., Macías, C & Aymerich, P. 2008. Noves contribucions al coneixement de la flora vascular del massís del Boumort (Prepirineus ibèrics, NE de la península Ibèrica). Orsis, 23 : 137162.
Sáez, L., Aymerich, P. & Blanché, C. 2010. Llibre Vermell de les plantes vasculars endèmiques i amenaçades de Catalunya. Ed. Argania. Barcelona. 811 p.
Soriano, I. 1984. Les valls del Bastareny (Alt Berguedà): Flora i paisatge vegetal. Tesi de llicenciatura. Universitat de Barcelona. 359 p.
52 Butlletí ICHN 85 (2), 2021
P. AYMERICH BOIXADER
Soriano, I. 1992. Estudi florístic i geobotànic de la serra de Moixeró i el massís de la Tosa d’Alp (Pirineus orientals). Tesi doctoral microfitxada. Universitat de Barcelona.
Soriano, I. 2019. Achillea L. P. 1753-1774. In: Benedí, C.; Buira, A.; Rico, E.; Crespo, M.B.; Quintanar, A; Aedo, C. (eds.) Flora iberica. Vol. XVI(III). Compositae (partim). Real Jardín BotánicoCSIC. Madrid. 2353 p.
Soriano, I. & Aymerich, P. 2017. Precisions sobre la distribució i l’estatus de Carex brevicollis (Cyperaceae) a Catalunya. Butlletí de la Institució Catalana d’Història Natural, 81: 117-119.
Valdés, B. 2000. Onobrychis Mill. P. 955-970. In: Talavera, S., Aedo, C., Castroviejo, S., Herrero, A., Romero, C., Salgueiro, F.
J. & Velayos, M. (eds.) Flora iberica. Vol. VII(II). Leguminosae (partim). Real Jardín Botánico-CSIC. Madrid. 1119 p.
Verllove, F. & Aymerich, P. 2020. Chorological novelties for the alien flora of northeastern Catalonia (Iberian Peninsula). Butlletí de la Institució Catalana d’Història Natural, 84: 137-153.
Vigo, J., Soriano, I., Carreras, J., Aymerich, P., Carrillo, E., Font, X., Masalles, R. M. & Ninot, J. M. 2003. Flora del Parc Natural del Cadí-Moixeró i de les serres veïnes. Monografies del Museu de Ciències Naturals-Institut Botànic de Barcelona. 407 p.
Vives, J. 1964. Vegetación de la alta cuenca del Cardener. Estudio florístico y fitocenológico comarcal. Acta Geobotanica Barcinonensia. Vol. I. 218 p.
53 Butlletí ICHN 85 (2), 2021
FLORA ET FAUNA
GEA,
ALTES DELS RIUS SEGRE I LLOBREGAT. V
NOTES FLORÍSTIQUES DE LES CONQUES
Butlletí de la Institució Catalana d’Història Natural, 85 (2): 55-58. 2021
NOTA BREU
NOTA BREU
La xinxa de l’eucaliptus, Thaumastocoris peregrinus Carpintero & Dellapé, nou heteròpter exòtic per a Catalunya (Hemiptera: Thaumastocoridae)
The bronze bug, Thaumastocoris peregrinus Carpintero & Dellapé, new exotic true bug for Catalonia (Hemiptera: Thaumastocoridae)
Jan Tomàs*, Adrià Miralles-Núñez** & Adrián Necoechea***
* C/ Escultor Armengol, 16, 2n 2a, 08222. Terrassa (Barcelona). A/e: jan.tomas.6@hotmail.com
** Servicios Depec. Responsable d’Entomologia. C/ Nena Casas, 71, 08017. Barcelona.
*** Servicios Depec. Direcció tècnica. C/ Nena Casas, 71, 08017. Barcelona.
Rebut: 12.04.2021. Acceptat: 09.06.2021. Publicat: 30.06.2021
El gènere Thaumastocoris Kirkaldy, 1908 (Thaumastocoridae), endèmic d’Austràlia, comprèn 14 espècies (Noack et al., 2011). Una d’aquestes, Thaumastocoris peregrinus Carpintero & Dellapé, 2006 (xinxa de l’eucaliptus), però, va ser descrita a partir d’exemplars coŀlectats el novembre de 2005 a La Plata (Buenos Aires, Argentina), on, segons sembla, hauria estat introduïda de manera accidental a través de la importació d’eucaliptus (Eucalyptus spp.) de la regió australiana. Tanmateix, aquesta espècie ja s’havia trobat el 2003 a la província sud-africana de Gauteng (Jacobs & Neser, 2005) i, posteriorment, l’abril de 2005 al municipi de Moreno (Buenos Aires, Argentina) (Noack & Coviella, 2006), si bé fins la publicació de Carpintero & Dellapé (2006) no es va reconèixer com a nova espècie sinó com a T. australicus Kirkaldy, 1908.
A Austràlia (Sydney), la xinxa de l’eucaliptus va començar a ocasionar estralls el 2001 (Noack et al., 2011). D’ençà d’aleshores, s’ha estès àmpliament per gairebé tot l’hemisferi sud, gran part de l’Europa meridional i alguns països de l’Amèrica del Nord i de l’Orient Mitjà (EPPO, 2021).
Des que el 2012 se’n varen capturar els primers exemplars a la península Ibèrica (Lisboa, Portugal) (Garcia et al., 2013), T. peregrinus ja està ben establert a Portugal, especialment a la meitat sud (ICNF, 2017), i s’ha detectat a Extremadura (Vivas et al., 2015), la Comunitat de Madrid (Machado et al., 2019) i Andalusia (Pérez-Gómez et al., 2020). Machado et al. (2019, 2020) apunten que el més probable és que s’introduís a Portugal a través de la importació de llenya d’eucaliptus de l’Amèrica del Sud, que és el principal proveïdor d’aquesta fusta a Europa (González-Gómez, 2019), i que posteriorment s’hagués expandit a Espanya de manera natural o bé com a conseqüència de l’activitat humana; no obstant, també plantegen la possibilitat que arribés a territori espanyol directament des de l’Amèrica Llatina. Tant la informació genètica obtinguda per aquests mateixos autors com la troballa a Portugal de dues larves del neuròpter neotropical Hemerobius
bolivari Banks, 1910 depredant sobre nimfes d’aquesta xinxa (Garcia et al., 2013) reforcen la hipòtesi sobre l’origen sudamericà de les poblacions ibèriques de T. peregrinus.
Actualment, T. peregrinus és una plaga emergent a nivell global que afecta una cinquantena (entre espècies i híbrids) de plantes llenyoses de la família Myrtaceae, concretament dels gèneres Eucalyptus L’Hér. (38 espècies i 8 híbrids) i Corymbia K.D. Hill & L.A.S. Johnson (4 espècies) (Saavedra et al., 2015b).
Segons el model de distribució potencial de Saavedra et al. (2015a), el litoral de Catalunya reuneix les condicions idònies per a l’establiment de T. peregrinus, si bé els esforços per trobar-l’hi de Machado et al. (2019) i dels mateixos autors d’aquesta nota, fins ara, havien estat infructuosos. Així doncs, en el present treball s’aporten les primeres citacions per a Catalunya de la xinxa de l’eucaliptus.
El 14 d’octubre de 2020, al municipi de Vilanova del Vallès (Vallès Oriental, Barcelona) (31TDG40) es van capturar una gran quantitat d’exemplars d’aquest heteròpter, tant adults com nimfes de tots els estadis (Fig. 1), sobre dos Eucalyptus, probablement E. globulus Labill., plantats en un jardí particular des de fa més de 30 anys. Els propietaris de la finca s’havien adonat de la presència de l’insecte feia cosa d’un mes, quan els arbres havien començat a perdre moltes fulles i l’elevada densitat d’individus (de l’ordre de centenars) ja els ocasionava molèsties. Aquell mateix dia es va detectar l’espècie en més peus d’E. globulus del municipi, cosa que sembla indicar que hi està força estesa. Gairebé un mes després, el 10 de novembre, es van observar diversos exemplars de T. peregrinus al port de Barcelona (31TDF27), sobre un eucaliptus (no identificat) plantat a la via pública (C. Pradera, com. pers.).
Tant les nimfes com els adults de T. peregrinus s’alimenten del mesofiŀle de les fulles madures de les seves plantes hostes, en concret del clorènquima (Santadino et al., 2017). El principal símptoma de la infestació és l’aparició de taques
55 Butlletí ICHN 85 (2), 2021
ISSN:
(print edition)
ISSN 2013-3987 (online edition):
1133-6889
10.2436/20.1502.01.73
DOI:

cloròtiques/necròtiques en les fulles (Fig. 2), que primer adquireixen tonalitats argentades, després bronzejades i finalment vermelloses o brunenques (Laudonia & Sasso, 2012), i que acaben caient de l’arbre prematurament (defoliació). Quan aquest està greument infestat, la caiguda massiva del fullatge pot donar lloc a la mort de branques (Nadel et al., 2010) i, en alguns casos, fins i tot del mateix arbre (Jacobs & Neser, 2005).
La xinxa de l’eucaliptus presenta un cicle vital curt, d’entre un i dos mesos en funció de la temperatura (vegeu, per exemple, Barbosa et al., 2019), de manera que pot tenir diverses generacions a l’any (Jacobs & Neser, 2005). Al llarg de la seva vida, cada femella és capaç de pondre uns 60 ous o més i tot, depenent de la temperatura i de la planta hoste (vegeu, per exemple, Soliman et al., 2012). Les nimfes, que neixen al cap d’uns sis dies, assoleixen l’estat adult en poc més de dues setmanes, període durant el qual passen per cinc estadis (Noack & Rose, 2007). La curta durada de les fases preimaginals permet que en una mateixa fulla puguin trobarse alhora postes, nimfes i adults (Wilcken et al., 2010).
Amb aquestes troballes, doncs, es constata l’arribada a Catalunya d’un nou heteròpter exòtic, que afecta un gran nombre d’espècies d’Eucalyptus. En el conjunt de la Península, les plantacions d’eucaliptus, especialment d’E. globulus, són molt importants per a les indústries paperera i de la fusta (FAO, 1981). A Espanya, Galícia és el principal productor de fusta d’eucaliptus, i és per això que l’establiment de T. peregrinus, que de moment no s’hi ha detectat (Machado et al., 2019), pot arribar a causar-hi estralls. Els eucaliptus, a més, són arbres
d’alt valor ornamental, i aquells peus molt longeus o de grans dimensions també d’un valor patrimonial indubtable, de manera que no s’ha de deixar de banda l’impacte estètic i de salut que hi pugui tenir la infestació per aquesta xinxa. Malgrat ser un insecte alat, T. peregrinus no és bon volador (Cuello et al., 2019); en canvi, sí que té una gran capacitat de dispersió, ja sigui a través dels desplaçaments de persones i mercaderies o, també, per mitjà del vent, d’ocells o de material vegetal infestat (Nadel et al., 2010; Machado et al., 2020).
En l’actualitat, no es disposa de cap estratègia de control efectiva per lluitar contra aquest insecte, encara que s’ha demostrat que alguns productes fitosanitaris poden reduir-ne significativament les poblacions (Machado et al., 2016). Ara bé, atès l’impacte ambiental d’aquests insecticides químics, el control biològic sembla l’única alternativa viable per fer front a aquesta plaga en plantacions comercials d’eucaliptus (Wilcken et al., 2019). En aquest sentit, a Austràlia s’han trobat dos himenòpters de la família Mymaridae que actuen com a parasitoides dels ous de T. peregrinus: Cleruchoides noackae Lin & Huber, 2007 i una espècie no identificada del gènere Stethynium Enock, 1909 (Lin et al., 2007).
Si bé no hi ha evidències clares sobre la incidència de T. peregrinus en la salut pública, s’han notificat casos d’irritació cutània per les seves picades en persones que havien estat a prop d’eucaliptus infestats, principalment en parcs urbans (Jacobs & Neser, 2005). Malgrat tot, no sembla tractar-se d’un insecte d’interès mèdic. Així doncs, cal estar atents a l’evolució d’aquesta espècie a Catalunya, en espera de veure quins efectes pot tenir-hi la seva arribada.
56 Butlletí ICHN 85 (2), 2021 NOTA BREU J. TOMÀS ET AL.
Figura 1. Adult (a) i nimfes de 3r, 4t i 5è estadi de Thaumastocoris peregrinus al revers d’una fulla d’eucaliptus. Escala = 1 mm. Foto: Adrià Miralles-Núñez.

D’acord amb Goula et al. (2020), la fauna catalana d’heteròpters està formada per 1.073 espècies, pertanyents a 416 gèneres. Amb T. peregrinus, Cimex hemipterus (Fabricius, 1803) (Cimicidae) (Pradera & Ruiz, 2020) i Stephanitis (S.) lauri Rietschel, 2014 (Tingidae) (Riba-Flinch & Goula, 2021), el nombre de gèneres s’eleva a 417 i el d’espècies a 1.076. De resultes de la troballa a Catalunya de T. peregrinus, la família Thaumastocoridae també s’hi cita per primera vegada.
Agraïments
Volem expressar la nostra més sincera gratitud a Yolanda Lorenzo, alcaldessa de Vilanova del Vallès, a Jordi Vallllovera, regidor del mateix consistori, i als propietaris de la finca particular per haver-nos atès tan amablement durant la visita al seu municipi. A Carlos Pradera per comunicar-nos la troballa de T. peregrinus al port de Barcelona. Finalment, i en especial, a Marta Goula i a Marcos Roca-Cusachs, de la Universitat de Barcelona, per la revisió crítica del manuscrit.
Bibliografia
Barbosa, L. R., Santos, F., Soliman, E. P., Rodrigues, A. P., Wilcken, C. F., Campos, J. M., Zanuncio, A. J. V. & Zanuncio, J. C. 2019.
Biological parameters, life table and thermal requirements of Thaumastocoris peregrinus (Heteroptera: Thaumastocoridae) at different temperatures. Scientific Reports, 9: 10174.
Carpintero, D. L. & Dellapé, P. M. 2006. A new species of Thaumastocoris Kirkaldy from Argentina (Heteroptera: Thaumastocoridae: Thaumastocorinae). Zootaxa, 1228: 61-68.
Cuello, E. M., Andorno, A. V., Hernández, C. M. & López, S. N. 2019. Population Development of the Invasive Species Thaumastocoris peregrinus (Hemiptera: Thaumastocoridae) on four Eucalyptus Species of the Subgenus Symphyomyrtus Neotropical Entomology, 48 (1): 152-161.
EPPO. 2021. EPPO Global Database. Thaumastocoris peregrinus Disponible en: https://gd.eppo.int/taxon/THMCPE/distribution [Data de consulta: 21 febrer 2021].
FAO. 1981. Eucalypts for Planting. 2a ed. FAO. Roma. 677 p.
Garcia, A., Figueiredo, E., Valente, C., Monserrat, V. J. & Branco, M. 2013. First record of Thaumastocoris peregrinus in Portugal and of the neotropical predator Hemerobius bolivari in Europe. Bulletin of Insectology, 66 (2): 251-256.
González-Gómez, M. 2019. Dynamics of pulp exports from South America to the European Union. Austrian Journal of Forest Science, 136 (4): 313-330.
Goula, M., Osorio, V., Tomàs, J. & Roca-Cusachs, M. 2020. Llista dels Heteròpters de Catalunya (Insecta, Hemiptera, Heteroptera) Versió 2, juliol 2020. Publicacions del Centre de Recursos de Biodiversitat Animal. Universitat de Barcelona, vol. 6, 46 p. ICNF. 2017. Programa Operacional de Sanidade Florestal. Relatório de execução – 2016. Disponible en: http://www.icnf.pt/portal/ florestas/prag-doe/resource/doc/posf/POSF-Relatorio-execucao2016-20JUL2017 [Data de consulta: 21 febrer 2021].
Jacobs, D. H. & Neser, S. 2005. Thaumastocoris australicus Kirkaldy (Heteroptera: Thaumastocoridae): a new insect arrival in South Africa, damaging to Eucalyptus trees. South African Journal of Science, 101 (5): 233-236.
Laudonia, S. & Sasso, R. 2012. The bronze bug Thaumastocoris peregrinus: a new insect recorded in Italy, damaging to Eucalyptus trees. Bulletin of Insectology, 65 (1): 89-93.
Lin, N. Q., Huber, J. T. & La Salle, J. 2007. The Australian Genera of Mymaridae (Hymenoptera: Chalcidoidea). Zootaxa, 1596: 1-111.
Machado, D. N., Costa, E. C., Garlet, J., Boscardin, J., Pedron, L., Perini, C. R. & Bolzan, L. 2016. Avaliação de Inseticidas no Controle de Thaumastocoris peregrinus (Hemiptera: Thaumastocoridae) Percevejo-bronzeado em Condições de Laboratório. Floresta e Ambiente, 23 (2): 245-250.
Machado, D. N., Costa, E. C., Guedes, J., Barbosa, L. R., Martínez, G., Ide, S., Ramos, S. O., Branco, M., Garcia, A., Vanegas-Rico, J. M., Jiménez-Quiroz, E., Laudonia, S., Novoselsky, T., Hodel, D. R., Arakelian, G., Silva, H., Perini, C. R., Valmorbida, I., Ugalde, G. A. & Arnemann, J. A. 2020. One maternal lineage leads the expansion of Thaumastocoris peregrinus (Hemiptera: Thaumastocoridae) in the New and Old Worlds. Scientific Reports, 10: 3487.
Machado, D. N., Costa, E. C., Perini, C. R., Ugalde, G. A., Saldanha, M. A., Leitão, J. V., Colpo, T. L., Arnemman, J. A. & CorderoRivera, A. 2019. The ongoing dispersion of the Eucalyptus bronze bug (Thaumastocoris peregrinus) in Spain. Forest Systems, 28 (3): eSC03, 7 p.
Nadel, R. L., Slippers, B., Scholes, M. C., Lawson, S. A., Noack, A. E., Wilcken, C. F., Bouvet, J. P. & Wingfield, M. J. 2010. DNA bar-
57 Butlletí ICHN 85 (2), 2021 NOTA BREU LA XINXA DE L'EUCALIPTUS, THAUMASTOCORIS PEREGRINUS, NOU HETERÒPTER EXÒTIC PER A CATALUNYA
Figura 2. Danys provocats per Thaumastocoris peregrinus en fulles d’Eucalyptus sp. Foto: Adrià Miralles-Núñez.
NOTA BREU
coding reveals source and patterns of Thaumastocoris peregrinus invasions in South Africa and South America. Biological Invasions, 12 (5): 1067-1077.
Noack, A. E., Cassis, G. & Rose, H. A. 2011. Systematic revision of Thaumastocoris Kirkaldy (Hemiptera: Heteroptera: Thaumastocoridae). Zootaxa, 3121: 1-60.
Noack, A. E. & Coviella, C. E. 2006. Thaumastocoris australicus Kirkaldy (Hemiptera: Thaumastocoridae): first record of this invasive pest of Eucalyptus in the Americas. General & Applied Entomology, 35: 13-14.
Noack, A. E. & Rose, H. A. 2007. Life-history of Thaumastocoris peregrinus and Thaumastocoris sp. in the laboratory with some observations on behaviour. General & Applied Entomology, 36: 27-33.
Pérez-Gómez, A., Sánchez, I. & Baena, M. 2020. Nuevos registros de hemípteros (Insecta: Hemiptera) alóctonos en Andalucía (sur de España). Revista de la Sociedad Gaditana de Historia Natural, 14: 9-19.
Pradera, C. & Ruiz, J. 2020. Primera detección de chinche de cama tropical, Cimex hemipterus (Fabricius, 1803) (Hemiptera: Cimicidae), para la península Ibérica. Butlletí de la Institució Catalana d’Història Natural, 84: 289-290.
Riba-Flinch, J. M. & Goula, M. 2021. Primeras citas del tigre del laurel, Stephanitis lauri Rietschel, 2014 (Hemiptera, Heteroptera, Tingidae) para la Península Ibérica. Boletín de la Asociación española de Entomología, 45 (1-2).
Saavedra, M. C., Avila, G. A., Withers, T. M. & Holwell, G. I. 2015a. The potential global distribution of the Bronze bug Thaumastocoris peregrinus Carpintero and Dellapé (Hemiptera: Thaumastocoridae). Agricultural and Forest Entomology, 17 (4): 375-388.
Saavedra, M. C., Withers, T. M. & Holwell, G. I. 2015b. Susceptibility of four Eucalyptus host species for the development of Thaumastocoris peregrinus Carpintero and Dellapé (Hemiptera: Thaumastocoridae). Forest Ecology and Management, 336: 210-216.
Santadino, M., Brentassi, M. E., Fanello, D. D. & Coviella, C. E. 2017. First Evidence of Thaumastocoris peregrinus (Heteroptera: Thaumastocoridae) Feeding From Mesophyll of Eucalyptus Leaves. Environmental Entomology, 46 (2): 251-257.
Soliman, E. P., Wilcken, C. F., Pereira, J. M., Dias, T. K. R., Zaché, B., Dal Pogetto, M. H. F. A. & Barbosa, L. R. 2012. Biology of Thaumastocoris peregrinus in different eucalyptus species and hybrids. Phytoparasitica, 40 (3): 223-230.
Vivas, L., Crespo, J. & Jacinto, V. 2015. Primer registro de la especie invasora Thaumastocoris peregrinus Carpintero & Dellapé, 2006 en España y nuevos datos para Portugal (Hemiptera: Thaumastocoridae). BV news Publicaciones Científicas, 4 (48): 30-35.
Wilcken, C. F., Dal Pogetto, M. H. F. A., Lima, A. C. V., Soliman, E. P., Fernandes, B. V., da Silva, I. M., Zanuncio, A. J. V., Barbosa, L. R. & Zanuncio, J. C. 2019. Chemical vs entomopathogenic control of Thaumastocoris peregrinus (Hemiptera: Thaumastocoridae) via aerial application in eucalyptus plantations. Scientific Reports, 9: 9416.
Wilcken, C. F., Soliman, E. P., Nogueira de Sá, L. A., Barbosa, L. R., Dias, T. K. R., Ferreira-Filho, P. J. & Rodrigues Oliveira, R. J. 2010. Bronze bug Thaumastocoris peregrinus Carpintero and Dellapé (Hemiptera: Thaumastocoridae) on Eucalyptus in Brazil and its distribution. Journal of Plant Protection Research, 50 (2): 201-205.
58 Butlletí ICHN 85 (2), 2021
J. TOMÀS ET AL.
Butlletí de la Institució Catalana d’Història Natural, 85 (2): 59-72. 2021
Diversitat, distribució i fenologia de les cigales (Hemiptera: Cicadidae) a Catalunya (NE Península Iberica)
Pere Pons1, Rafael Carbonell Font2, Martí Franch3, Josep M. Bas1, Daniel Espejo Fraga4, Ferran Fontelles5, David Funosas6, Marc Fusellas7, Roger Puig-Gironès1, Carles Tobella1 & Marc Franch1
1 Grup de Recerca en Biologia Animal. Departament de Ciències Ambientals. Universitat de Girona. 17003 Girona
2 Can Grau. 17850 Beuda
3 Institut Català d’Ornitologia. Edifici Fòrum, Plaça Leonardo da Vinci 4-5. 08019 Barcelona
4 Can Vidaló, Freixenet de Segarra. 25271 Sant Guim de Freixenet
5 Passatge Vall Fred, 2. 25280 Solsona
6 INRAE - Unitat de recerca EABX. 50 Avenue de Verdun. 33612 Cestas, França
7 Carrer dels Estricadors, 28-30, 1r, 2na. 17820 Banyoles
Autor per a la correspondència: Pere Pons Ferran. A/e: pere.pons@udg.edu
Rebut: 12.04.2021; Acceptat: 09.06.2021; Publicat: 30.06.2021
Resum
Les cigales o cicàdids són insectes amb fase juvenil endogea, cicle vital llarg i cants audibles, importants a les xarxes tròfiques d’ecosistemes terrestres. Malgrat l’interès que han despertat arreu, la informació històrica a la península ibèrica, i en particular a Catalunya, és escassa. El catàleg d’hemípters de Martorell y Peña (1879) hi cita set espècies. Després d’aquesta data, les cigales van rebre molt poca atenció fins al segle XXI. La informació acumulada els darrers anys permet estudiar la diversitat, distribució i fenologia del grup. A tal efecte, es va realitzar una cerca bibliogràfica mitjançant cercadors acadèmics, i es van analitzar les dades del portal ornitho.cat pel període 2006-2020 (N=3468 registres). Es presenta el catàleg de cicàdids de Catalunya que conté 12 espècies, un nombre comparable al d’altres territoris europeus: les cigales grossa, del pi, de l’olivera i negra, respectivament Lyristes plebejus Scopoli, 1763, Cicada orni Linné, 1758, C. barbara Stal, 1866 (al·lòctona introduïda) i Cicadatra atra Olivier, 1790; les brunzidores ala-roja, sigiŀlada, de garriga i cotonosa, respectivament Tibicina haematodes Scopoli, 1763, T. quadrisignata Hagen, 1855, T. garricola Boulard, 1983 i T. tomentosa Olivier, 1790; i les cigalelles europea occidental, vespera, culprima i argentada, respectivament Cicadetta petryi Schumacher, 1924, Hilaphura varipes Waltl, 1837, Euryphara dubia Rambur, 1840 i Tettigettalna argentata Olivier, 1790. Es mostren mapes de riquesa i distribució d’espècies en quadrícules UTM de 5 × 5 km, diagrames de distribució altitudinal i fenologia per les espècies més comunes, així com iŀlustracions originals de suport a la identificació. Aquest treball iŀlustra el potencial de la ciència ciutadana per ampliar el coneixement de grups d’insectes poc estudiats i estableix un escenari de referència sobre el que fonamentar futurs estudis de la distribució i ecologia dels cicàdids.
Paraules clau: Cicadidae, Catalunya, Península Ibèrica, ciència ciutadana, dades històriques, distribució, fenologia, catàleg d’espècies.
Abstract
Diversity, distribution and phenology of cicadas (Hemiptera: Cicadidae) in Catalonia (NE Iberian Peninsula)
True cicadas (Cicadidae) are insects with a juvenile phase spent belowground, a long life cycle and audible songs, that are important in the trophic networks of terrestrial ecosystems. Despite the interest they arouse, historical information in the Iberian Peninsula, and more specifically in Catalonia, is scarce. The catalogue of Hemipteran by Martorell y Peña (1879) cited seven species. After that date, cicadas received very little attention until the 21st century. The information accumulated in recent years allowed us to study the diversity, distribution and phenology of the group. To this end, a bibliographic search was carried out using the academic search engines, and the data hosted at ornitho.cat website was analysed for the period 2006-2020 (N= 3468 records). The catalogue of cicadas of Catalonia presented here contains 12 species, a number comparable to that of other European territories: Cicadinae Lyristes plebejus Scopoli, 1763, Cicada orni Linné, 1758, C. barbara Stal, 1866 (introduced non-native) and Cicadatra atra Olivier, 1790; Tibicininae Tibicina haematodes Scopoli, 1763, T. quadrisignata Hagen, 1855, T. garricola Boulard, 1983 and T. tomentosa Olivier, 1790; and Cicadettinae Cicadetta petryi Schumacher, 1924, Hilaphura varipes Waltl, 1837, Euryphara dubia Rambur, 1840 and Tettigettalna argentata Olivier, 1790. We provide species richness and distribution maps in 5 × 5 km UTM grids, altitudinal distribution and phenology diagrams for the most common species, as well as original illustrations to support identification. This work shows the potential of citizen science to expand the knowledge of a scarcely studied group of insects and establishes a baseline scenario on which to base future studies of the distribution and ecology of cicadas.
Key words: Cicadidae, Catalonia, Iberian Peninsula, citizen science, historical data, distribution, phenology, species catalogue.
ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition) DOI:
59 Butlletí ICHN 85 (2), 2021 GEA, FLORA ET FAUNA GEA, FLORA ET FAUNA
10.2436/20.1502.01.74
Introducció
Les cigales o cicàdids (Hemiptera: Cicadidae) són insectes presents a tots els continents, excepte a l’Antàrtida, i a la major part de grans illes (Marshall et al., 2018). Actualment es reconeixen almenys cinc subfamílies de cicàdids: Cicadinae, Cicadettinae, Tibicininae, Tettigomyiinae i Derotettiginae (Marshall et al., 2018; Simon et al., 2019). D’aquestes, les tres primeres són presents a la regió Paleàrtica. Els cicàdids presenten una llarga fase juvenil endogea (en galeries que excaven al sòl) i una curta fase adulta epigea (sobre la vegetació), durant la qual es fan notoris a molts ecosistemes terrestres on dominen el paisatge sonor durant les hores de més calor (Farina et al., 2011). Es tracta d’un grup taxonòmic que ha encuriosit els investigadors pel seu peculiar cicle vital, dels més llargs entre els insectes no socials, especialment les cigales periòdiques nord-americanes que arriben a 17 anys de vida (Williams & Simon, 1995). També és peculiar la seva alimentació a base de la poc nutritiva saba del xilema, que obtenen a partir d’arrels quan són nimfes, i de tiges o branques quan són adultes (Boulard & Mondon, 1995). Altres aspectes d’estudi han estat l’aparell sonor i el cant dels mascles, que s’utilitza en la sistemàtica dels Cicadidae, en revisió actualment (Marshall et al., 2018; Gogala, 2019). Es tracta, concretament, d’un grup que ha divergit més en senyals acústics que en morfologia externa (Pinto-Juma et al., 2009). Les cigales han atret també l’atenció per la seva abundància i importància en la xarxa tròfica dels boscos (Koenig & Liebhold, 2005), així com per la seva afectació a certs conreus, com l’olivera o la vinya (Mehdipour et al., 2016). En l’àmbit de la tecnologia, la nanoestructura de les seves ales s’ha utilitzat com a model per generar superfícies amb propietats bactericides, entre altres (Lin et al., 2018). Finalment, en l’àmbit social, són font d’aliment i de mitologia a moltes cultures (Srivastava et al., 2009). No és estrany que els cicàdids hagin estat objecte de nombrosos estudis arreu del món. A Europa, concretament, hi ha una llarga tradició d’estudi de la biologia, filogènia i bioacústica d’aquest grup, del qual a data de 2021 se’n coneixen almenys 71 espècies: 15 Cicadinae, 8 Tibicininae i 48 Cicadettinae (Gogala, 2019). El sud d’Espanya, Grècia i Europa Oriental apareixen com a punts calents (hotspots) de biodiversitat de cicàdids, on s’hi han descrit noves espècies i subespècies de Cicadinae i Cicadettinae durant els darrers anys (Gogala et al., 2005; Puissant & Sueur, 2010; Simoes et al., 2013). Menys diverses, tanmateix, les illes de la Mediterrània Occidental estan proporcionant noves espècies de Tibicininae (Hertach, 2020).
Amb aquests antecedents, és sorprenent l’escassa atenció que tradicionalment han rebut les cigales per part de naturalistes, zoòlegs i ecòlegs a la península Ibèrica, i en particular a Catalunya. Les observacions de cicàdids han estat sempre associades a treballs generals sobre el conjunt de l’entomofauna, sobre els Hemípters o, com a molt, sobre els Homòpters (grup parafilètic). El 1879 es va publicar un catàleg d’hemípters (Martorell i Peña, 1879) en base a la coŀlecció d’insectes conservada al Museu de Ciències Naturals de Barcelona i, segons el seu autor, era la «primera vegada que els hemípters
de Catalunya són presos en consideració, amb algun ordre, pels naturalistes Catalans». Aquest catàleg contenia set espècies de cicàdids: Cicada orni Linné, 1758 (citada com a Tettigia orni), Lyristes plebejus Scopoli, 1763 (com a Cicada plebeja), Cicadatra atra Olivier, 1790, Tibicina hematodes Scopoli, 1763, Cicadetta sp. (com a Cicadetta montana Scopoli 1772), Tettigettalna argentata Olivier, 1790 (com a Cicadetta argentata), així com Cicadetta flaveola Brull, 1832 que resultaria ser erròniament determinada. D’altra banda, als tres catàlegs entomològics publicats per Cuní i Martorell (Cuní i Martorell, 1883, 1885, 1888) apareixen tan sols dues espècies de cicàdids, de les més abundants i ja citades a l’obra del 1879. Cal esperar quatre dècades (Codina, 1920) per veure tres espècies d’aquest grup no citades anteriorment a Catalunya: Hilaphura varipes Waltl, 1837 (com a Melampsalta picta Germar 1830), Tibicina quadrisignata Hagen, 1855 (com a Tibicen quadrisignata) i Oligoglena tibialis Panzer, 1798 (com a Melampsalta (Pauropsalta) tibialis). Aquesta darrera espècie va ser fruit d’una classificació incorrecta, com s’argumentarà a la Discussió. Una dècada després, Melampsalta montana (ara s’ha de considerar com a Cicadetta sp., de determinació específica incerta) és citada a la Vall de la Noguera de Cardós (Navàs, 1930).
No és fins l’any 1957 quan apareix finalment una monografia de cicàdids amb informació per Catalunya, en una obra que abastava el conjunt d’Espanya (Gómez-Menor, 1957). Aquesta publicació recollia les observacions dels treballs anteriors de Martorell i Peña (1879) i de Codina (1920), i no aportava novetats pel territori català. La web del projecte «Fauna Ibérica» descriu la diversitat dels Cicadoidea, però utilitzant una taxonomia i un llistat d’espècies no actualitzats (Alonso-Zarazaga, 2002). D’entre les escasses publicacions recents que mencionen cigales a Catalunya, hi ha una guia sonora d’insectes que conté enregistraments de cinc taxons de cigales (Carbonell & Matheu, 2010) i un treball sobre els invertebrats del massís del Mont a l’Alta Garrotxa que cita L. plebejus i C. orni (Carbonell, 2011).
La primera publicació científica específica d’aquest grup a Catalunya no apareix fins al 2015 i tracta sobre l’efecte de l’incendi i la tala en l’abundància de C. orni (Pons, 2015). A partir del 2014 el coŀlectiu cicada.cat, impulsat pel Grup de Recerca en Biologia Animal de la Universitat de Girona, recull informació sobre les espècies de cicàdids presents a Catalunya, la seva distribució i ecologia i la divulga mitjançant una web a tal efecte (GR-Biologia Animal, 2021). A partir del 2015 la plataforma de biodiversitat ornitho.cat (www. ornitho.cat) inclou els Cicadidae entre els grups taxonòmics dels quals hom pot entrar informació georeferenciada. Altres plataformes com iNaturalist (www.inaturalist.org) i Natusfera (https://natusfera.gbif.es) també permeten aquesta possibilitat però el volum de dades, i les capacitats de consulta i verificació de les mateixes són menors.
Els recents canvis taxonòmics, que afecten sobretot als Cicadettinae, influeixen en el nombre d’espècies del territori europeu. En el cas de Catalunya, els exemplars recoŀlectats del gènere Cicadetta Kolenati, 1857, s’havien determinat com a C. montana. Aquesta denominació abastava en realitat un complex d’almenys 13 espècies, distingibles fona-
60 Butlletí ICHN 85 (2), 2021
FLORA ET FAUNA P. PONS ET AL.
GEA,
mentalment per tècniques bioacústiques i moleculars. Amb la nomenclatura actual, les formes del gènere que, per distribució, podrien probablement trobar-se a Catalunya són C. petryi Schumacher, 1924, C. brevipennis litoralis Puissant & Hertach, 2016 i C. cerdaniensis Puissant & Boulard, 2000 (Hertach et al., 2016). Per tant, aquest és un dels gèneres que convé estudiar preferentment.
Per tal d’afavorir l’interès ciutadà per les cigales es va treballar en una proposta de noms catalans pels diferents nivells taxonòmics. La materialització d’aquests noms, majoritàriament neologismes, va ser possible l’any 2019, fruit d’una coŀlaboració del coŀlectiu cicada.cat amb el Termcat. A banda del nom ja existent de cigales pels Cicadidae, es va decidir un nom per designar cada subfamília: cigales específicament pels més comuns i audibles Cicadinae, brunzidores pels Tibicininae i cigalelles pels Cicadettinae. Els noms catalans de les espècies, presents o potencialment presents a Catalunya, apareixen a la Taula 1 i es poden consultar online al Cercaterm i a la Neoloteca (Termcat, 2021), amb la seva justificació etimològica. Així mateix, han estat objecte d’una infografia expressa per difondre’ls.
Amb els antecedents esmentats, els objectius del present article són: (1) descriure els canvis històrics en el coneixement de la diversitat de cicàdids a Catalunya; (2) sintetitzar la informació disponible sobre la diversitat, distribució, fenologia i ecologia d’aquest grup d’hemípters a Catalunya, per realitzar un catàleg actualitzat d’espècies, iŀlustracions de les mateixes, mapes de distribució i diagrames fenològics i altitudinals; i (3) discutir aquest coneixement i presentar propostes per futurs estudis.
Material i mètodes
Per tal de descriure l’evolució històrica del coneixement dels cicàdids a Catalunya, s’ha realitzat una cerca bibliogràfica utilitzant els cercadors acadèmics habituals (Web of Science, Scopus, Google Scholar). S’ha accedit a les obres més antigues mercès a la digitalització realitzada per la Institució Catalana d’Història Natural, El Real Jardín Botánico de Madrid i la Biblioteca Digital Hispánica. Hem consultat treballs generals sobre Espanya i França, i específicament sobre Catalunya i les regions veïnes d’Aragó, País Valencià i Languedoc-Roussillon, així com treballs d’àmbit local. Per comprovar que no s’ha omès informació rellevant, s’ha realitzat també cerques selectives de cites, imatges i enregistraments de cigales a internet, incloent-hi les plataformes de biodiversitat Biodiversidad Virtual (www.biodiversidadvirtual. org/taxofoto), Natusfera i Banc de Dades de Biodiversitat de Catalunya (http://biodiver.bio.ub.es/biocat/index.jsp)
Pel que fa a coŀleccions, es van revisar tots els Cicadidae de localitat catalana conservats al Museu de Ciències Naturals de Barcelona (MCNB). La coŀlecció conté gairebé un centenar d’exemplars dels segles XIX, XX i XXI recollectats a Catalunya, alguns d’ells citats en publicacions històriques.
La part principal d’aquest treball s’ha basat en les dades de cicàdids contingudes a ornitho.cat. Aquest és un portal web
de ciència ciutadana impulsat per l’Institut Català d’Ornitologia i dedicat a l’intercanvi d’informació sobre observacions de biodiversitat, especialment animal, de Catalunya. Encara que les cigales es van incloure al portal l’any 2015, s’hi han entrat dades des del 2006, i anecdòticament abans (de 0 a 3 registres/any entre 1999 i 2005). Entre 2006 i 2014 el nombre de registres (definits com a cites d’un taxó a una localitat i data, independentment del nombre d’individus) per any va osciŀlar entre 33 i 118, mentre que del 2015 al 2020 ha osciŀlat entre 255 i 937 (Fig.1; Taula 2). Les dades passen un procés de verificació per part membres de cicada. cat que actuen com a revisors habituals. Per considerar vàlids certs registres, han d’anar acompanyats de fotografia, enregistrament sonor o captura d’un exemplar. Aquest fet depèn del gènere, localitat i data de l’observació, i en cas de dubte es demana a l’autor més detalls sobre l’observació. En diverses ocasions ens hem desplaçat al lloc de l’observació per intentar fotografiar, enregistrar o capturar-ne un exemplar. Els membres del coŀlectiu cicada.cat han realitzat també campanyes específiques de prospecció de cicàdids, entrant també les dades més rellevants a ornitho.cat. Els exemplars capturats (habitualment un sol mascle de cada espècie per minimitzar l’impacte de la captura) es conserven a la col·lecció de referència de la Universitat de Girona. En els casos més complexes, s’ha comptat amb l’assessorament dels especialistes Stéphane Puissant, del Muséum National d’Histoire Naturelle, i Matija Gogala del Slovenian Museum of Natural History, per confirmar o determinar un exemplar.
Els registres d’ornitho.cat (Taula 2), convenientment filtrats per minimitzar errades, s’han utilitzat per generar cartografia i gràfics. S’han realitzat mapes de presència de les espècies en les quadrícules UTM de 5 × 5 km de Catalunya. En la interpretació dels mateixos, cal tenir en compte que el mostreig no ha estat extensiu ni estandarditzat, i que la resolució de les dades originals és d’un km2. Amb els mateixos supòsits s’han elaborat diagrames de fenologia i distribució altitudinal per aquelles espècies amb més de 20 registres acumulats.
Finalment, les iŀlustracions de les espècies de cigales es van elaborar en base a fotografies de camp sempre que ha estat possible, per tenir referències de la posició i coloració en estat natural. Es van utilitzar fotografies de coŀlecció i els
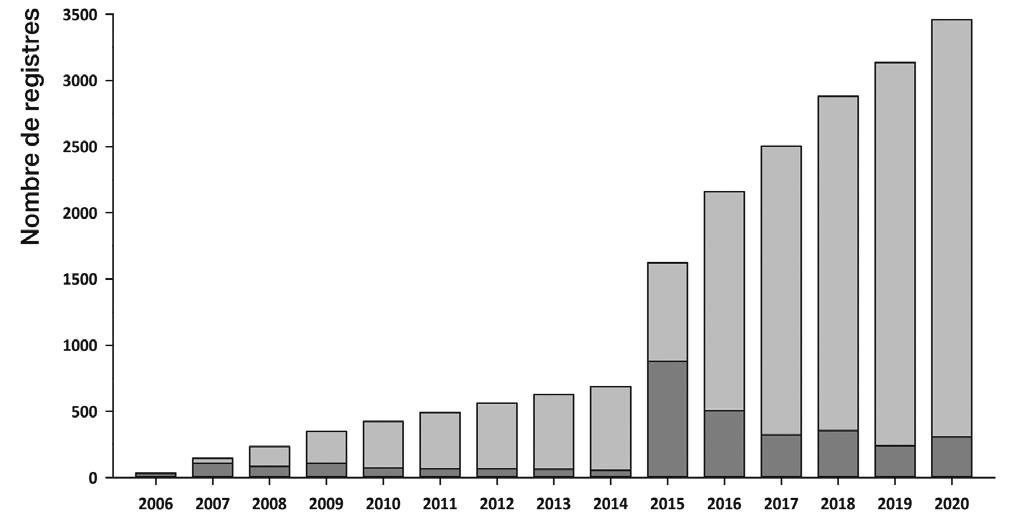
61 Butlletí ICHN 85 (2), 2021 GEA, FLORA ET FAUNA DIVERSITAT, DISTRIBUCIÓ I FENOLOGIA DE LES CIGALES (HEMIPTERA: CICADIDAE) A CATALUNYA
Figura 1. Nombre de registres anuals (gris fosc) i acumulats (gris clar) de cigales al portal ornitho.cat (2006-2020).
GEA, FLORA ET FAUNA
exemplars de la coŀlecció de referència de la Universitat de Girona per als detalls morfològics. Totes les espècies es van iŀlustrar en visió dors-lateral per tal de mostrar de manera comparable els caràcters visibles en observacions o fotografies de camp, i es van dibuixar les ales per mostrar caràcters diagnòstics de les nervacions alars.
Resultats
A Catalunya han estat citades amb certesa 12 espècies de cigales fins l’actualitat (Taula 1): les cigales grossa, del pi, de l’olivera i negra, respectivament Lyristes plebejus, Cicada orni, C. barbara Stal, 1866 (aŀlòctona introduïda) i Cicadatra atra; les brunzidores ala-roja, sigiŀlada, de garriga i cotonosa, respectivament Tibicina haematodes, T. quadrisignata, T. garricola Boulard, 1983 i T. tomentosa Olivier, 1790; i les cigalelles europea occidental, vespera, culprima i argentada, respectivament Cicadetta petryi, Hilaphura varipes, Euryphara dubia Rambur, 1840 i Tettigettalna argentata. El coneixement de la diversitat del grup ha evolucionat lentament fins ben entrat el segle XXI. En els anys compresos entre 2012 i 2018 l’activitat del col·lectiu cicada.cat i la ciència ciutadana han permès localitzar espècies mai abans citades a Catalunya i redescobrir-ne les que s’havien citat molts anys abans. La immensa majoria de cites recents de Cicàdids a Catalunya estan recollides a ornitho.cat (N = 3468 registres), mentre que la resta de portals consultats contenen escassos registres del grup.
Noves cites d’espècies a Catalunya
Aquest treball aporta cinc espècies noves per Catalunya (Taula 1): C. barbara, T. garricola, T. tomentosa, C. petryi i E. dubia. La cigala de l’olivera C. barbara es va detectar per primera vegada a Catalunya l’estiu del 2017 dins la finca d’una distribuïdora d’oliveres, a cavall entre la Selva i el Vallès Oriental. Aquesta cigala es detecta i determina fàcilment pel seu cant continu, similar al so d’un drone, encara que per morfologia externa és molt similar a la cigala del pi. Des d’aleshores s’han detectat cada estiu a la mateixa finca i entorns, fins a un màxim d’uns 15 mascles cantant. Ha estat citada també al Maresme i al Barcelonès, dins un viver de planta ornamental amb oliveres i un parc urbà, respectivament. Com es discutirà més endavant, es considera una espècie d’introducció recent.
Pel que fa a les noves observacions de brunzidores, Tibicina spp., es van determinar a partir de la morfologia externa i de l’anàlisi del sonograma del cant. La brunzidora de garriga T. garricola es va detectar per primer cop al Vallès Occidental el 2015 i posteriorment s’ha citat en sis comarques més (Alt Empordà, Anoia, Bages, Garrotxa, Pallars Jussà i Segarra) La brunzidora cotonosa T. tomentosa es va detectar inicialment al Segrià el 2013 i després a la Segarra i l’Alt Empordà. Ambdues espècies coincideixen a les dues darreres comarques, encara que en localitats i hàbitats diferents (Fig. 5).
L’any 2012 es va confirmar la presència de C. petryi al Priorat mitjançant l’anàlisi del seu sonograma (determinació
Taula 1. Espècies i subespècies de Cicadidae trobades a Catalunya, potencialment presents i citades erròniament. Es detallen els noms científics i els seus autors, el nom català, l’any de la primera cita a Catalunya i els noms de les espècies a Martorell i Peña (1879) i a Codina (1920). Els dos anys entre parèntesi es refereixen a cites actualment considerades errònies. Els gèneres Cicada, Lyristes i Cicadatra pertanyen als Cicadinae, Tibicina als Tibicininae i la resta de gèneres als Cicadettinae.
Nom científic Autors
Tàxons citats a Catalunya
Cicada orni Linné, 1758
Cicada barbara Stal, 1866
Nom català
1a cita Sinònim 1879 Sinònim 1920
Cigala del pi 1879 Tettigia orni
Cigala de l’olivera 2017
Lyristes plebejus Scopoli, 1763 Cigala grossa 1879
Cicadatra atra Olivier, 1790
Tibicina garricola Boulard, 1983
Tibicina haematodes Scopoli, 1763
Tibicina quadrisignata Hagen, 1855
Tibicina tomentosa
Olivier, 1790
Cicadetta petryi Schumacher, 1924
Tettigettalna argentata
Olivier, 1790
Hilaphura varipes Waltl, 1837
Euryphara dubia Rambur, 1840
Tàxons potencialment presents a Catalunya
Tibicina corsica fairmairei Boulard, 1983
Cigala negra 1879
Brunzidora de garriga 2015
Brunzidora ala-roja 1879
Brunzidora sigiŀlada 1920
Brunzidora cotonosa 2013
Cigalella europea occidental 2012
Cigalella argentada 1879
Cigalella vespera 1920
Cigalella culprima 2018
Brunzidora d’Occitània
Tettigettula pygmea Olivier, 1790 Cigalella menuda
Cicadetta brevipennis litoralis Puissant & Hertach, 2016 Cigalella costanera
Cicadetta cerdaniensis Puissant & Boulard, 2000 Cigalella pirinenca
Tàxons amb cites a Catalunya actualment considerades errònies
Cicada plebeja
Cicadatra atra
Tibicina haematodes
Cicadetta montana
Cicadetta argentata
Cicada orni
Cicada barbara
Lyristes plebeja
Cicadatra atra
Tibicen haematodes
Tibicen quadrisignata
Tibicen tomentosa
Melampsalta montana
Melampsalta argentata
Melampsalta picta
Melampsalta dubia
Oligoglena flaveola Brull, 1832 (1879) Cicadetta flaveola
Oligoglena tibialis Panzer, 1798 (1920)
Melampsalta flaveola
Melampsalta (Pauropsalta) tibialis
62 Butlletí ICHN 85 (2), 2021
P. PONS ET AL.
DIVERSITAT, DISTRIBUCIÓ I FENOLOGIA DE LES CIGALES (HEMIPTERA: CICADIDAE) A CATALUNYA
63 Butlletí ICHN 85 (2), 2021 GEA, FLORA ET FAUNA Nom català Nom científic 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012 2013 2014 2015 2016 2017 2018 2019 2020 ∑ Cigales (Cicadinae) Cigala del pi Cicada orni 0 0 0 0 0 1 2 14 39 36 51 40 31 39 38 27 508 268 130 144 116 145 1629 Cigala de l’olivera Cicada barbara 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 5 4 2 1 12 Cigala grossa Lyristes plebejus 1 0 0 1 0 0 1 6 18 15 17 13 15 13 14 5 107 53 93 39 30 35 476 Cigala negra Cicadatra atra 0 0 0 0 0 0 0 0 0 0 10 0 1 0 2 12 110 42 34 39 25 40 315 Cigala indeterminada Cicada sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 3 3 2 9 Brunzidores (Tibicininae) Brunzidora de garriga Tibicina garricola 0 0 0 0 0 0 0 0 0 0 0 0 0 2 0 0 12 12 4 5 0 3 38 Brunzidora ala-roja Tibicina haematodes 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 5 6 1 4 7 1 24 Brunzidora sigiŀlada Tibicina quadrisignata 0 0 0 0 0 0 0 0 0 0 0 0 0 0 5 0 0 2 0 5 8 1 21 Brunzidora cotonosa Tibicina tomentosa 0 0 0 0 0 0 0 0 0 0 0 0 0 0 2 0 0 0 1 6 0 0 9 Brunzidora indeterminada Tibicina sp. 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 1 3 7 2 1 2 5 22 Cigalelles (Cicadettinae) Cigalella europea occidental Cicadetta petryi 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 1 1 0 3 Cigalella argentada Tettigettalna argentata 0 0 0 0 0 0 0 13 49 30 38 22 21 14 5 12 185 145 73 114 56 84 861 Cigalella vespera Hilaphura varipes 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 5 0 0 5 0 4 14 Cigalella culprima Euryphara dubia 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 2 0 0 2 Cigalella indeterminada Cicadetta sp. 0 0 0 0 0 0 0 0 7 6 1 0 0 0 1 0 2 1 0 4 1 1 24 Cigalella indeterminada Cicadetta / Tettigettalna / Tettigettula sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 1 3 Cigala indeterminada Cicadidae sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 4 2 6 1 0 0 1 0 1 3 33 113 87 118 75 68 69 67 57 937 538 343 377 255 325 3468 Taula 2. Registres per taxó i any dels Cicadidae allotjats del portal ornitho.cat , que configuren la
dades utilitzada
base de
al present estudi.
GEA,
de S. Puissant), ja que els exemplars citats com a C. montana a una localitat desconeguda (Martorell i Peña, 1879), i com a Melampsalta montana a la Serra del Vert (Gómez-Menor, 1957), corresponen a un taxó que, com s’ha dit anteriorment, s’ha escindit en diverses espècies.
Finalment, un mascle de cigalella culprima E. dubia va ser recoŀlectat al Solsonès el juny de 2018 i posteriorment determinat per S. Puissant en base a la genitàlia. L’espècie no
s’ha retrobat des d’aleshores. Tanmateix, a manca d’anàlisi de la genitàlia, no es descarta que un exemplar conservat al MCNB pugui pertànyer a aquest taxó.
Altres cites destacables
A més de les espècies noves per Catalunya, és destacable la redescoberta de la cigalella vespera, H. varipes, considerada un endemisme ibèric (Puissant, 2005). L’espècie havia
64 Butlletí ICHN 85 (2), 2021
FLORA
ET FAUNA
Figura 2. Espècies de cigales trobades a Catalunya o possiblement presents, dibuixades a escala. Dels gèneres Cicada i Cicadetta, que contenen espècies difícilment distingibles per morfologia externa, se n’iŀlustra un sol representant. Es representa un mascle de cada espècie. De la cigala negra se n’iŀlustren la forma nominal (negra) i la forma flava (groga).
P. PONS ET AL.
DIVERSITAT,
a les ales anteriors.
estat citada fa un segle a la Segarra (Codina, 1920). Posteriorment se la menciona al Segrià (Gómez-Menor, 1957), tot i que aquestes cites les atribueix l’autor a Codina i la data de captura no s’esmenta. En el marc d’aquest estudi es va retrobar el 2015 a l’Anoia i la Segarra, i en anys posteriors a les Garrigues, el Priorat i l’Urgell.
D’altra banda, vam poder identificar dos exemplars antics de T. haematodes al MCNB: un sense determinar, recol·lectat
a Sarroca de Lleida el juny de 1933, i un altre del Montsant datat el 1918, determinat per A. Codina com a T. quadrisignata. Ambdues espècies es distingeixen entre elles per la coloració de les nervacions alars. Aquests exemplars de museu suposen, fins ara, les úniques cites de la brunzidora ala-roja al Segrià i al Priorat. Finalment, els 7 exemplars del MCNB recoŀlectats durant la primera meitat del segle XX i determinats com a Cicadetta flaveola provenen d’Andalusia
65 Butlletí ICHN 85 (2), 2021 GEA, FLORA ET FAUNA
Figura 3. Ales de les espècies de cigales trobades a Catalunya o possiblement presents, per ressaltar-ne les nervacions. Dibuix a escala. Noti’s que les cigales del pi i de l’olivera es poden distingir pel nombre i aparença de les taques grans de les nervacions en ziga-zaga
DISTRIBUCIÓ I FENOLOGIA DE LES CIGALES (HEMIPTERA: CICADIDAE) A CATALUNYA
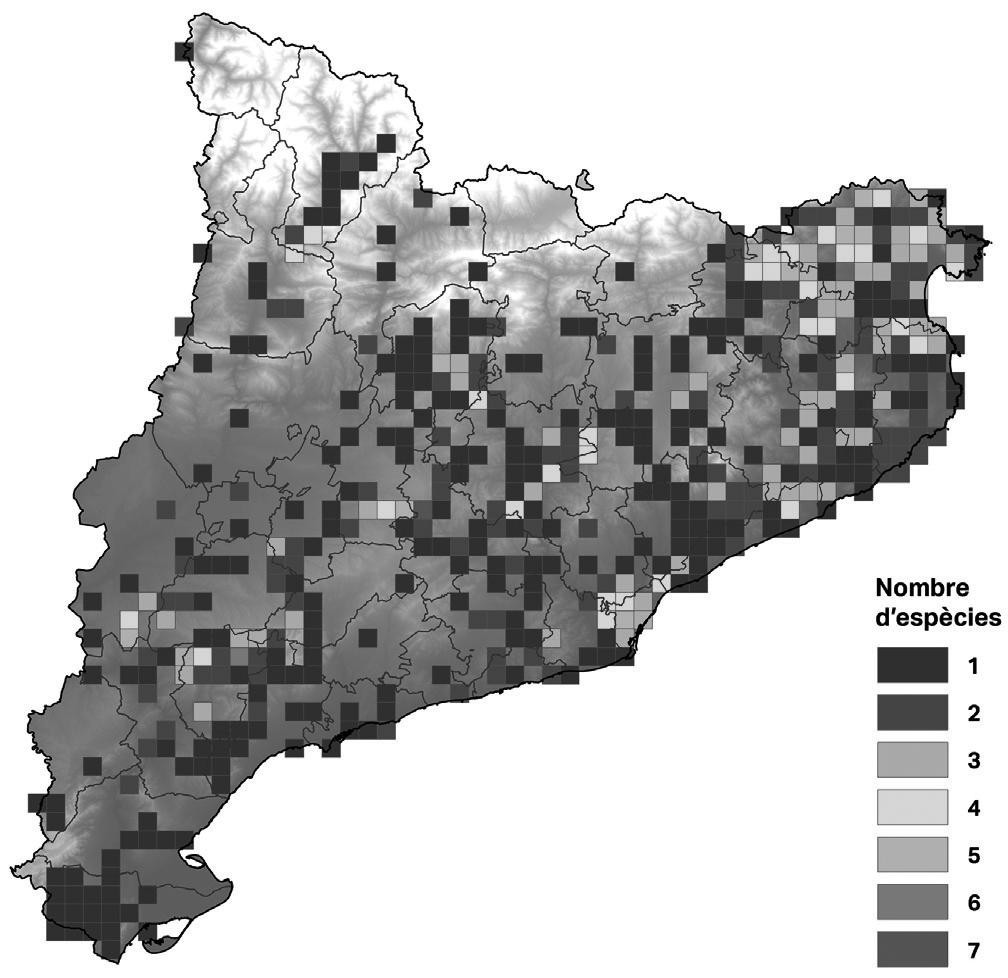
Figura 4. Riquesa d’espècies de Cicadidae a Catalunya en base a les dades recollides al portal ornitho.cat (2006-2020). Es mostren el nombre d’espècies de les quadrícules UTM 5 × 5 km amb registres i els límits comarcals.
i Navarra. Per tant, no s’han pogut estudiar el/s exemplar/s suposadament recoŀlectats a Catalunya, citats per Martorell i Peña (1879).
Iŀlustracions, distribució, rang altitudinal i fenologia
La Figura 2 mostra les iŀlustracions de mascles de les espècies de cigales trobades a Catalunya o potencialment presents per la seva distribució en regions veïnes. L’habitus, la coloració i la mida són rellevants per la determinació de bona part dels taxons. Tanmateix, en altres casos cal recórrer a la disposició i coloració de les nervacions alars, així com la presència de taques a les ales, com s’iŀlustra a la Figura 3.
La riquesa del grup, en base a les dades del portal ornitho. cat pel període 2006-20, va osciŀlar entre zero i set espècies per quadrícula UTM 5 × 5 km (Fig. 4). La majoria de quadrícules on s’han citat cigales contenen entre una i tres espècies. Amb el coneixement actual, les quadrícules més diverses (N ≥ 4 espècies) es localitzen al litoral i prelitoral, Prepirineu i relleus situats al voltant de la depressió central; (sobretot a les comarques de l’Alt i Baix Empordà, Garrotxa, Barcelonès, Solsonès, Anoia, Segarra, Priorat i Segrià).
Els mapes de distribució d’espècies per quadrícula UTM 5 × 5 km mostren que les Cicadinae, especialment la cigala del pi, són les més extensament repartides (Fig. 5). Les cigales grossa i negra es localitzen majoritàriament a la Catalunya humida, tot i que aquest patró deriva en part del volum de dades de cada regió. Els buits d’informació són més acusats per Cicadettinae i Tibicinae, insectes de cant menys audible, així com per comarques de la meitat oest de Catalunya i pirinenques. Les brunzidores presenten una distribució fragmentada en escasses localitats. L’ala-roja s’ha trobat a la Catalunya central, la sigiŀlada a l’Albera i Pirineu occidental,
la de garriga a 12 quadrícules des del prelitoral fins l’interior i Prepirineu, i la cotonosa en àrees de la Depressió Central i Aiguamolls de l’Alt Empordà. Pel que fa a les cigalelles, no hi ha encara prou informació per determinar l’abast de la distribució de l’europea occidental i la culprima. La vespera es localitza a una àrea compacta del sud de la Depressió Central. Finalment, l’argentada s’ha localitzat a bona part del territori a excepció del Pirineu axial, sectors de l’interior de Lleida i Terres de l’Ebre.
La majoria d’espècies localitzades fins ara a Catalunya són de caràcter mediterrani i els seus registres es localitzen més aviat per sota dels 800 m d’altitud (Fig. 6). Tanmateix, bona part d’espècies s’han trobat per sobre dels 1000 m d’altitud. Destaca, en aquest sentit el caràcter més muntà de la cigalella argentada, la brunzidora sigiŀlada i les cigalelles Cicadetta spp.
La fase adulta d’aquests insectes a Catalunya apareix de finals de maig al setembre, anecdòticament a l’octubre en el cas de la cigala del pi. El període de major abundància i riquesa d’espècies abasta de juny a agost i totes les espècies presenten el pic de registres al juliol (Fig. 7). Les espècies amb major nombre d’observacions mostren, lògicament, una fenologia dels adults més perllongada, especialment la cigalella argentada. A l’altre extrem, la brunzidora de garriga només ha estat detectada als mesos de juny i juliol.
Discussió
El coneixement sobre els Cicadidae a Catalunya ha estat escàs fins fa pocs anys, probablement per un conjunt de motius. El primer és l’escassetat d’especialistes en hemípters posteriors a Ascensi Codina, almenys fins a finals del segle XX. Encara que la informació disponible ha millorat darrerament pels heteròpters (Goula et al., 2010 i treballs posteriors), no ho ha fet per altres grups d’hemípters. Un altre motiu és, probablement, la curta durada de la fase adulta en les cigales. Aquestes no són habitualment capturades pels mètodes estàtics de mostreig d’insectes voladors i cal recórrer a captura activa amb caçapapallones, que no acostuma a ser senzilla ja que sovint es troben amagades per la vegetació. Un repte addicional per l’estudi de les cigales, encara que no exclusiu d’aquest grup, és l’existència d’espècies críptiques que cal diferenciar a través d’informació i de tècniques bioacústiques o moleculars (Hertach et al., 2016), que no han estat disponibles fins recentment. D’altra banda, el fet que la taxonomia hagi estat confusa a Europa durant molt temps, tampoc ha ajudat a l’estudi d’aquests insectes. Finalment, tot i considerar que les referències més importants han estat localitzades en aquest treball, probablement hi hagi altres estudis que citen cicàdids a Catalunya. Per tant, la tasca de recopilar estudis històrics i literatura grisa entomològica ha de continuar.
El treball efectuat els darrers anys en el marc de cicada. cat ha permès conèixer millor la diversitat de cigales presents a Catalunya. És destacable la descoberta de la cigalella culprima E. dubia, probable endemisme ibèric (Aguin-Pombo et al., 2007), les localitats conegudes de la qual més properes a
66 Butlletí ICHN 85 (2), 2021 GEA, FLORA ET FAUNA
P. PONS ET AL.
DIVERSITAT,
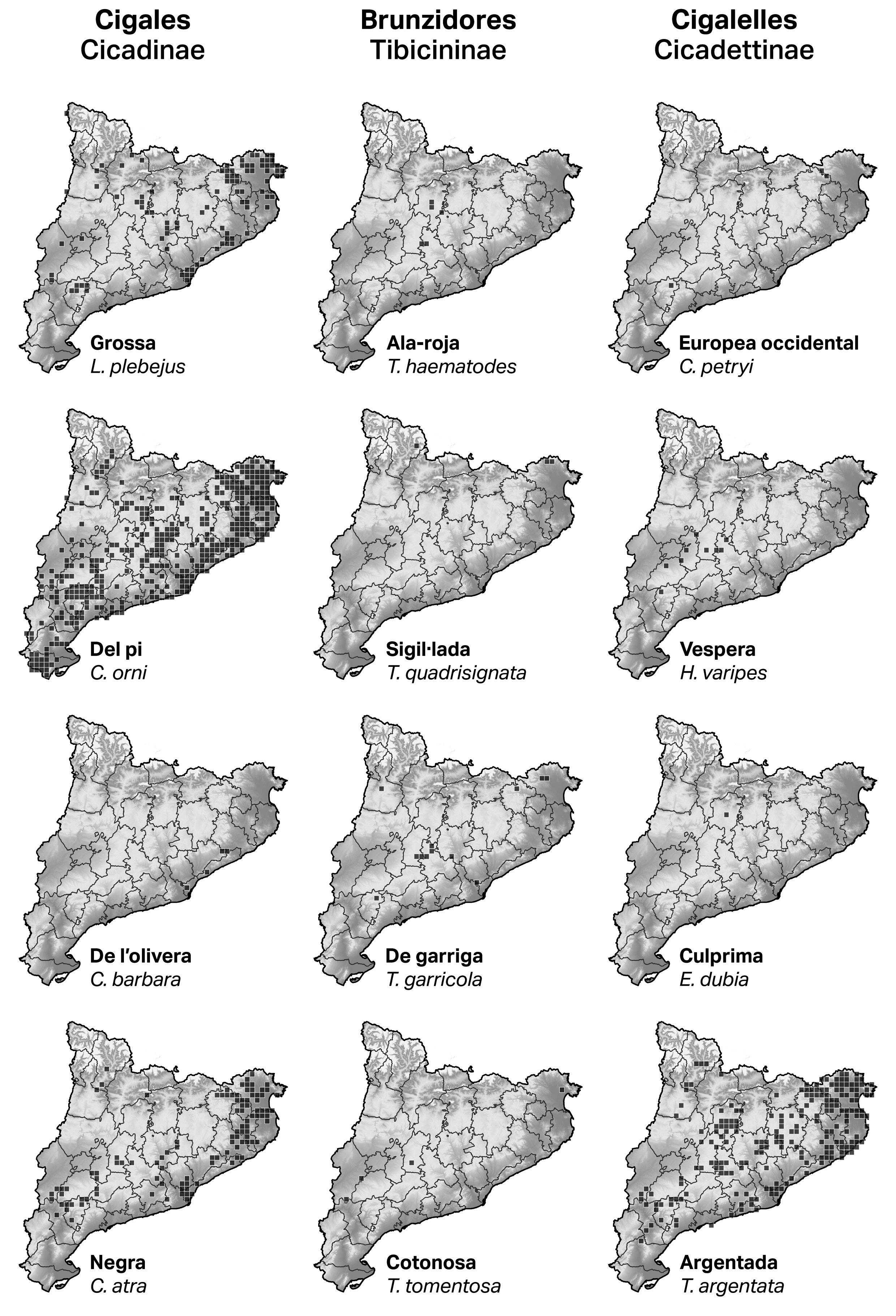
Figura 5. Distribució a Catalunya de les 12 espècies de cigales citades amb certesa en base a les dades recollides al portal ornitho.cat (2006-2020). Es mostra la presència a les quadrícules UTM 5 × 5 km sobre el fons altimètric i els límits comarcals.
67 Butlletí ICHN 85 (2), 2021
GEA, FLORA ET FAUNA
FENOLOGIA
LES CIGALES
DISTRIBUCIÓ I
DE
(HEMIPTERA: CICADIDAE) A CATALUNYA
GEA,
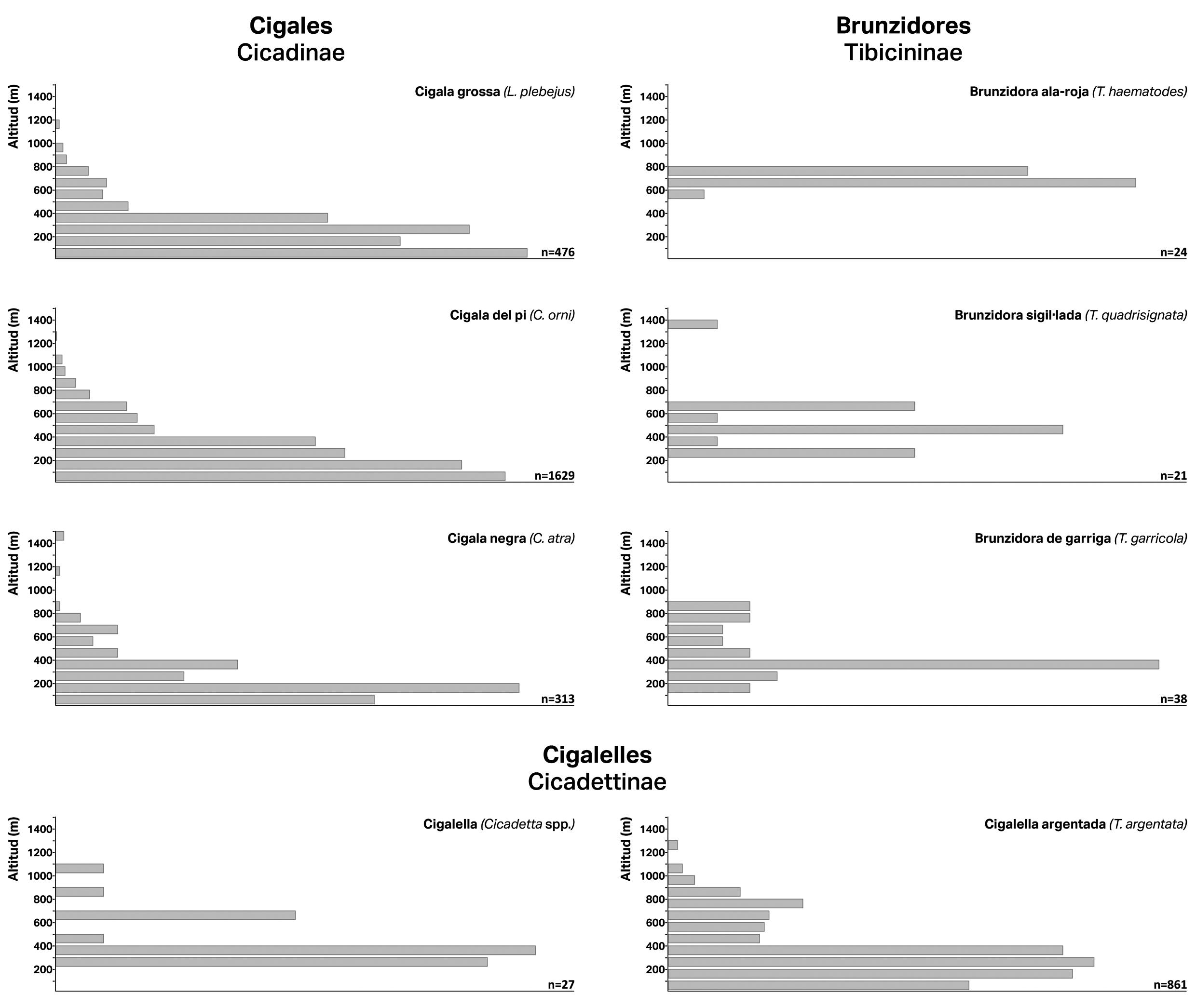
Catalunya són Tiermas (Saragossa) i Bétera (València) (Gómez-Menor, 1957). També nova per Catalunya, la brunzidora de garriga T. garricola és comuna a França (Puissant 2006), però no apareix citada a l’Aragó per Aguin-Pombo et al. (2007), ni a la Península Ibèrica per Alonso-Zarazaga (2002). Probablement això és degut a què l’espècie va ser descrita al 1983, amb posterioritat a la majoria de prospeccions entomològiques. En canvi, la brunzidora cotonosa T. tomentosa, també nova a Catalunya, és rara a França, on manté petites poblacions fragmentades (Puissant, 2006) però se la coneix de la Serra de Guara (Aguin-Pombo et al., 2007) i sembla comuna en zones pseudo-estèpiques de la meitat sud peninsular. A Catalunya es va descobrir en secans del Segrià al 2013, gràcies a imatges pujades a ornitho.cat per I. Tejedor. Posteriorment els autors l’han localitzat a la Segarra (2017) i als Aiguamolls de l’Empordà (2018), i en tots tres casos sembla tractar-se de petites poblacions.
La descoberta de la cigala de l’olivera C. barbara per primer cop a Catalunya confirma el potencial dels vivers de planta ornamental com a vies d’entrada accidental d’organismes exòtics acompanyants. En concret, la hipòtesi més probable de l’aparició d’aquest cicàdid a dos vivers catalans
és el transport d’oliveres provinents del sud de la Península Ibèrica, on l’espècie és abundant als oliverars. Els exemplars poden ser transportats com a nimfes a les arrels dins el pa de terra i/o com adults a les capçades protegides per malles de transport. Aquesta forma de transport ha estat descrita per altres artròpodes introduïts, com l’aranya Macrothele calpeiana (Bellvert & Arnedo, 2016). Les observacions repetides anualment a un dels vivers plantegen una possible reproducció, malgrat que també podria tractar-se d’emergències de nimfes transportades amb diferents edats o en diferents anys. El fet que també se l’hagi trobat en un parc en ple eixample de Barcelona, però que mai se l’hagi trobat en medi natural allunyada de vivers, reforça la hipòtesi de la introducció. D’altra banda, es coneixen poblacions de C. barbara al País Valencià (Gómez-Menor, 1957) i Balears (exemplars del MCNB, obs. pers.), on podria ser un immigrant relativament recent (Pinto-Juma et al., 2009), originari del nord d’Àfrica, però no ha estat localitzada fins ara a les Terres de l’Ebre. S’ha descrit que, quan és molt abundant, la femella provoca danys a les tiges tendres de plantes cultivades, degut a les ferides causades per l’ovopositor durant la posta (González et al., 1998). Encara que no estan quantificades pèrdues de
68 Butlletí ICHN 85 (2), 2021
FLORA
ET FAUNA
P. PONS ET AL.
Figura 6. Distribució altitudinal a Catalunya de les espècies amb més de 20 registres recollits al portal ornitho.cat (2006-2020).
DIVERSITAT, DISTRIBUCIÓ I FENOLOGIA DE LES CIGALES (HEMIPTERA: CICADIDAE) A CATALUNYA
GEA, FLORA ET FAUNA
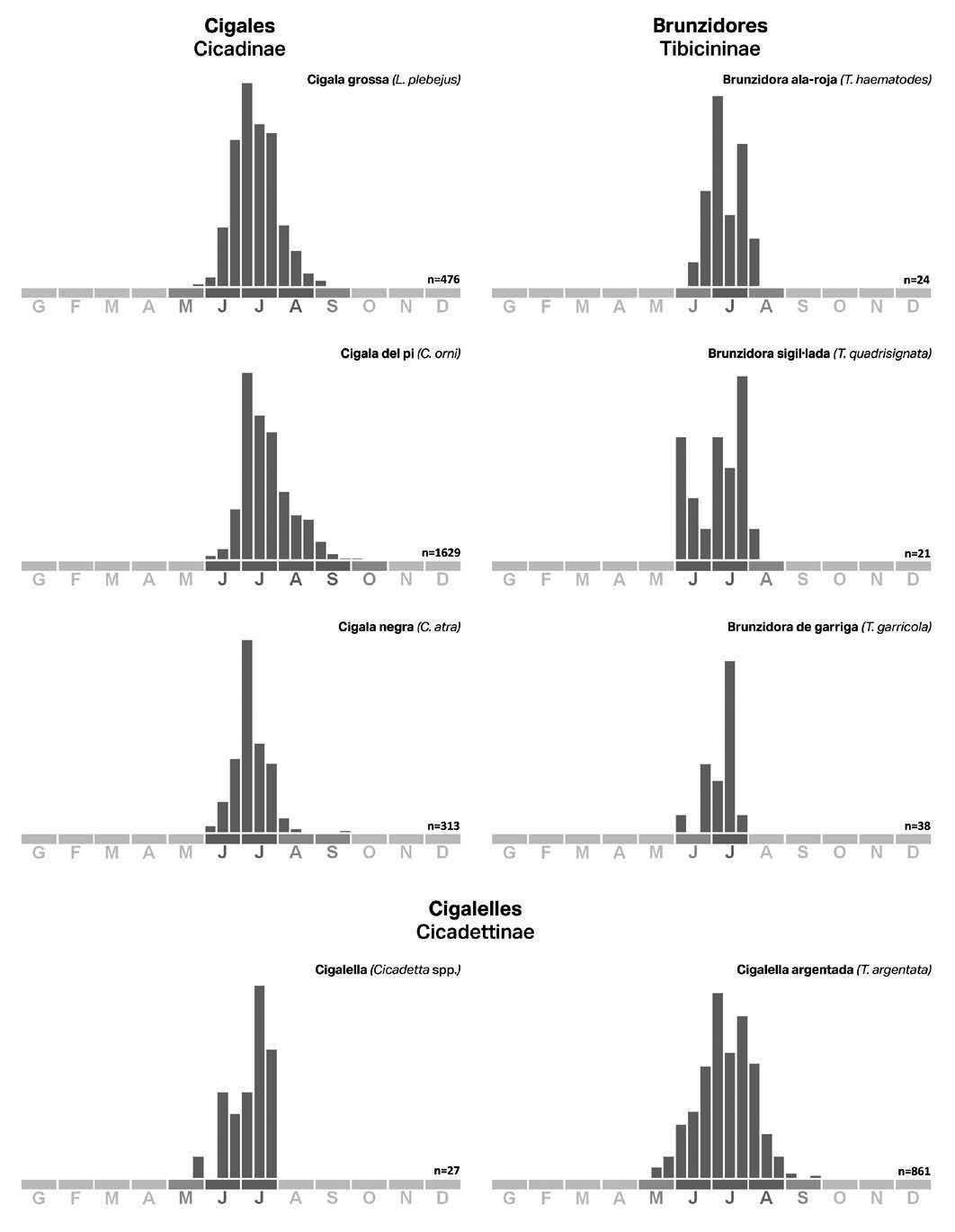
localitat) per Martorell i Peña (1879) com a Cicadetta flaveola. Malgrat això, cap dels set exemplars conservats al MCNB i etiquetats amb aquesta denominació és de localitat catalana (Pons i Carbonell, obs. pers.).
Es coneixen actualment 12 espècies de Cicadidae a Catalunya, nombre comparable a les 10 conegudes a Aragó (Aguin-Pombo et al., 2007); el nombre disminueix en excloure Cicadivetta tibialis, fruit de determinació errònia, com ja s’ha comentat. A la Península Ibérica es coneixen almenys 35 espècies, de les quals 13 es troben a Portugal, i un nombre major a Espanya (on no existeix un catàleg actualitzat), i la màxima diversitat aparentment a Andalusia (Alonso-Zarazaga, 2002; Sueur et al., 2004; Puissant & Sueur, 2010). Els departaments francesos de clima mediterrani, de menor extensió que Catalunya, tenen entre 4 (per exemple, la Lozère o Corse du Sud) i 11 espècies (l’Hérault), i el conjunt de França conté 20 espècies (Puissant, 2006). També són 11 les espècies d’Eslovènia, un país de mida similar al nostre però amb gran tradició d’estudi dels cicàdids. A Suïssa es coneixen 10 espècies (Hertach & Nagel, 2013) i a Àustria 8 (Trilar et al., 2020). El nombre baixa considerablement a major latitud i encara més si són territoris insulars, amb 5 espècies presents a Alemanya i tan sols una al Regne Unit (Hill et al., 2018). Cap a l’est, en canvi, es coneixen 15 espècies a Macedònia del Nord (Gogala et al., 2005) i Romania (Trilar & Gogala, 2008), 16-20 a Bulgària i 33 a Grècia (Trilar et al., 2020), països que han estat objecte de campanyes per l’equip del Museu d’Història Natural d’Eslovènia. Cal dir que Catalunya és l’únic d’aquests territoris on bona part de les cites de cicàdids provenen de ciència ciutadana, degudament validada amb el coneixement actual sobre determinació pels autors d’aquest treball, amb el suport d’especialistes en la taxonomia del grup.
producció d’olives degudes a la cigala de l’olivera (Martin & Ruiz, 2014), convindria monitoritzar la seva distribució, tant per potencials afectacions directes als oliverars com per si pot ser vectora del patogen de l’olivera Xylella fastidiosa (Serrano et al., 2016). D’altra banda no es descarta possible competència amb la cigala del pi, amb la que apareix en simpatria en àrees de la península (Seabra et al., 2006).
Hi ha dues espècies, citades històricament, que considerem determinacions errònies. Oligoglena tibialis va ser citada a la Garriga, Vallès Oriental, per Codina (1920), amb la denominació Pauropsalta tibialis. Aquest exemplar es conserva al MCNB, però no ha estat ha estat possible determinarne l’espècie. Segons Gogala et al. (2017), O. tibialis no és present a la península ibèrica, essent el límit occidental de la seva distribució l’extrem sud-est de França, concretament el Departament dels Alps Marítims. Aquests autors consideren que l’espècie es va identificar erròniament a partir d’exemplars pertanyents a alguna espècie del gènere Tympanistalna D’altra banda, segons Puissant & Sueur (2010) i Gogala et al. (2017) Oligoglena flaveola (Brull 1832) ha estat equivocadament citada al llarg dels segles XIX i XX a Espanya (per exemple, a Gómez-Menor, 1957), considerant aquests autors que ha estat confosa amb Euryphara virens i E. undulata, taxons que sí són presents a Espanya. Oligoglena flaveola havia estat citat anteriorment a Catalunya (sense detalls de la
Aquest estudi posa, per tant, de relleu el valor de la ciència ciutadana pel seguiment de la biodiversitat a escala regional (Callaghan et al., 2020). Entre les seves aplicacions, hi ha el tenir una millor imatge de la distribució, altitud i fenologia d’un grup animal, com s’exemplifica en aquest cas amb les cigales. El monitoratge d’espècies exòtiques invasores és un altre camp en què la ciència ciutadana pot tenir una gran rellevància, ja que permet obtenir moltes dades a temps real en àrees extenses. Cal remarcar la importància que els observadors reportin localitzacions precises, per poder associar-les a un hàbitat o altitud determinades. La participació de naturalistes amateurs, adequadament coordinada amb la tasca d’especialistes taxonòmics i investigadors en biodiversitat té un gran potencial per assolir un grau de coneixement encara més avançat, que permeti una millor gestió per la conservació. En el cas de les cigales, les dades de les plataformes de biodiversitat han contribuït a la major part de descobertes d’espècies anteriorment no detectades, a ampliar en gran manera l’extensió de la distribució coneguda, i han estat fonamentals pel coneixement de la fenologia a Catalunya.
Segons les dades del portal ornitho.cat (2006-2020) les comarques més diverses en cicàdids són l’Alt Empordà, amb 8 espècies, i la Segarra i l’Anoia, amb 7 espècies cadascuna. A l’altre extrem, no hi ha registres de cigales a l’Alta Ribagorça ni a la Cerdanya, i són escassos a les comarques més
69 Butlletí ICHN 85 (2), 2021
Figura 7. Nombre de registres per períodes de 10 dies, de les espècies amb més de 20 registres recollits al portal ornitho.cat (2006-2020).
GEA, FLORA ET FAUNA
pirinenques. Això es relaciona principalment amb la manca de prospecció, encara que el clima d’aquestes comarques també limiti la presència de les espècies més mediterrànies. Les cigales grossa, del pi i negre i la cigalella argentada són les més extensament repartides i presenten poblacions denses en hàbitats favorables. Les brunzidors tenen poblacions aparentment menors però que poden trobar-se any rere any als mateixos llocs, com demostren les observacions del portal ornitho.cat de les brunzidores sigiŀlada i de garriga. Les cigalelles de major mida i cant més audible (vespera i argentada) han estat lògicament localitzades a moltes més quadrícules que aquelles que requereixen anàlisi de sonogrames o de la genitàlia per la seva identificació (europea occidental i culprima). Aquest defecte de mostreig pot ser compensat amb l’ús d’equipament adequat de detecció i enregistrament sonor.
Aquest treball permet disposar també d’una aproximació a la distribució altitudinal i la fenologia les espècies més comunes a Catalunya. Els nostres resultats concorden amb el rang d’altituds i dates descrits a (Puissant, 2006) per França, tot i que amplien l’extensió del període d’activitat dels adults de les cigales del pi i negra, brunzidora sigiŀlada i cigalella argentada.
Amb la informació disponible, no és possible conèixer l’estat de conservació de les poblacions. Atenent a l’evolució dels hàbitats a les darreres dècades a Catalunya, hom pot pensar que les espècies generalistes i forestals, incloent-hi les cigales del pi, grossa i negra, així com part de les brunzidores i cigalelles (Puissant, 2006), poden tenir estatus més favorables. La cigala del pi, en concret, s’hauria beneficiat de l’expansió de les pinedes mediterrànies que, quan tenen obertura de capçada, poden contenir poblacions molt nombroses. Aquesta cigala, juntament amb la grossa i la negra, són també habituals en parcs i jardins urbans. Tot i així, l’important declivi dels insectes a Europa (Shortall et al., 2009; Hallmann et al., 2017) indica que part de les espècies de cigales poden haver disminuït a Catalunya des de mitjans del segle XX. Cal prospectar millor les especialistes d’ambients oberts, que presenten poblacions fragmentades i aparentment petites, específicament la brunzidora cotonosa i la cigalella vespera (endemisme ibèric). I cal augmentar l’esforç també per aquelles que es coneixen de molt poques localitats, com les cigalelles culprima (endemisme ibèric) i europea occidental i la brunzidora sigiŀlada.
En els propers anys és d’esperar que puguin aparèixer algunes espècies conegudes del sud de França, Aragó o País Valencià, encara no trobades a Catalunya (Taula 1). Dues d’aquestes són les cigalelles pirinenca Cicadetta cerdaniensis i la costanera C. brevipennis litoralis, localitzades a la Catalunya Nord a escassos quilòmetres del Principat (Hertach et al., 2016). Altres espècies noves poden provenir de revisions taxonòmiques dels Cicadettinae, ja realitzades pel gènere Cicadetta, però aparentment pendents per Euryphara Horváth, 1912 i Tettigettalna Puissant, 2010. En ambdós supòsits caldran noves prospeccions al territori i el suport de tècniques bioacústiques i moleculars per la determinació de les espècies. D’altra banda, manca coneixement sobre els hàbitats
seleccionats per les cigales a Catalunya, ja que una mateixa espècie pot presentar preferències distintes en diferents territoris (Puissant, 2006). Cal completar, així mateix, la informació fenològica per la majoria d’espècies, aspecte fonamental per millorar-ne les prospeccions i per gestionar els hàbitats. Aplicar aquest coneixement per aquelles amb interès de conservació, permetria evitar estassades o cremes dels estrats herbaci i llenyós durant el període en què els ous són a la vegetació. Altres aspectes de l’ecologia del grup, especialment de la fase juvenil endogea, encara són força desconeguts i mereixedors d’estudi, ja que pertorbacions que afectin el sistema radicular poden perjudicar el relleu de les poblacions de cigales (Pons, 2015). Finalment, suggerim començar a monitoritzar les poblacions dels endemismes ibèrics i altres espècies de distribució reduïda, com a informació fonamental per valorar possibles mesures de conservació.
Agraïments
L’estudi ha utilitzat les dades del portal ornitho.cat, cedides per l’Institut Català d’Ornitologia. Volem agrair la contribució dels observadors i dels gestors d’ornitho.cat, i en especial de Marc Anton, Toni Arrizabalaga, Ignasi Tejedor i Oriol Baltà, per la seva coŀlaboració. Les estudiants de la Universitat de Girona (UdG), Anna Gràcia i Aina Perich han ajudat amb les prospeccions realitzades i Aitor Domingo amb la creació de la pàgina web de cicada.cat. Glòria Fontova va vehicular la coŀlaboració amb el Termcat i Berta Caballero ens va facilitar la consulta de la coŀlecció del Museu de Ciències Naturals de Barcelona. La biblioteca de la UdG i Montserrat Grabolosa ens van ajudar en la cerca bibliogràfica. Agraïm als especialistes Stéphane Puissant, Michel Boulard i Matija Gogala la informació taxonòmica proporcionada i el seu suport.
Bibliografia
Aguin-Pombo, D., Freitas, C., Álvarez, P. A. & Bourgoin, T. 2007. Catálogo de los hemípteros Cicadomorpha y Fulgoromorpha de Aragón. Catalogus de la entomofauna aragonesa, 34: 3-22.
Alonso-Zarazaga, M. A. 2002. Fauna Ibérica: Hemípteros. Disponible a: http://www.fauna-iberica.mncn.csic.es/faunaib/ arthropoda/insecta/hemiptera [Data de consulta: 16 febrer 2021].
Bellvert, A. & Arnedo, M. A. 2016. Threatened or threatening? Evidence for independent introductions of Macrothele calpeiana (Araneae: Hexathelidae) and first observation of reproduction outside its natural distribution range. Arachnology, 17: 137-141.
Boulard, M. & Mondon, B.. 1995. Vies et mémoires de cigales. Editions de l’Equinoxe. Barbentane, France. 160 p.
Callaghan, C. T., Poore, A. G. B., Mesaglio, T., Moles, A. T., Nakagawa, S., Roberts, C., Rowley, J. J. L., Vergés, A., Wilshire, J. H. & Cornwell, W. K. 2020. Three frontiers for the future of biodiversity research using citizen science data. Bioscience, 71: 55-63.
Carbonell, R. 2011. Invertebrats del massís del Mont. Annals de l’Institut d’Estudis Empordanesos, 42: 48-60.
Carbonell, R. & Matheu, E. 2010. Guia sonora dels insectes de Catalunya: grills, saltamartins i cigales. Alosa i Museu de Granollers. Barcelona. CD i llibret.
70 Butlletí ICHN 85 (2), 2021
P. PONS ET AL.
DIVERSITAT, DISTRIBUCIÓ I FENOLOGIA DE LES CIGALES (HEMIPTERA: CICADIDAE) A CATALUNYA
Codina, A. 1920. Lista lnédita de Homópteros (Hemípteros) de Cataluña. Boletín de la Sociedad Entomológica de España, 3: 100-104.
Cuní i Martorell, M. 1883. Resultado de una exploracion entomológica y botánica por el termino de la Garriga (Cataluña). Anales de Historia Natural, 12: 83-101.
Cuní i Martorell, M. 1885. Excursion entomológica á varias localidades de la Provincia de Gerona (Cataluña). Anales de Historia Natural, 14: 51-73.
Cuní i Martorell, M. 1888. Insectos observados en los Alrededores de Barcelona. Anales de Historia Natural, 17: 133-191.
Farina, A., Pieretti, N. & Piccioli, L. 2011. The soundscape methodology for long-term bird monitoring: A Mediterranean Europe case-study. Ecological Informatics, 6: 354-363.
Gogala, M. 2019. Songs of European singing cicadas. Disponible a: http://www.cicadasong.eu [Data de consulta: 20 gener 2021].
Gogala, M., Puissant, S., & Trilar, T. 2017. Revision and resurrection of the genus name Mezammira Fieber, 1876 (Hemiptera: Cicadidae) with special focus on its species from Greece and the description of two new species. Acta Entomologica Slovenica, 25: 5-64.
Gogala, M., Trilar, T. & Krpač, V. T. 2005. Fauna of singing cicadas (Auchenorrhyncha: Cicadoidea) of Macedonia - a bioacoustic survey. Acta Entomologica Slovenica, 13: 103-126.
Gómez-Menor, J. 1957. Monografía de Cicádidos (Homoptera) de España. Memoria de la Real Academia de Ciencias Físicas, Exactas y Naturales de Madrid. Serie de Ciencias Naturales, 19: 1-88.
Goula, M., Ribes, J. & Serra, A. 2010. Llista dels Heteròpters de Catalunya (Insecta, Hemiptera, Heteroptera).Versió 1, juny 2010. Centre de Recursos de Biodiversitat Animal. Universitat de Barcelona, Barcelona. 38 p.
GR-Biologia Animal. 2021. Cicada.cat: Els Cicadidae a Catalunya. Disponible a: http://cicada.cat [Data de consulta: 20 febrer 2021].
Hallmann, C. A., Sorg, M., Jongejans, E. Siepel, H., Hofland, N., Schwan, H., Stenmans, W., Müller, A., Sumser, H., Hörren, T., Goulson, D. & De Kroon, H. 2017. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLOS ONE, 12: e0185809.
Hertach, T. 2020. Look closely and listen carefully: unexpected cicada diversity in northern Sardinia, with the description of a new species (Cicadidae: Tibicina). Zoological Journal of the Linnean Society, 20: 1-23.
Hertach, T. & Nagel, P. 2013. Cicadas in Switzerland: a scientific overview of the historic and current knowledge of a popular taxon (Hemiptera: Cicadidae). Revue Suisse de Zoologie, 120: 229-269.
Hertach, T., Puissant, S., Gogala, M., Trilar, T., Hagmann, R., Baur, H., Kunz, G., Wade, E. J., Loader, S. P., Simon, C. & Nagel, P. 2016. Complex within a complex: integrative taxonomy reveals hidden diversity in Cicadetta brevipennis (Hemiptera: Cicadidae) and unexpected relationships with a song divergent relative. PLOS ONE, 11: e0165562.
Hill, A. P., Prince, P., Piña-Covarrubias, E., Doncaster, C. P., Snaddon, J. L. & Rogers, A. 2018. AudioMoth: Evaluation of a smart open acoustic device for monitoring biodiversity and the environment. Methods in Ecology and Evolution, 9: 1199-1211.
Koenig, W. D. & Liebhold, A. M. 2005. Effects of periodical cicada emergences on abundance and synchrony of avian populations. Ecology, 86: 1873-1882.
Lin, N., Berton, P., Moraes, C., Rogers, D. & Tufenkji, N. 2018. Nanodarts, nanoblades, and nanospikes: mechano-bactericidal
nanostructures and where to find them. Advances in Colloid and Interface Science, 252: 55-68.
Marshall, D. C., Moulds, M., Hill, K. B. R., Price, B., W., Wade, E. J., Owen, C. L., Goemans, G., Marathe, K., Sarkar, V., Cooley, J. R., Sanborn, A. F., Kunte, K., Villet, M. H. & Simon, C. 2018. A molecular phylogeny of the cicadas (Hemiptera: Cicadidae) with a review of tribe and subfamily classification. Zootaxa, 4424: 1-64.
Martin, A. & Ruiz, M. J. 2014. Guía de gestión integrada de plagas: Olivar. Ministerio de Agricultura, Alimentación y Medio Ambiente. Madrid. 178 p.
Martorell i Peña, M. 1879. Catálogos sinonímicos de los Insectos encontrados en Cataluña: Hemipteros. Barcelona. 202 p
Mehdipour, M., Zamanian, H., Farazmand, H. & Hosseini-Gharalari, A. 2016. Disruption of reproductive behavior of grapevine cicada, Cicadatra alhageos, by acoustic signals playback. Entomologia Experimentalis et Applicata 158:210-216.
Navàs, L. 1930. Excursió entomològica a la vall de Noguera de Cardós (Lleida). Butlletí de la Institució Catalana d’Història Natural, 30: 156-169.
Pinto-Juma, G. A., Quartau, J. A. & Bruford, M. W. 2009. Mitochondrial DNA variation and the evolutionary history of the Mediterranean species of Cicada L. (Hemiptera, Cicadoidea). Zoological Journal of the Linnean Society, 155: 266-288.
Pons, P. 2015. Delayed effects of fire and logging on cicada nymph abundance. Journal of Insect Conservation, 19: 601-606.
Puissant, S. 2005. Taxonomy, distribution and first eco-ethological data of Melampsalta varipes (Waltl, 1837), an unrecognized cicada (Hemiptera, Cicadidae). Insect Systematics & Evolution, 36: 301-315.
Puissant, S. 2006. Contribution a la connaissance des cigales de France: Géonémie et écologie des populations (Hemiptera, Cicadidae) Ascete, Bédeilhac et Aynat, France.
Puissant, S. & Sueur, F. 2010. A hotspot for Mediterranean cicadas (Insecta: Hemiptera: Cicadidae): new genera, species and songs from southern Spain. Sistematics and Biodiversity, 8: 555–574.
Seabra, S. G., Pinto-Juma, G. & Quartau, J. A. 2006. Calling songs of sympatric and allopatric populations of Cicada barbara and C. orni (Hemiptera: Cicadidae) on the Iberian Peninsula. Zoological Studies, 103: 843-852.
Serrano, A., González, M. I., Sánchez, A. M. & Durán, J. M. 2016. Vectores potenciales de Xylella fastidiosa en el olivar de la provincia de Sevilla. Phytoma, 277: 54-59.
Shortall, C. R., Moore, A., Smith, E., Hall, M. J., Woiwod, I. P. & Harington, R. 2009. Long-term changes in the abundance of flying insects. Insect Conservation and Diversity, 2: 251-260.
Simoes, P. C., Sanborn, A. & Quartau, J. A. 2013. Two new species of Cicadatra (Hemiptera: Cicadoidea) from Greece. Entomological Science, 16: 83-90.
Simon, C., Gordon, E. R. L., Moulds, M. S., Cole, J. A., Haji, D., Lemmon, A. R., Lemmon, E. M., Kortyna, M., Nazario, K., Wade, E. J., Meister, R. C., Goemans, G., Chiswell, S. M., Pessacq, P., Veloso, C., Mccutcheon, J. P. & Łukasik, P. 2019. Off-target capture data, endosymbiont genes and morphology reveal a relict lineage that is sister to all other singing cicadas. Biological Journal of the Linnean Society, 128: 865-886.
Srivastava, S. K., Babu, N. & Pandey, H. 2009. Traditional insect bioprospecting - As human food and medicine. Indian Journal of Traditional Knowledge, 8: 485-494.
Sueur, J., Puissant, S., Simoes, P., Seabra, S., Boulard, M. & Quartau, J. A. 2004. Cicadas from Portugal: revised list of species with eco-ethological data (Hemiptera: Cicadidae). Insect Systematics & Evolution, 35: 177-187.
71 Butlletí ICHN 85 (2), 2021 GEA, FLORA ET FAUNA
GEA, FLORA ET FAUNA
Termcat. 2021. Neoloteca. Disponible a:https://www.termcat.cat/ ca/neoloteca. [Data de consulta: 18 febrer 2021].
Trilar, T., Gjonov, I. & Gogala, M. 2020. Checklist and provisional atlas of singing cicadas (Hemiptera: Cicadidae) of Bulgaria, based on bioacoustics. Biodiversity Data Journal, 8: e54424.
Trilar, T. & Gogala, M. 2008. New data on singing cicadas (Hemiptera: Cicadidae) of Romania. Entomologica Romanica, 13: 29-33.
Williams, K. S. & Simon, C. 1995. The Ecology, Behavior, and Evolution of Periodical Cicadas. Annual Review of Entomology, 40: 269-295.
72 Butlletí ICHN 85 (2), 2021
ET AL.
P. PONS
Butlletí de la Institució Catalana d’Història Natural, 85 (2): 73-75. 2021
NOTA BREU
Alloeorhynchus (Alloeorhynchus) flavipes (Fieber, 1836), espècie reconfirmada per a la fauna ibèrica (Hemiptera: Heteroptera; Nabidae)
Alloeorhynchus (Alloeorhynchus) flavipes (Fieber, 1836), reconfirmed species to the Iberian fauna (Hemiptera: Heteroptera: Nabidae)
Marta Goula*, Rafael Carbonell**, Elena López Gallego*** & Jan Tomàs****
* Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals & IRBio. Facultat de Biologia. Universitat de Barcelona. Av. Diagonal, 643, 08028 Barcelona. ORCID 0000-0001-6308-342X.
** Can Grau, 17850 Beuda (Girona).
*** Instituto Murciano de Investigación y Desarrollo Agrario y Alimentario (IMIDA). Equipo de Control Biológico y Servicios Ecosistémicos. C/ Mayor, s/n, 30150 La Alberca (Murcia). ORCID 0000-0002-1747-7468.
**** C/ Escultor Armengol, 16, 2n 2a, 08222 Terrassa (Barcelona).
Autora per a la correspondència: Marta Goula. A/e: mgoula@ub.edu
Rebut: 07.06.2021. Acceptat: 10.06.2021. Publicat: 30.06.2021
El gènere Alloeorhynchus Fieber, 1860 inclou espècies amb formes braquípteres i macròpteres, relativament piloses. Certes regions del cos, com ara el cap, el pronot i l’abdomen, són lluents, mentre que l’escudet i els hemèlitres són mats o gairebé. Els profèmurs i les protíbies engrossits els enquadren dins de la subfamília Prostemmatinae, que en el context paleàrtic inclou els gèneres Prostemma Laporte de Castelnau, 1832 i Alloeorhynchus (ambdós de la tribu Prostemmatini, amb el connexiu no delimitat ventralment), i Phorticus Stål, 1860 i Rhamphocoris Kirkaldy, 1901 (ambdós de la tribu Phorticini, amb el connexiu delimitat ventralment) (Kerzhner, 1996). Prostemma acostuma a fer més de 5 mm de llarg, té els profèmurs espinosos i amb una prominència al mig de la cara ventral, i la tercera esternita abdominal més curta que el doble de la llargada de la quarta. En canvi, Alloeorhynchus fa com a màxim 5 mm de llarg, té espines als profèmurs i als mesofèmurs i ambdós són prominents al mig de la cara ventral, i la tercera esternita abdominal més llarga que el doble de la llargada de la quarta (Péricart, 1987).
En l’àmbit paleàrtic es coneixen nou espècies d’Alloeorhynchus, de les quals set pertanyen al subgènere nominal (A. aegyptius El-Sebaey, 2002, A. flavipes (Fieber, 1836), A. notatus Distant, 1919, A. putoni Kirkaldy, 1901, A. reinhardi Kerzhner & Günther, 1999, A. sinicus Ren & Bu, 1998 i A. vinulus Stål, 1864), una al subgènere Psilistus Stål, 1873 (A. corallinus (Stål, 1873)) i la restant, A. wau Linnavuori, 1974, es considera incertae sedis (Kment & Carapezza, 2017). D’aquestes, la majoria són de distribució principalment asiàtica, tret d’A. aegyptius, del nord d’Àfrica (Egipte), A. putoni, del Mediterrani occidental (les citacions d’Hongria, Grècia i Síria són errònies segons Kerzhner (1996)) i A flavipes, que és present en nombrosos països d’Europa i en alguns de l’Àsia occidental (Aukema, 2018).
A la península Ibèrica, A. putoni s’ha assenyalat de Catalunya (Barcelona), el País Valencià (Castelló) i Andalusia (Cadis), en localitats costaneres o properes al litoral (Péricart, 1987), i també de Portugal (Ganilhos, a 1 km d'Alcobaça, sobre noguera) (Ribeiro & Pinto de Abreu, 1997). En la coŀlecció de Jordi Ribes, dipositada al Centre de Recursos de Biodiversitat Animal (CRBA) de la Facultat de Biologia de la Universitat de Barcelona, hi ha una femella macròptera d’aquesta espècie capturada a Pina de Ebro (Saragossa) (vegeu Material estudiat), dada no publicada que amplia la seva àrea de distribució coneguda cap a l’interior peninsular. Per a A. flavipes, existeix una citació de Badajoz (Extremadura), Uhagon leg. (Chicote, 1880), que Péricart (1987) recull amb un interrogant, amb el comentari «citation pouvant se rapporter à flavipes ou à putoni». S’ha intentat localitzar l’exemplar en qüestió al Museo Nacional de Ciencias Naturales de Madrid (vegeu Agraïments), lamentablement sense èxit, i, per tant, aquesta citació roman pendent de verificació.
La clau publicada per Péricart (1987) permet diferenciar clarament A. putoni d’A. flavipes sense necessitat d’haver d’estudiar la genitàlia masculina. En efecte, A. putoni, més petit (3,3-4,3 mm), és d’aspecte més esvelt (Fig. 1a, b), té el cap més estret (< 0,65 mm) i els ulls més petits, i els hemèlitres de les seves formes braquípteres són més aviat aguts a l’àpex i generalment depassen la meitat de la segona tergita abdominal. Alloeorhynchus flavipes és d’aspecte més robust (Fig. 1c, d) i acostuma a ser més gros (3,6-5,2 mm), té el cap més ample (> 0,68 mm) i els ulls més grossos, i els hemèlitres de les seves formes braquípteres són més o menys arrodonits a l’àpex i no s’estenen més enllà de la meitat de la segona tergita abdominal. La majoria d’exemplars d’A. putoni, a més, presenten una taca central grogosa més o menys extensa entre la tercera i la sisena tergita (Fig. 1b), mentre que en A.
73 Butlletí ICHN 85 (2), 2021
BREU
NOTA
edition)
ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print
DOI: 10.2436/20.1502.01.75

Figura 1. Habitus de les dues espècies ibèriques d’Alloeorhynhus. a) ♀ macròptera d’A. putoni (Pina de Ebro, Saragossa; CRBA-68921), b) ♂ braquípter d’A. putoni (Gallifa, Barcelona; CRBA-68915), c) ♀ braquíptera d’A. flavipes (Beuda, Girona) i d) ♂ braquípter d’A. flavipes (Trenčín,
CRBA-68914). Escala = 1 mm.
Taula 1. Valors biomètrics (en mm) d’interès taxonòmic en les espècies ibèriques d’Alloeorhynchus. br.: braquípter/a, macr.: macròptera.
flavipes l’abdomen sempre és completament negre (Carayon, 1949) (Fig. 1 c-d). L’estudi de material dipositat al CRBA ha permès confirmar aquests caràcters; els valors biomètrics principals mesurats en tots els exemplars examinats d’ambdues espècies es poden consultar a la Taula 1.
Material estudiat
A. putoni
ESPANYA: 1 ♀ braquíptera (br.), San Roque, Campo de Gibraltar, Cadis, 00/07/1971 (CRBA-43642, coŀlecció Vives & Vives); 1 ♂ br., Gallifa, Vallès Occidental, Barcelona,
Amb aquesta nota, queda ben confirmada la presència d’A. flavipes a territori ibèric, i la posició septentrional de la localitat on s’ha trobat (a la comarca de la Garrotxa) és coherent amb la distribució general de l’espècie, molt present al centre d’Europa. Ara bé, segons Péricart (1987), aquest nàbid habita llocs assolellats, boscosos o herbacis, concretament (almenys a França) on abundin els brucs o les molses, o a vores de pinedes. Durant el dia es queda més aviat a la base de les plantes, prop de les seves arrels o sota pedres. Per tant, detalls de la seva biologia que no indiquen exigències estrictes o hàbitats molt particulars, als quals la troballa de l’exemplar origen d’aquesta nota no permet afegir res, ja que es va col·lectar mort en una bassa d’una finca particular (Can Grau, Beuda). Aquesta poca especialització aparent fa pensar que pugui tractar-se d’una espècie més estesa i freqüent a la Península del que indiquen les citacions fins al moment present, i no es pot descartar la veracitat de la citació de Badajoz de Chicote (1880). És cert, però, que la distinció d’A. flavipes i A. putoni requereix una observació atenta, i potser la revisió de coŀleccions ens descobriria exemplars de la primera espècie que incurosament s’han atribuït a la segona, segurament per la inèrcia de considerar A. flavipes estesa al nord dels Pirineus (Carayon, 1949).
74 Butlletí ICHN 85 (2), 2021 NOTA BREU M. GOULA ET AL.
Eslovàquia;
Espècie Localitat Exemplar Sexe Amplada cap Amplada ull Amplada màxima Longitud total putoni Gallifa, Barcelona CRBA-68915 ♂ br. 0,60 0,16 1,45 3,40 Tírig, Castelló CRBA-68920 ♂ br. 0,60 0,17 1,15 3,05 Pina de Ebro, Saragossa CRBA-68921 ♀ macr. 0,60 0,16 1,50 4,00 San Roque, Cadis CRBA-43642 ♀ br. 0,60 0,17 1,50 3,75 flavipes Turquia CRBA-68913 ♂ br. 0,80 0,25 1,75 4,50 Eslovàquia CRBA-68914 ♂ br. 0,80 0,25 1,80 4,40 Catalunya Can Grau, Beuda ♀ br. 0,80 0,24 1,95 4,65 Alemanya CRBA-68912 ♀ br. 0,80 0,26 1,90 4,65
14/04/1963 (CRBA-68915, col·lecció J. Ribes) (Fig. 1b); 1 ♂ br., Tírig, Alt Maestrat, Castelló, 01/04/1972 (CRBA68920, coŀlecció J. Ribes); 1 ♀ macròptera, Pina de Ebro, Monegres, Saragossa, 20/05/1991 (CRBA-68921, coŀlecció J. Ribes) (Fig. 1a).
A. flavipes
ALEMANYA: 1 ♀ br., Treuchtlingen, Weißenburg-Gunzenhausen, Baviera (Freistaat Bayern), 25/02/1957 (CRBA68912, coŀlecció J. Ribes). TURQUIA: 1 ♂ br., Çekirge, Bursa, 04/06/1961 (CRBA-68913, col·lecció J. Ribes). ESLOVÀQUIA: 1 ♂ br., Trenčín, s.d. (CRBA-68914, coŀlecció J. Ribes) (Fig. 1d). ESPANYA: 1 ♀ br., Can Grau, Beuda, Girona, 31TDG7477, 395 msnm, 04/04/2021, ofegat en una bassa, R. Carbonell leg., coŀlecció M. Goula (Fig. 1c).
Agraïments
A Mercedes París (Museo Nacional de Ciencias Naturales, Madrid) i a Marcos Roca-Cusachs (Universitat de Barcelona), per la cerca conjunta a la coŀlecció del Museo de l’exemplar d’A. flavipes de Badajoz, Uhagon leg., durant una estada del segon a aquella institució l’abril de 2021, tot i que la cerca fos infructuosa. A Helena Basas (CRBA) per la seva amabilitat i eficàcia en relació amb el préstec dels exemplars estudiats. Finalment, i en especial, també agraïm a en Marcos
Roca-Cusachs i a Juli Pujade-Villar (Universitat de Barcelona) la revisió crítica del manuscrit.
Bibliografia
Aukema, B. 2018. Catalogue of Palaearctic Heteroptera. Disponible en: https://catpalhet.linnaeus.naturalis.nl/linnaeus_ng/app/views/ introduction/topic.php?id=9&epi=1 [Data de consulta: 5 juny 2021].
Carayon, J. 1949. Caractères distinctifs, répartitions géographiques et habitats des espèces paléarctiques du genre Alloeorhynchus Fieb. (Hem. Nabidae). Bulletin de la Société entomologique de France, 54 (9): 136-142.
Chicote, C. 1880. Adiciones a la enumeración de los Hemípteros observados en España y Portugal. Anales de la Sociedad Española de Historia Natural, 9: 185-203.
Kerzhner, I. M. 1996. Family Nabidae A. Costa, 1853 – damsel bugs P. 84-107. In: Aukema, B. & Rieger, Chr. (ed.). Catalogue of the Heteroptera of the Palaearctic Region. Volume 2. Cimicomorpha
I. The Netherlands Entomological Society. Amsterdam. Països Baixos. 361 p.
Kment, P. & Carapezza, A. 2017. List of true bug taxa described by Rauno E. Linnavuori (Hemiptera: Heteroptera). Entomologica Americana, 122 (4): 528-621.
Péricart, J. 1987. Hémiptères Nabidae d’Europe occidentale et du Maghreb. Fédération Française des Sociétés de Sciences Naturelles. París. 185 p. (Faune de France, 71)
Ribeiro, F. N. & Pinto de Abreu, C. 1997. Studies on beneficial insects from the superficial stratum of a walnut orchard soil. ISHS Acta Horticulturae, 442: 407-418.
75 Butlletí ICHN 85 (2), 2021 NOTA BREU ALLOEORHYNCHUS (ALLOEORHYNCHUS) FLAVIPES, ESPÈCIE RECONFIRMADA PER A LA FAUNA IBÈRICA
Un nou gènere i espècie de Molopina hipogeu del Pirineu Català (Alt Urgell, Lleida) (Coleoptera: Carabidae: Pterostichini)
Eduard Vives* & Jordi Comas*
* Museu de Ciències Naturals de Barcelona. Laboratori de Natura. Col·lecció d’artròpodes. Passeig Picasso, s/n. 08003 Barcelona. A/e: eduard_vives@ hotmail.com; jcomas2@gmail.com
Autor per a la correspondència: Eduard Vives: A/e: eduard_vives@hotmail.com
Rebut: 13.05.2021; Acceptat: 14.06.2021; Publicat:30.06.2021
Resum
Es descriu un nou gènere i una i nova espècie hipogea de la subtribu Molopina (Coleoptera: Carabidae: Pterostichini). Speleopidius joanivivesi n. gen. & n. sp., procedent de l’Avenc de la Cabana d’en Garraba, Pont de Bar (L’Alt Urgell, Lleida, Espanya). Es compara la relació del nou gènere i nova espècie amb d’altres Molopina hipogeus de la mediterrània occidental.
Paraules clau: Coleoptera, Carabidae, Pterostichini, Molopina, Speleopidius n. gen. joanivivesi n. sp., L’Alt Urgell, Lleida, Catalunya, Espanya.
Abstract
A new genus and new species of hypogeum Molopina from Catalan Pyrenean area (L’Alt Urgell, Lleida) (Coleoptera: Carabidae: Pterostichini)
It is described a new genus and a new hypogean species of the subtribe Molopina, tribe Pterostichini (Coleoptera, Carabidae), namely Speleopidius joanivivesi n. gen. & n. sp., from the Avenc de la Cabana d’en Garraba, Pont de Bar (L’Alt Urgell), Lleida, Spain. The relationships of the new taxa with other hypogean Molopina of the Western Mediterranean are discussed.
Key words: Coleoptera, Carabidae, Pterostichini, Molopina, Speleopidius n. gen. joanivivesi n. sp., L’Alt Urgell, Lleida, Catalonia, Spain.
urn:lsid:zoobank.org:pub:043B7D4E-F903-4509-9414-720F94DBDE99
Introducció
En el llarg procés d’elaboració i publicació del «Catàleg dels Coleòpters Cavernícoles de Catalunya» (Pallisé, 2020) el nostre amic i company en bioespeleologia, Joan Pallisé i el seu equip de coŀlaboradors de recerques han visitat un gran nombre de cavitats subterrànies de zones catalanes poc explorades, per tal de completar la distribució d’algunes espècies de coleòpters cavernícoles ja coneguts, però també han visitat d’altres cavitats conegudes per confirmar la presència d’algunes espècies. Entre elles visitaren cavitats de L’Alt Urgell i La Cerdanya, per tal de recollir noves mostres o citacions.
El mes d’agost de 2018 s’exploraren les cavitats de la zona de Toloriu i de Pont de Bar, entre aquestes l’Avenc de la Cabana d’en Garraba, del qual ja es coneixia la presència del Leioidae cavernícola Troglocharinus mengeli (Jeannel, 1910) i del Carabidae Trechini hipogeu Geotrechus incantatus Faille, Bourdeau, Bellés & Fresneda, 2015. Entre el material recollit aparegueren dues larves del coleòpter Cantharidae Armidia unicolor (Dufour, 1851) que realitza una fase del seu desenvolupament a les cavitats subterrànies (Español
& Belles, 1980), i els restes d’uns èlitres d’una espècie de caràbid desconegut. Aquest descobriment propicià que es repetissin les visites a aquest avenc de L’Alt Urgell i a d’altres cavitats properes, per tal d’intentar aconseguir algun exemplar sencer. Durant tres anys de repetides exploracions i paranys a la mencionada cavitat i del seu entorn, ha estat possible capturar dos exemplars de femelles d’aquest caràbid hipogeu que ha resultat tractar-se d’un nou gènere i nova espècie de Pterostichini, Molopina que tot seguit descrivim.
Materials i mètode
Els exemplars i les restes es conserven en sec, muntats sobre cartolina entomològica. La genitàlia femenina extreta per dissecció, ha estat tractada en immersió amb lactofenol durant vint-i-quatre hores, procedint posteriorment a la tinció amb negre de clorazol, i muntada sobre plàstic transparent dins de polivinil-lactofenol. Per l’estudi dels materials s’ha utilitzat una lupa estereoscòpica WILD M5A, i les fotografies s’han fet amb una càmera Canon EOS 30 amb objectiu
77 Butlletí ICHN 85 (2), 2021 GEA, FLORA ET FAUNA GEA, FLORA ET FAUNA
(2): 77-82. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition) DOI: 10.2436/20.1502.01.76
Butlletí de la Institució Catalana d’Història Natural, 85
Canon, EF 100m-F/2.8 Macro IS. Les fotografies per capes s’han processat amb l’aplicació Combine-ZP i algunes estan compensades amb el programa Adobe Photoshop-CS6 extended.
Els acrònims utilitzats són els següents: MCNB (coŀlecció Museu de Ciències Naturals de Barcelona), CEV (coŀlecció Eduard Vives, Terrassa), CJC (coŀlecció Jordi Comas. Barcelona) i MSS (Medi subterrani superficial).
Resultats
Speleopidius n. gen. (Fig. 1)
Espècie tipus: Speleopidius joanivivesi n. sp. urn:lsid:zoobank.org:act:DA4A7171-A287-4AEC-B687-B8FBAD4727BB

Diagnosi
Aquest nou gènere forma part del grup de coleòpters Carabidae de la tribu Pterostichini, subtribu Molopina (Serrano, 2013 i 2020) que viuen al medi hipogeu. Per la seva morfologia s’assembla al gènere Zariquieya Jeannel, 1924, del qual se separa per la forma dels èlitres ja que presenten la zona discal lleugerament convexa, al contrari de Zariquieya que els té fortament deprimits formant una concavitat. La zona apical és amplament arrodonida en Speleopidus n. gen., mentre que és truncat a Zariquieya i amb un gros mugró preapical que falta a Speleopidius n. gen A diferència dels gèneres de Molopina hipogeus de la fauna ibèrica Oscadytes Lagar, 1975 i Zariquieya que no tenen estriola juxta-escutelar, el nou gènere posseeix una petita estriola juxtaescutelar com el gènere Molopidius Jeannel, 1942.
Com la majoria de Molopina hipogeus de la mediterrània occidental, Speleopidius n. gen. és cec, i amb una cicatriu de la resta d’ommatidis.
Descripció
Speleopidius n. gen. té el cap arrodonit, lleugerament convex a la part superior. Els ulls reduïts a una petita taca groga que assenyala la resta dels ommatidis. Els costats del cap són molt convexos, però sense arribar a formar la gran protuberància que té Zariquieya. Les antenes són llargues i fines, amb els artells pubescents a partir del tercer. El pronot és aplanat i amb els costats sinuosos, amples a la part anterior, estrenyent-se cap a la part posterior que és recte formant un petit angle recte a cada costat. Els angles anteriors són petits i arrodonits. El prostern (Fig. 3b) és més ample que llarg i amb el procés prosternal arrodonit a la seva part posterior. El mesostern (Fig. 3c) és llarg i estret, bífid a la part posterior del seu procés mesosternal. El metastern és ample i curt, aplanat a la part central. Èlitres llargs i estrets, amb els costats poc paraŀlels, àmpliament arrodonits a la part apical. Les estries elitrals són llises, interestries planes, sense restes de cap porus pilífer com passa en d’altres gèneres de Molopina hipogeus de la mediterrània occidental.
Tota la zona central del disc elitral és lleugerament convex. Vora lateral dels èlitres molt marcada i amb llargues setes umbilicades. Part inferior de l’abdomen amb el tercer esternit visible més llarg que la resta d’esternits. Potes moderadament allargades, amb els profèmurs i mesofèmurs engruixits a la seva part apical. Tarses no dilatats, llargs i recoberts de pilositat a la seva cara superior. Ungles simples i divergents.
Etimologia
Nom compost de: Speleo- (del llatí spelunca = cova) i -pidius (del gènere Molo-pidius Jeannel). Gènere masculí.
Speleopidius joanivivesi n. sp. (Fig. 2a)
urn:lsid:zoobank.org:act:F609F432-38B3-4913-8FE3-8FBB5B8D2F98
Diagnosi
Cap molt arrodonit globulós, proveït de dues llargues setes supraorbitals. Restes dels ulls en forma de cicatriu enfonsada en una fosseta arrugada. Coll molt ample i convex. Pronot subquadrat, aplanat a la cara superior, amb els angles anteriors arrodonits i poc sortints. Els angles posteriors en forma d’angle recte. Escudet triangular i amb una estriola a cada costat. Un profund solc longitudinal a la meitat del disc que no arriba als marges. Fossetes basals del pronot aplanades i rugoses. Èlitres llargs i estrets, amb una vora ampla proveïda de llargues setes umbilicades que es distribueixen amb una topografia molt concreta, 6 d’humerals, 6 apicals, i una seta petita al final de la 3a estria (Fig. 2b).
Material estudiat
HOLOTIPUS: ♀, etiquetat: «Avenc de la Cabana d’en Garraba, Pont de Bar (387476, 4689433, 1.312 msnm), L’Alt Urgell, Lleida, Catalunya / 1.312 msnm. 1-VIII-2020, Joan Pallisé i Manel Vives leg.» (etiqueta blanca); «Holotipus/ Speleopidius joanivivesi nov./ E. Vives & J. Comas des.2021» (etiqueta vermella). (dipositat a MCNB).
78 Butlletí ICHN 85 (2), 2021
E. VIVES & J. COMAS
GEA, FLORA ET FAUNA
Figura 1. Speleopidius joanivivesi n. gen. i n. sp. al seu hàbitat natural, dia 3-IV-2021a temperatura de 6 ºC i humitat del 96 %.

Figura 2. Hàbitus de Speleopidius joanivivesi n. gen. i n. sp. holotipus femella: a) visió dorsal (fotografia: Agustí Messeguer), b) èlitres amb senyalització de la quetotàxia de la vora elitral.
PARATIPUS: 1 ♀, «Avenc de la Cabana d’en Garraba, Pont de Bar (387476, 4689433, 1.312 msnm), L’Alt Urgell, Lleida, Catalunya / 1.312 msnm. 3-IV-2021, Joan Pallisé i Eduard Badiella leg.» (etiqueta blanca); «Paratipus/ Speleopidius joanivivesi nov./ E. Vives & J. Comas des. 2021» (etiqueta vermella). (dipositat a CEV).
Material addicional estudiat
«Restes d’uns èlitres, Avenc de la Cabana d’en Garraba, Pont de Bar (387476, 4689433, 1.312 msnm), L’Alt Urgell, Lleida, Catalunya / 1.312 msnm. 8-VII-2018, Joan Pallisé i Ramon Sendra leg.» (etiqueta blanca), «Speleopidius joanivivesi nov./ E. Vives & J. Comas des. 2021» (etiqueta blanca) (dipositat a CJC); «Restes d’uns èlitres i un prostern, Avenc de la Cabana d’en Garraba, Pont de Bar (387476, 4689433, 1.312 msnm), L’Alt Urgell, Lleida, Catalunya / 1.312 msnm. 3-IV-2021, Joan Pallisé i Eduard Badiella leg.» (etiqueta blanca), «Speleopidius joanivivesi nov./ E. Vives & J. Comas des. 2021» (etiqueta blanca) (dipositat a CEV).
Descripció
Longitud de 9 a 10 mm de la punta de les mandíbules a l’àpex abdominal; amplada de 4,0 a 4,2 mm. Aspecte general allargat amb el cap arrodonit i molt convex, i el pronot i èlitres més aplanats. Àpter. Teguments de color marró-vermellosos a causa de la despigmentació adaptativa a la vida hipogea.

Figura 3. Speleopidius joanivivesi n. gen. i n.sp.: a) detall del cap amb els replecs de la zona ocular (fotografia Agustí Meseguer), b) detalls del prostern i del procés prosternal, c) detalls del mesostern, mesocoxes i del procés mesosternal.
Cap arrodonit, gros i amb les temples poc allargades i arrodonides, molt convexes, marcant un coll molt gruixut. Ulls reduïts a una petita àrea grogosa a l’extrem de les temples. El cap és convex i llis en la part superior amb dues fossetes frontals poc marcades i lleugerament arquejades. Epistoma en forma de trapezi amb dues llargues setes a cada angle anterior, solcat transversalment a la seva part anterior. Labre rectangular, lleugerament més ample que llarg, amb la vora anterior enfonsada a la seva meitat en forma de V. Mandíbules grosses, allargades, molt corbades a la punta, amb els costats aplanats en la base i amb una carena sortida a la part lateral inferior. Palps maxiŀlars glabres, estrets i amb el darrer artell ovalat; els palps labials més fins i més curts, amb tres llargues setes al primer artell i sense setes al segon. Llavi ovalat, transvers, molt escotat a la part anterior amb una dent bífida al mig. Coll convex a la part inferior i amb petites arrugues transverses. Dues grans setes supraorbitals. Costats del cap lleugerament convexos i amb arrugues transverses. Cella orbital desdoblada en dues carenes convergents a la base de les antenes. Restes oculars al mig d’una depressió amb arrugues (Fig. 3a). Antenes llargues sobrepassant la base del pronot a partir del vuitè artell. Artells de les antenes pubescents a partir del tercer, que és molt més llarg que el primer i que el quart, la resta subiguals, comprimits i amb tres setes llargues a la part apical.
79 Butlletí ICHN 85 (2), 2021 GEA, FLORA ET FAUNA UN NOU GÈNERE I ESPÈCIE DE MOLOPINA HIPOGEU DEL PIRINEU CATALÀ
a b a b c
GEA, FLORA ET FAUNA
Pronot aplanat, subquadrat poc convex, lleugerament més ample que llarg, la seva màxima amplada és a nivell de les setes laterals anteriors. El costats són sinuats, amples a la meitat anterior i estrets a la posterior. Té els angles anteriors arrodonits i poc sortints cap a endavant, els posteriors són molt marcats i gairebé rectes. El marge anterior lleugerament arquejat i proveït d’un serrell de petites setes daurades, el marge posterior és gairebé recte. La vora del pronot té una llarga seta a la part anterior i una altra just a l’angle posterior. La superfície del disc del pronot té unes fines arrugues transverses als costats i a la base posterior. Prostern (Fig. 3b) molt més ample que llarg, convex, llis i brillant, amb les cavitats procoxals amb una vora gruixuda. Procés prosternal ample i aplanat, ovalat a la seva cara inferior i amb l’àpex arrodonit, en la seva part lateral és convex i s’eixampla per tancar les cavitats procoxals. Mesostern (Fig. 3c) molt estret i profundament deprimit transversalment en la seva part mitjana. Procés mesosternal ample i aplanat, bífid a la seva part posterior. Metastern llarg i estret, aplanat en la zona central. Metaepisternes amagades sota de la vora elitral. Escudet triangular i llis, amb una estriola escutelar a cada costat.
Èlitres (Fig. 2b) llargs i estrets, poc paraŀlels, molt poc estrets a la base i molt arrodonits a la part apical. La zona basal posseeix una vora transversa que gairebé connecta amb l’estriola escutelar. Angles humerals molt arquejats i amb un petit mugró. Les estries elitrals ben marcades i llises, les interestries aplanades i molt finament puntejades a la meitat anterior, sense porus pilífers. Sutura molt fina i sense vora. Zona del disc elitral molt lleugerament convex. Vora lateral molt marcada i una mica aplanada, especialment a la zona apical. Tota la vora elitral proveïda d’una sèrie de setes umbilicades. Epipleures molt amples al seu terç basal, s’estrenyen de forma homogènia fins a l’àpex. Part inferior de l’abdomen amb el tercer esternit visible més ample que els altres, els esternits del 3r al 6è són curts i convexos, amb dues llargues setes centrals a la part posterior, excepte el pigidi que en té quatre. Potes moderadament allargades i fines, amb els profèmurs i els mesofèmurs engruixits a la seva part mitjana, els metafèmurs més llargs i poc engruixits, però amb els metatrocànters allargats com la meitat dels fèmurs. Les tíbies rectes, les protíbies amb un ben desenvolupat òrgan netejador i un gruixut esperó apical. Les mesotíbies i metatíbies amb llargues carenes longitudinals molt irregulars. Els tarses són bastant fins i molt pubescents a la seva cara superior, a la part inferior només amb unes curtes setes inclinades. Ungles llises, arquejades i molt divergents.
Genitàlia de la femella amb gonòpodes dímers (Fig. 4); gonocoxits arquejats, en punta poc aguda i amb dues petites espines laterals externes. El gonosubcoxit té de forma ovalada, bastant irregular. El sac copulador és ample, transparent i té una base piramidal. Espermateca llarga i tubular amb una petita glàndula annexa.
Etimologia
Espècie dedicada a l’eminent entomòleg català Joan Vives Duran (1918-2000), gran especialista en la família Carabidae, que dedicà molt de temps i esforços a la recerca de la fauna cavernícola ibèrica.
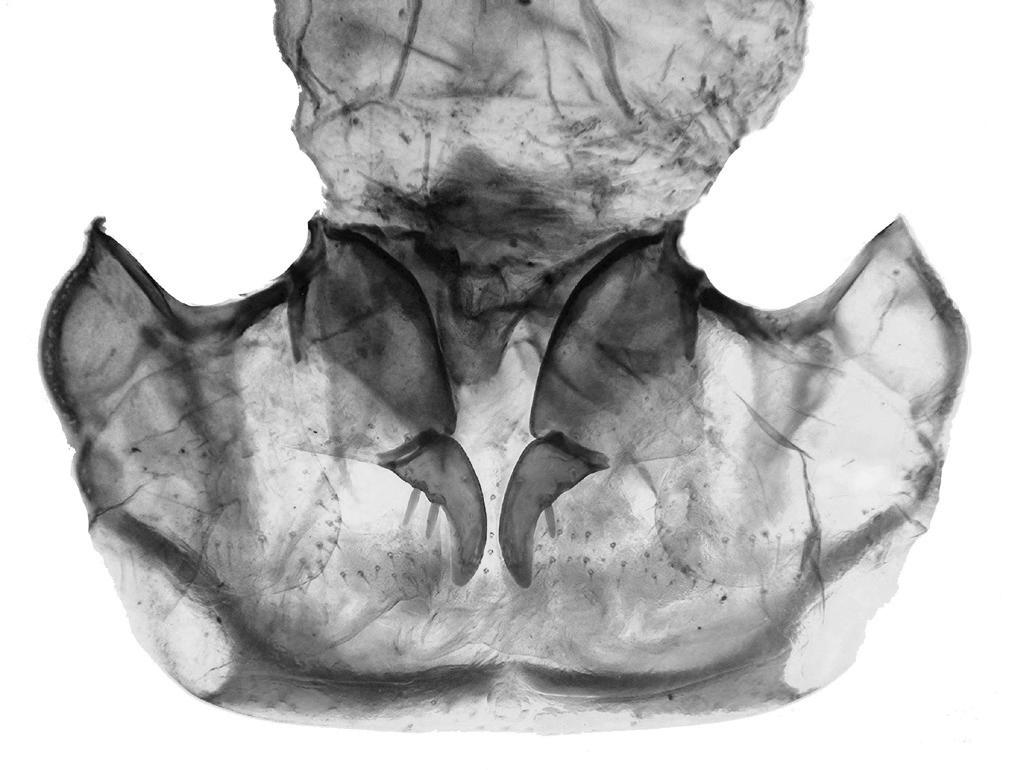
Comentaris
Aquest nou gènere i espècie (Fig. 5a) pertany al grup dels Pteroschini hipogeus de la subtribu Molopina (sensu Casale & Ribera, 2008) que han poblat la part més occidental de l’àrea mediterrània. Tots ells adaptats a la vida subterrània, despigmentats, cecs i amb apèndix allargats, a excepció de Molopidius spinicollis (Dejean, 1828) (Fig. 5b) que es pot considerar una espècie humícola que viu entre la fullaraca dels boscos humits de les fagedes de Catalunya al nord del riu Ter i que rarament es pot trobar a les coves en règim de troglòfil (Jeannel, 1936; Español et al., 1983). Un altre gènere i espècie pobla el nord de l’illa de Mallorca, a les cavitats de Serra de Tramuntana i alentorns; és tracta d’Henrotius jordai (Reitter, 1914) (Fig. 5e) que forma part de les comunitats de fauna cavernícola d’aquesta illa, però ocasionalment es captura al MSS de les zones càrstiques, (Jeannel, 1947; 1950). Speleopidius també està estretament relacionat amb el gènere Zariquieya Jeannel, 1924, que viu a les coves de les comarques gironines sobre tot amb, amb Z. troglodytes (Jeannel, 1924; Negre, 1977) (Fig. 5c), i més allunyat de Z. boumortensis Faille, Fresneda & Bourdeau, 2011, només coneguda de l’Avenc del Pla Fornesa a L’Alt Urgell, (Ortuño & Gilgado, 2010; Mateu, 1955; Jeannel, 1924).
També podem relacionar aquest nou tàxon amb Oscadytes rovirai Lagar, 1975, (Fig. 5d) de les cavitats i MSS del Prepirineu d’Osca, més propi del mitjà intersticial de les zones càrstiques i que accidentalment es pot trobar a les cavitats subterrànies (Lagar, 1975; Fresneda et al., 1997). Finalment podem relacionar el nou gènere amb Speomolops sardous Patrizi, 1955 (Fig. 5f), endemisme de l’illa de Sardenya. tant sols conegut d’unes poques cavitats del centre i l’est de l’illa (Patrizi, 1955; Faille, et al., 2018). La manca de setes a les interestries del disc elitral, allunya força el nou gènere dels altres gèneres de Pterostichini, Pterostichina com Troglorites, Jeannel,1919, Pterostichus subgen. Lianoe Gozis,1882 i Tinautius, Mateu, 1997. Veure Taula 1.
80 Butlletí ICHN 85 (2), 2021
E. VIVES & J. COMAS
Figura 4. Speleopidius joanivivesi n. gen. i n.sp., detall del vuitè segment abdominal amb els gonocoxits i gonòpodes.






81 Butlletí ICHN 85 (2), 2021 GEA, FLORA ET FAUNA f e d c b a
UN NOU GÈNERE I ESPÈCIE DE MOLOPINA HIPOGEU DEL PIRINEU CATALÀ
Figura 5. Hàbitus dels sis gèneres hipogeus de Molopina de la mediterrània occidental: a) Speleopidius joanivivesi n. gen. i n. sp., b) Molopidius spinicollis, c) Zariquieya troglodytes, d) Oscadytes rovirai, e) Henrotius jordai, f) Speomolops sardous
GEA, FLORA ET FAUNA
Clau de determinació dels gèneres de Molopina hipogeus de la Mediterrània occidental
1 Espècies amb setes a la tercera interestria elitral .............2
– Espècies sense setes a la tercera interestria elitral ...........3
2 Espècies amb ulls reduïts però funcionals
..................................................Molopidius Jeannel, 1942 – Espècies sense ulls, reduïts a una cicatriu Henrotius Jeannel, 1950
3 Dues o tres setes al marge pronotal, una seta prescutelar
...................................................... Oscadytes Lagar, 1975
– Una sola seta al marge pronotal, sense seta prescutelar ...4
4 Tarses sense pilositat a la cara superior Speomolops Patrizi, 1955
– Tarses amb pilositat a la cara superior .............................5
5 Procés prosternal truncat a la part posterior, sense estriola escutelar Zariquieya Jeannel, 1924
– Procés prosternal arrodonit a la part posterior, estriola escutelar present ....................................Speleopidius n. gen.
Agraïments
Aquest treball no hauria estat possible sense la col·laboració d’una colla d’amics i coŀlegues que han aportat material, collaboracions i coneixements. Especialment al bon amic i bioespeleòleg Joan Pallisé, al company i fotògraf, Agustí Messeguer, als companys de la Secció d’Investigacions Subterrànies del C.E.T., Eduard Badiella, Rebeca Calvo i Manel Vives, i molt especialment a Alberto Tinaut, (Universidad de Granada) que ens ha facilitat important material de comparació i un agraïment també a José Serrano (Universidad de Murcia) per les seves aportacions a la millora del manuscrit.
Bibliografia
Casale, A. & Ribera, I. 2008. Are Molopina of the EuroMediterranean region related to the Madagascan, South African and Australian Pterostichini? (Coleoptera, Carabidae). Biogeographia, 29: 33-44.
Español, F. & Bellés, X. 1980. Armidia unicolor Dult. (Col. Cantharidae). un curioso elemento de nuestra coleopterofauna cavernícola. Munibe, 32 (3-4): 281-282.
Español, F., Escolà, O. & Vives, E. 1983. Alguns invertebrats interessants del domini subterrani gironí. Revista de Girona, 102: 41-44.
Faille, A., Fresneda, J. & Bourdeau, C. 2011. Les Molopina hypogés des Pyrénées avec la description d’une nouvelle espèce de Zariquieya Jeannel, 1924 d’Espagne (Insecta, Coleoptera, Carabidae, Pterostichini). Zoosystema, 33 (4): 429-441.
Faille, A., Casale, A., Hernando, C., Mouloud, S. A. & Ribera, I. 2018. Tectonic vicariance versus messinian dispersal in western mediterranean ground beetles. Zoologica Scripta, 47: 565-581.
Fresneda, J., Hernando, C., Lagar, A. & Ortuño, V. 1997. Sistemática y geonemia de un coleóptero subterráneo de España Oscadytes rovirai Lagar, 1975. (Coleopotera: Pterostichidae).Annales de la Societé entomologique de France, 33 (2): 205-213.
Jeannel, R. 1924. Coléoptères nouveaux de Catalogne. Trabajos del Museo de Ciencias Naturales de Barcelona, 4 (8): 3-18.
Jeannel, R. 1936. Sur la Feronia spinicollis de Dejean (Coleopt. Carabidae). In: Livre Jubilaire. Societé entomologique de France. Ed. E.L. Bouvier. P. 213-217.
Jeannel, R. 1947. Le peuplement des Pyrénées. Revue Française d’Entomologie, 14 (1): 53-104.
Jeannel, R. 1950. Sur deux Ptérostichides cavernicoles de Majorque Revue Française d’Entomologie.17 (3): 157-165.
Lagar, A. 1975. Nuevos coleópteros troglobios de la provincia de Huesca. Espelosie, 17: 41-47.
Mateu, J. 1955. Los Molopini Bon. de la Península Ibérica. Eos, Madrid, 31: 297-301.
Negre, J. 1977. Sur les Molopini hypogés de Catalogne. Comunicacions del 6è. Simposium d’Espeleologia. Biospeleologia. Terrassa. P.139-141.
Ortuño, V. M. & Gilgado, J. D. 2010. Update of the knowledge of the Ibero-Balearic hypogean Carabidae (Coleoptera): Faunistics, biology and distribution. Entomological Blätter, 106: 233-264.
Pallisé, J. 2020. Coleòpters Cavernícoles de Catalunya. Centre Excursionista de Terrassa. Omnia Books. 107 p.
Patrizi, S. 1955. Nuovo genere e nuova specie di Pterostichidae troglobio della Sardegna orientale. Fragmenta Entomologica, 2(4): 29-40.
Serrano, J. 2013. New Catalogus of the family Carabidae of the Iberian Peninsula (Coleoptera). Ed. Universidad de Murica, 192 p.
Serrano, J. 2020. Catálogo electrónico de los Cicindelidae y Carabidae de la Península Ibérica (Coleoptera, Caraboidea). Monografías electrónicas de la S.E.A., 9. Sociedad Entomológica Aragonesa. 10+32+44 p.
Taula 1. Caràcters principals que separen el gènere Speleopidius dels altres cinc gèneres de Molopina mencionats.
Henrotius Molopidius Zariquieya Oscadytes Speleopidius Speomolops
Estriola juxta–escutelar X X – – X –
Àpex del prostern truncat X X X X – X
Setes al disc elitral X X – – – –
Disc elitral convexa X X – – X –
Temporals estriats – – X X X X
Temporals amb protuberància cònica – X X – – –
Ulls funcionals – X – – – –
Labre quadrat – X X – – –
Carenes tibials – – X – X –
Base pronotal escotada – – X X – X
Àpex elitral truncat – – X X – –
Tarses pilosos per sobre
– – X X X –
Part inferior del cap llisa X X – – – X
Meta–trocantins allargats
82 Butlletí ICHN 85 (2), 2021
E. VIVES & J. COMAS
– X X X X X
Butlletí
Institució Catalana d’Història Natural, 85 (3): 83-85. 2021
NOTA BREU
ISSN
On the presence of a small population of Sargassum trichocarpum J. Agardh (Phaeophyceae: Fucales) in Catalonia (Northwestern Mediterranean)
Sobre la presència d’una petita població de Sargassum trichocarpum J. Agardh (Phaeophyceae: Fucales) a Catalunya (Mediterrània nord-occidental)
Enric Ballesteros* & Boris Weitzmann**
* Centre d’Estudis Avançats de Blanes-CSIC. Acc. Cala Sant Francesc, 14. 17300 Blanes, Girona.
** PNMMBT. Passeig del Port, Espigó de Llevant, s/n. 17258 L’Estartit, Girona.
Corresponding author: Enric Ballesteros. A/e:kike@ceab.csic.es
Rebut: 17.07.2021. Acceptat: 28.08.2021. Publicat: 30.09.2021
Members of the order Fucales (Phaeophyceae) are canopy-forming algae that constitute different marine habitats at the intertidal, infralittoral and upper circalittoral zones from all the oceans and main enclosed seas on Earth (Boudouresque et al., 2017). The genus Sargassum C. Agardh is one of the genera with more described specific entities. Besides being a major component of the flora from tropical to subtropical marine environments, the genus Sargassum is also present in the Mediterranean Sea, where up to nine species have been reported (Cormaci et al., 2012; Aouissi et al., 2018). Amongst those species, Sargassum trichocarpum J. Agardh is considered a Mediterranean endemism (Giaccone, 2003) with a very restricted distribution in the western Mediterranean basin (Thibaut et al., 2016; Aouissi et al., 2018). Here we report on a new population of this species found in northern Catalonia, which adds a new locality for this scarce taxon in the northwestern Mediterranean.
The population was found in July 12th 2021 at the northern side of Cala Montgó in the MPA of the Parc Natural del Montgrí, illes Medes i Baix Ter (L’Escala, Girona, 42.107124ºN, 3.175160ºE) growing over a flat calcareous rocky bottom surrounded by sediment, between 20.4 and 21.1 m depth. Most specimens were heavily covered by silt (Fig. 1). The rocky bottom hold a dense population of Ericaria zosteroides (C. Agardh) Molinari-Novoa & Guiry (habitat: «Algal-dominated circalittoral rock with Ericaria zosteroides and Gongolaria montagnei», LPRE code 0302010101) (Ballesteros, 2019) with other common species including the erect algae Dictyota sp., Phyllophora crispa (Hudson) P.S. Dixon, Zanardinia typus (Nardo) P.C. Silva, Arthrocladia villosa (Hudson) Duby, Dudresnaya verticillata (Withering) Le Jolis and Chrysymenia ventricosa (J.V. Lamouroux) J. Agardh, and the encrusting red algae Mesophyllum alternans (Foslie) Cabioch & M.L. Mendoza, Neogoniolithon mamillosum (Hauck) Setchell & Mason, Peyssonnelia rosa-marina Boudouresque & Denizot and Lithophyllum stictiforme (Areschoug) Hauck. Sargassum trichocarpum was absent inside the dense cano-
py of E. zosteroides and was mainly present in the margins of the rocky outcrop, in places partially covered or situated very close to the sediment. We counted approximately 200 specimens, most of them reduced to the main axis and small branches with divided leaves. Only a few individuals showed long primary branches with fertile structures.

Completely developed individuals measured 25-40 cm long, with one to three cylindrical main axis of 2-3 cm arising from a basal disc 8-12 mm in diameter, strongly attached to rock (Fig. 2). Primary branches cylindrical, smooth, non muriculate, up to 35 cm long. Leaves with a conspicuous midrib, those present in the basal part of the primary branches always divided up to five times, 4-6 mm wide, up to 8 cm long, wavy, denticulate with blunt tips. Leaves from the upper part of the branches were much smaller and thinner, usually undivided. Aerocysts spherical, usually pedicellate, 3-5
83 Butlletí ICHN 85 (3), 2021
BREU
NOTA
de la
2013-3987 (online edition): ISSN: 1133-6889 (print edition))
DOI: 10.2436.20.1502.01.77
Figure 1. In situ aspect of the specimens of Sargassum trichocarpum found in Cala Montgó.

mm in diameter, sometimes mucronate. Fertile branches 2-6 cm long, with cylindrical receptacles 1 mm in diameter, up to 2 cm long, divided 1-4 times (Fig. 3). Three vouchers have been deposited at the Herbarium of the University of Girona (HGI-A 20877, HGI-A 20878, HGI-A 20879).
The closest records of Sargassum trichocarpum come from Martigues-Ponteau (Provence, France) (Verlaque, 1977) and Sausset les Pins (Verlaque & Boudouresque, 1981) situated 200 km away, where it was extremely rare around forty years ago and has not been collected again since 1982 (Thibaut et al., 2016). The only other northwestern Mediterranean records are located in Columbretes islands (Gómez-
Garreta et al., 2001) where, although uncommon, the species has been observed in different diving spots (EB, unpublished data). No other localities are known from the northwestern Mediterranean, which indicates the rarity of this species. In fact, S. trichocarpum is included in the Annex II of the Barcelona Convention (Verlaque et al., 2019), which establishes a list of Mediterranean endangered or threatened species. Given that most species of the genera Sargassum seem to be in steady decline across most of the Mediterranean Sea (see Thibaut et al., 2016) and the rarity of S. trichocarpum in the northwestern Mediterranean, the monitoring of this population through time and the searching for other possible locations in the nearby rocky outcrops is highly recommended.
Acknowledgements
Funding was provided by Fundació Alive. Thanks are due to Andreu Pujades and Patricia Besem for their assistance during the scuba surveys.
References
Aouissi, M., Sellam, L. N., Boudouresque, C. F., Blanfuné, A., Derbal, F., Frihi, H., Perret-Boudouresque, M., Rebzani-Zahaf, C., Verlaque, M. & Thibaut, T. 2018. Insights into the species diversity of the genus Sargassum (Phaeophyceae) in the Mediterranean Sea, with a focus on a previously unnoticed taxon from Algeria. Mediterranean Marine Science, 19: 48-57.
Ballesteros, E. 2019. Llista dels hàbitats marins presents a Catalunya i la seva correspondència amb altres classificacions. Generalitat de Catalunya. Departament de Territori i Sostenibilitat. Barcelona. 40 p.
84 Butlletí ICHN 85 (3), 2021 NOTA BREU E. BALLESTEROS & B. WEITZMANN
Figure 2. Specimen of Sargassum trichocarpum collected in Cala Montgó. Bar = 5 cm.
Figure 3. Close-up of aerocysts and receptacles heavily epiphytized by hydroids.
Boudouresque, C. F., Blanfuné, A., Harmelin-Vivien, M., Personnic, S., Ruitton, S., Thibaut, T. & Verlaque, M. 2017. Where seaweed forests meet animal forests: the examples of macroalgae in coral reefs and the Mediterranean coralligenous ecosystem. P. 369-396. In: Rossi, S.; Bramanti, L.; Gori, A.; Orejas C. J. (eds.). Marine animal forests. Springer International Publishing. Switzerland. 1366 p.
Cormaci, M., Furnari, G., Catra, M. & Alongi, G. 2012. Flora marina bentonica del Mediterraneo: Phaeophyceae. Bollettino dell’Accademia Gioenia di Scienze Naturali, 45: 1-508.
Giaccone, G. 2003. Biodiversity of the Mediterranean Sea: an introductory speech to the Marine algae Symposium. Bocconea Palermo, 16: 182-192.
Gómez-Garreta, A., Barceló-Martí, M. C., Pérez-Ruzafa, I. M., Ribera-Siguan, M. A. & Rull-Lluch, J. 2001. Flora Phycologica Iberica. Vol. 1. Fucales. Universidad de Murcia. Murcia. 192 p.
Thibaut, T., Blanfuné, A., Verlaque, M., Boudouresque, C. F. & Ruitton, S. 2016. The Sargassum conundrum: highly rare, threatened or locally extinct in the NW Mediterranean and still lacking protection. Hydrobiologia, 781: 25-42.
Verlaque, M. 1977. Étude du peuplement phytobenthique au voisinage de la centrale thermique de Martigues-Ponteau (golfe de Fos, France). Ph. D. Thesis. Université d’Aix-Marseille II. France. 172 p.
Verlaque, M. & Boudouresque, C. F. 1981. Végétation marine de la Corse (Méditerranée). V. Documents pour la flore des algues. Biologie: Écologie méditerranéenne, 8: 139-156.
Verlaque, M., Boudouresque, C. F. & Perret-Boudouresque, M. 2019. Mediterranean seaweeds listed as threatened under the Barcelona Convention: a critical analysis. Scientific Reports PortCros national Park, 33: 179-214.
85 Butlletí ICHN 85 (3), 2021 NOTA BREU ON THE PRESENCE OF A SMALL POPULATION OF SARGASSUM TRICHOCARPUM J. AGARDH
Butlletí de la Institució Catalana d’Història Natural, 85 (3): 87-90. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print
NOTA BREU
Osmoderma eremita (Scopoli, 1763) al Paratge Natural d’Interès Nacional de l’Albera (Coleoptera: Scarabaeidae: Cetoniinae) amb notes sobre altres coleòpters saproxílics interessants
Osmoderma eremita (Scopoli, 1763) at the «Paratge Natural d’Interès Nacional de l’Albera» (Coleoptera: Scarabaeidae: Cetoniinae) with notes on other interesting saproxylic beetles
Josep Muñoz-Batet*, Joaquim Soler* & Amador Viñolas*
* Museu de Ciències Naturals de Barcelona. Laboratori de Natura. Coŀlecció d’artròpodes. Passeig Picasso, s/n. 08003 Barcelona.
Autor per a la correspondència: Josep Muñoz-Batet. A/e: pepmba@gmail.com
Rebut: 15.08.2021. Acceptat: 31.08.2021. Publicat: 30.09.2021
La direcció del Paratge Natural d’Interès Nacional de l’Albera (PNIN), Girona, ens va soŀlicitar efectuar un mostreig de las fagedes (Fagus sylvatica L.) de la zona de protecció integral del PNIN, per veure la població de coleòpters saproxílics existents i, sobretot, la possibilitat de poder localitzar poblacions d’Osmoderma eremita (Scopoli, 1763) considerada l’espècie saproxílica per exceŀlència i màxima indicadora de la maduresa d’un bosc.
Arran d’aquesta petició i desprès de reunir-nos amb els tècnics del PNIN, es decidí començar els mostrejos per las fagedes ubicades en la zona de Requesens, en el terme municipal de la Jonquera.
Es varen mostrejar un total de nou fagedes (Fig. 1) amb la col·locació d’un total de 30 paranys d’intercepció tipus po-
L’O. eremita es una espècie present a la Directiva Hàbitat (Galante & Verdú, 2000), amb la classificació UICN de «espècie vulnerable» (VU A1c), a l’«Atlas y Libro Rojo de los Invertebrados Amenazados de España (Especies Vulnerables)» (Micó et al., 2011) amb la categoria UICN per l’Espanya «VU B1ab(i,ii,iii) i la categoria UICN mundial «NT», i a la llista vermella dels coleòpters saproxílics del Mediterrani (García et al., 2018) amb la categoria IUCN «NE».
87 Butlletí ICHN 85 (3), 2021 NOTA BREU
edition)
DOI: 10.2436.20.1502.01.78
Figura 1. Mapa de les fagedes estudiades en el PNIN amb indicació de la situació dels paranys.
lytrap amb esquer, que es van mantenir actius des del 15 de maig fins el 5 d’agost, revisant-los setmanalment.
Dintre del nombrós material recoŀlectat de coleòpters, encara en fase de determinació, el 20 de juliol es varen capturar dos exemplars mascles d’Osmoderma eremita (Fig. 2) en els paranys 21.2 (Xalami) i 35.3 (Fageda de Roc Colom) respectivament; un dels exemplars estava mort i l’altre que estava viu es va alliberar. Aquesta troballa indica l’alt valor ecològic de les fagedes de la zona de protecció integral del PNIN i alhora confirma la sospita de que es pogués trobar l’O. eremita al PNIN donada la seva presència a la Reserva Natural Nacional del Bosc de la Maçana situada a la vessant nord del Massís de l’Albera i a uns 6 km de distancia en línia recta.
L’espècie fou recoŀlectada per primer cop a Catalunya l’any 1951 per Español i Mateu en una vella fageda situada en una de les vessants del Matagalls, concretament a la zona de Sant Segimon, Viladrau (Girona, Parc Natural del Montseny) (Micó et al., 2011), exemplars que es troben dipositats en la coŀlecció del Museu de Ciències Naturals de Barcelona. Des d’aquelles dates l’espècie no ha estat localitzada en altres indrets de Catalunya, essent els exemplars de l’Albera la segona cita catalana.

Així mateix, es citen a continuació altres coleòpters interessants recoŀlectats al PNIN de l’Albera, un d’ells, nou per a la Península que també s’ha localitzat a la província de Barcelona.
Família Eucnemidae Eschscholtz, 1829
Dromaeolus barnabita (A. Villa & J. B. Villa 1838)
Material estudiat
5 ex., etiquetats: 2 ex. «2-VII-2011, Pla de les Voltes, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.» amb polytrap; 1 ex. «2-VII-2011, Torrent de la Vernosa, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.» amb parany de finestra; 2 ex. «30-VII-2016, Corral del Sorrer, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.» amb parany de finestra.
Recalde Irurzun (2008) cita l’espècie per primer cop d’Espanya amb exemplars recoŀlectats a Araba i Navarra, els de la Jonquera són la primera cita per a Catalunya.
Família Cleridae Latreille, 1802
Opilo pallidus (Olivier 1795)
Material estudiat
3 ex., etiquetats: 1 ex. «18-VII-2011, Mas Boencs, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.» amb polytrap; 2 ex. «4-IX-2017, Mas Gamarús, Espolla, l’Albera, Girona, J. Muñoz & J. Soler leg.» amb parany de finestra.
Bahillo de la Puebla & López Colón (2000) citen la espècie de San Sebastián i dels Pirineus orientals francesos, aquesta última reflectida en el catàleg de Fuente (1931), essent les d’Espolla i la Jonquera les primeres cites de la espècie per a Catalunya.
Família Biphyllidae LeConte, 1861
Biphyllus frater (Aubé 1850) (Fig. 3)
Material estudiat
5 ex., etiquetats: 3 ex. «30-V-2021, Roc de l’Atalaidor, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.» amb polytrap; 1 ex. «6-VII-2021, Fageda de Roc Colom, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.» amb polytrap; 1 ex. «Puig de les Canals, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.» amb polytrap.
Recalde Irurzun & San Martín Moreno (2014) citen l’espècie del Parc Natural de Urbasa-Andía (Navarra), única localitat peninsular coneguda de la espècie. Els exemplars del PNIN de l’Albera (Girona) són la primera localitat de Catalunya i la segona ibèrica.
Família Ciidae Leach, 1819
Ropalodontus baudueri Abeille 1874 (Fig. 4)
Material estudiat
88 Butlletí ICHN 85 (3), 2021
J. MUÑOZ-BATET ET AL.
NOTA BREU
Figura 2. Habitus d’Osmoderma eremita (Scopoli, 1763).



1 ex., etiquetat: «6-VII-2021, Fageda de Roc Colom, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.» amb polytrap.
Espècie ben separada de R. perforatus (Gyllenhal, 1813) per la puntuació del protòrax i èlitres, així com per la longitud del tercer i quart artell de les antenes i pròpia de la mediterrània (Rose & Zagatti, 2016). Espècie ja coneguda de la península Ibèrica per dos exemplars de Masquefa (Barcelona) (Viñolas et al., 2016). Els exemplars de la Jonquera (Girona) són la segona cita de l'espècie per a la Península.
Família Melandryidae Leach, 1815
Osphya aeneipennis Kriechbaumer, 1848 (Fig. 5)
Material estudiat
1 ex., etiquetat: «30-V-2021, els Cavallets, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.» amb polytrap.
Viñolas et al. (2014) van cita per primer cop l’espècie per a la península Ibèrica amb un exemplar recoŀlectat a Saldes (Barcelona, Parc Natural del Cadí-Moixeró). El present exemplar de la Jonquera (Girona, PNIN) es la segona cita ibèrica i per a Catalunya.
89 Butlletí ICHN 85 (3), 2021 NOTA BREU OSMODERMA EREMITA (SCOPOLI, 1763) AL PARATGE NATURAL D'INTERÈS NACIONAL DE L'ALBERA
Figura 3. Habitus del Biphyllidae Biphyllus frater (Aubé 1850).
Figura 4. Habitus del Ciidae Ropalodontus baudueri Abeille 1874.
Figura 5. Habitus del Melandryidae Osphya aeneipennis Kriechbaumer, 1848.
NOTA BREU
Agraïments
A Bartomeu Borras, director, i Pau Federico i Marc Fernández, tècnics del Paratge Natural d’Interès Nacional de l’Albera, pels ajuts i esforços realitzats en les recoŀleccions sistemàtiques realitzades en les diferents espais zones prospectades. A Conxita Esteba, propietària de Requesens, per la facilitat d’accés que sempre ens proporciona.
Bibliografia
Bahillo de la Puebla, P. & López Colón, J. I. 2000. El género Opilo Latreille, 1802 en la Península Ibérica (Coleóptera, Cleridae). Boletín de la Asociación española de Entomología, 24 (1-2): 213227.
Fuente, J. M. de la 1931. Catálogo sistemático-geográfico de los coleópteros observados en la Península Ibérica, Pirineos propiamente dichos y Baleares (Continuación). Boletín de la Sociedad entomología Española, 14: 147-153. Galante, E. & Verdú, J. R. 2000. Los Artrópodos de la “Directiva Hábitat” en España. Organismo Autónomo Parques Nacionales. Ministerio de Medio Ambiente. Madrid. 247 p.
García, N., Numa, C., Bartolozzi, L., Brustel, H., Buse, J., Norbiato, M., Recalde, J. I., Zapata, J. L., Dodelin, B., Alcázar, E., Barrios, V., Verdugo, A., Audisio, P., Micó, E., Otero, J. C., Bahillo, P., Viñolas, A., Valladares, L., Méndez, M., El Antry, S. & Galante, E.
2018. The conservation status and distribution of Mediterranean saproxylic beetles. Malaga, Spain: IUCN. xii + 58 p.
Micó, E., Murria, E. & Galante, E. 2011. Osmoderma eremita (Scopoli, 1763). P. 215-220. En: Verdú, J. R., Numa, C. & Galante, E. (eds.). Atlas y Libro Rojo de los Invertebrados amenazados de España (Especies Vulnerables). Dirección General de Medio Natural y Política Forestal, Ministerio de Medio Ambiente, Medio Rural y Marino. Madrid. 719 p.
Recalde Irurzun, J. I. 2008. Elementos para el conocimiento de los eucnémidos del norte de España, y actualización del catálogo de especies ibéricas (Coleoptera: Elateroidea: Eucnemidae). Heteropterus Revista de Entomología, 8 (2): 233-252.
Recalde Irurzun, J. I. & San Martín Moreno, A. F. 2014. Algunos Cucujoidea del norte de Navarra nuevos o poco conocidos para la fauna ibérica (Coleoptera: Endomychidae, Cerylonidae, Biphyllidae, Laemophloeidae & Erotylidae). Arquivos Entomolóxicos, 10: 175-180.
Rose, O. & Zagatti, P. 2016. Les Ciidae de la faune de France continentale et de Corsé: clé d’identification illustrée des espèces (Coleóptera Tenebrionoidea). L’Entomologiste, 72 (5): 287-306.
Viñolas, A., Muñoz-Batet, J. & Soler, J. 2016. Noves o interessants localitzacions d’espècies de coleòpters per a la península Ibèrica i illes Canàries (Coleoptera). Butlletí de la Institució Catalana d’Història Natural, 80: 101-112.
Viñolas, A., Muñoz Batet, J., Masó. G. & Soler, J. 2014. Nuevos registros de coleópteros interesantes del Parque Natural del CadíMoixeró, Cataluña (Península Ibérica) (Coleoptera). Arquivos Entomolóxicos, 12: 91-96.
90 Butlletí ICHN 85 (3), 2021
J. MUÑOZ-BATET ET AL.
Butlletí de la Institució Catalana d’Història Natural, 85 (3): 91-109. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition)
Revisión de la familia Salpingidae Leach, 1815 en el área iberobalear (Coleoptera)
Amador Viñolas*, José Ignacio Recalde Irurzun** & Josep Muñoz-Batet*
* Museu de Ciències Naturals de Barcelona. Laboratori de Natura. Coŀlecció d’artròpodes. Passeig Picasso s/n. 08003 Barcelona. Catalunya. ** C/ Andreszar, 21. 31610 Villava-Atarrabia, Navarra. A/e: jirecalde93@gmail.com
Autor para la correspondencia: Amador Viñolas. A/e: av.rodama@gmail.com
Rebut: 18.08.2021; Acceptat: 01.09.2021; Publicat: 30.09.2021
Resumen
Se revisa y actualiza la fauna de la familia Salpingidae Leach, 1815 en la Península Ibérica e Islas Baleares (13 especies pertenecientes a 6 géneros). Junto con las claves de separación genérica y específica se relacionan los ejemplares estudiados o revisados, se da una breve descripción de cada especie, se comenta su distribución conocida y se aportan datos sobre su biología. Se representa el habitus y el mapa de distribución de cada una de las especies. Asimismo se establece la nueva sinonimia de Aglenus brunneus chilensis Dajoz, 1969 n. syn. con Aglenus brunneus (Gyllenhal, 1813).
Palabras clave: Coleoptera, Salpingidae, revisión, nuevas citas, Península Ibérica, Islas Baleares.
Abstract
Review of the family Salpingidae Leach, 1815 from the Iberian-Balearic area (Coleoptera)
The fauna of Salpingidae Leach family, 1815 in the Iberian Peninsula and the Balearic Islands (13 species appertaining to 6 genera) is reviewed and updated. Along with the generic and specific separation keys, the specimens studied or reviewed are listed, a brief description of each species is given, its known distribution is discussed, and data on its biology are provided. The habitus and the distribution map of each species are represented. The synonymy of Aglenus brunneus chilensis Dajoz, 1969 n. syn. with Aglenus brunneus (Gyllenhal, 1813) is established.
Key word: Coleoptera, Salpingidae, review, new records, Iberian Peninsula, Balearic Islands.
Resum
Revisió de la família Salpingidae Leach, 1815 de l’àrea iberobalear (Coleoptera)
Es realitza una revisió i actualització de la fauna de la família Salpingidae Leach, 1815 a la península Ibèrica i illes Balears, constituïda per 13 espècies pertanyents a 6 gèneres. Juntament amb les claus de separació genèrica i específica es relacionen els exemplars estudiats o revisats, es dóna una breu descripció de cada espècie, es comenta la seva distribució coneguda i s’aporten dades sobre la seva biologia. Es representa l’habitus i el mapa de distribució de cadascuna de les espècies. Així mateix s’estableix la sinonímia d’Aglenus brunneus chilensis Dajoz, 1969 n. syn. amb Aglenus brunneus (Gyllenhal, 1813).
Paraules clau: Coleoptera, Salpingidae, revisió, noves cites, península Ibèrica, illes Balears. urn:lsid:zoobank.org:pub:EE94BC1A-0A25-4221-9807-F587D87325EE
Introducción
Hasta el trabajo de Pérez Moreno (2005) la familia Salpingidae no había sido objeto de trabajo concreto alguno dentro del ámbito Iberobalear. Este trabajo sirvió de referencia de partida para posteriores aportaciones que permitieron clarificar la asignación específica de algunos individuos y/o añadieron nuevas especies al elenco conocido: Salpingus tapirus (Abeille de Perrin, 1874), Lissodema denticolle (Gyllenhal, 1813) y Sphaeriestes stockmanni (Biström, 1977), y así como
ampliar la incipiente distribución conocida de las especies de esta familia en el área mencionada (Lencina et al., 2008; Viñolas et al., 2018). Los avances en el conocimiento de esta familia y las nuevas citas que aportamos en este trabajo permiten actualizar la fauna Iberobalear junto con la distribución conocida de las especies que la integran.
También se clarifica la confusión existente en la distribución de Sphaeriestes reyi (Abeille de Perrin, 1874) provocado por las determinaciones de Cuni i Martorell & Martorell i Peña (1876) y Fuente (1932).
91 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA GEA, FLORA ET FAUNA
DOI: 10.2436.20.1502.01.79
GEA, FLORA ET FAUNA A.
En la presente nota se dan las claves de separación de subfamilias, géneros y especies. De cada una de las especies se relaciona el material estudiado y el revisado, se acompaña una breve descripción de cada una de las tratadas, comentándose los datos que se tienen sobre su biología, se representa el habitus de cada una de ellas y se proporciona el mapa con la distribución de todas las especies conocidas en el área de estudio.
Además se ha estudiado la subespecie de Aglenus brunneus (Gyllenhal, 1813) descrita por Dajoz (1969) de Chile y a la vista de los datos obtenidos en el estudio de 173 especímenes de diferentes procedencias se ha podido observar, tal como se indica en el estudio de la especie, que Aglenus brunneus chilensis Dajoz, 1969 n. syn. de Aglenus brunneus (Gyllenhal, 1813).
Material y métodos
Para la realización del estudio se han revisado los ejemplares depositados en el Museu de Ciències Naturals de Barcelona (MZB) y en el Museo Nacional de Ciencias Naturales de Madrid (MNCN), así como los depositados en diferentes colecciones particulares. También se ha obtenido un elevado número de ejemplares en las recolecciones realizadas en diferentes proyectos, muchos de ellos en áreas protegidas. Se ha revisado toda la bibliografía existente sobre la familia relacionada con el área de estudio, junto con diversos trabajos sobre la fauna europea de Salpingidae.
Las fotografías se han realizado con una cámara Canon® model EOS 760D, con objetivo de microscopía mediante el método de capas, con tratamiento de las imágenes obtenidas mediante el programa Zerene Stacker®. Los dibujos se han
realizado con el programa de Adobe® Ilustrator CS5, con la obtención de archivos PostScript® 3™.
Resultados
Familia Salpingidae Leach, 1815
Clave de subfamilias
1 Especie anoftálmica; áptera; maza antenal de tres artejos, bien diferenciada (Fig. 1); base del protórax tan ancha como la de los élitros (Fig. 9); todos los tarsos de cuatro artejos; edeago con el lóbulo medio más corto que la base, ésta provista de lóbulos tan largos como el medio (Fig. 3) ............................................................Agleninae – Especies con los ojos bien desarrollados y salientes: aladas; antenas sin maza terminal, sólo con los últimos artejos, en número variable, más o menos engrosados progresivamente, en ocasiones diferenciando una maza terminal (Fig. 2); base del protórax más estrecha que la de los élitros (Fig. 19); pro- y mesotarsos de cinco artejos, metatarsos de cuatro; edeago con el lóbulo medio mucho más largo que la base, ésta desprovista de lóbulos (Fig. 4) .........................................................Salpinginae
Subfamila Agleninae Horn, 1878
Género Aglenus Erichson, 1845
Aglenus Erichson, 1845. Nat. Ins. Deutsch.: 285
Especie tipo: Hypophloeus brunneus Gyllenhal, 1813.
92 Butlletí ICHN 85 (3), 2021
ET AL.
VIÑOLAS
1 8 6 7 5 4 3 2
Figuras 1-8. Antena de: 1) Aglenus; 2) Vincenzellus. Edeago de: 3) Aglenus (redibujado de Dajoz, 1974: Bull. Soc. Ent. Fr., 74 (9-10)); 4) Vincenzellus (redibujado de Spilman, 1952; The Coleopt. Bull., 6 (1)); Cabeza de: 5) Lissodema; 6) Sphaeriestes; 7) Rabdocerus; 8) Salpingus.

Aglenus brunneus (Gyllenhal, 1813) (Fig. 9)
Hypophloeus brunneus Gyllenhal, 1813. Ins. Suec. descr. Class, 1 (3): 711
Aglenus obsoletus Shuckard. 1839. Elem. Brist. Ent.: 249
Aglenus major Schaufuss, 1882. Ann. Soc. Ent. Fr., (6) 2: 48
Aglenus brunneus var. rugipennis Schaufuss, 1882. Ann. Soc. Ent. Fr., (6) 2: 48
Anommatus basalis var. rocellae Ragusa, 1892. Nat. Sic., 11 (1891-1892): 137
Aglenus brunneus var. longior Pic, 1923. Echange, 39 (413): 9
Aglenus brunneus chilensis Dajoz, 1969: Bull. Soc. Ent. Fr., 74 (9-10): 230. n. syn.
Material estudiado
128 ej., etiquetados: 1 ej. «424. Aglenus brunneus, Paris / Fairm» «ex col. Martorell i Peña» «75-5892 / MZB»; 2 ej. «Pego, Alicante, Torres Sala» «75-5903 / MZB»; 75 ej. «To-
tana, Murcia, Balaguer» «75-5888 / MZB»; 15 ej. «Totana, Murcia, Balaguer» «75-5893 / MZB»; 4 ej. «VIII-32, Barcelona, S. Gervasi, en un palomar, Montrós leg.» «75-5906 / MZB»; 1 ej. «10-IV-38, Porto-Pi, sota les escorces de pi» «75-5882 / MZB»; 2 ej. «IX-40, Cadaqués, Zaraquiey leg.» «75-5905 / MZB»; 1 ej. «VI-44, Caparroso, Navarra» «755898 / MZB»; 3 ej. «VI-44, Caparroso, Navarra, raíces olivo» «75-5902 / MZB»; 1 ej. «XII-44, Bonanova, (Barna), raíces dalia, Español leg.» «75-5887 / MZB»; 23 ej. «20-XII1960, Los Rodeos, Tenerife, Canarias, tamizado, F. Español leg.» «75-5879 / MZB; 75-5880 / MZB; 75-5881 / MZB»; 3 ej. «20-XII-1960, Los Rodeos, Tenerife, Canarias, F. Español leg.». Depositados en la colección del Museu de Cièncias Naturals de Barcelona y de A. Viñolas.
Material revisado
45 ej., etiquetados: 7 ej. «Pozuelo de Calatrava (Ciudad Real) Lafuente» «Ex col. A. Codina» «75-5894 / MZB»; 1 ej. «Italia, I. Boyer» «75-5897 / MZB»; 1 ej. «Wien, Umgebg» «Ex col. Müller» «75-5891 /MZB»; 1 ej. «Wien, Umgebung» «Ex col. Müller» «75-5889 / MZB»; 1 ej. «Wien, Umgebung» «Ex col. Müller» «75-5890 / MZB»; 1 ej. «Peney» «Ex. col. Martorell i Peña» «75-5899 / MZB»; 1 ej. «Env. de Toulon, Mol. de Boissy» «75-5878 / MZB»; 3 ej. «Cala Fornells, Rosas» «Prov. Gerona, Dr. Zariquiey» «75-5883 / MZB»; 1 ej. «Cala Fornells, Rosas» «Prov. Gerona, Dr. Zariquiey» «755884 / MZB»; 2 ej. «Cala Fornells, Rosas» «Prov. Gerona, Dr. Zariquiey» «75-5885 / MZB»; 2 ej. «Valls (Tarragona), col. Español» «75-5886 / MZB»; 4 ej. «VIII-32, Barcelona, S. Gervasi, en un palomar, Montrós leg.» «75-5900 / MZB»; 2 ej. «VIII-32, Barcelona, S. Gervasi, en un palomar, Montrós leg.» «75-5904 / MZB»; 4 ej. «Molinicos, 15-V-1938 (MNCN)»; 1 ej. «Molinicos, 14-V-1938 (MNCN)»; 1 ej. «Molinicos, 26-VI-1938 (MNCN)»; 2 ej. «Molinicos, 23-X1938 (MNCN)»; 1 ej. «7-VIII-69, Cv. de l’avenc Traiguera Cs, J. Castell leg.» «75-5907 / MZB»; 6 ej. «Parque Nacional de la, Caldera de Taburiente, La Palma (I. Canarias), EspañaSpain» «La Palma, Casas Taburiente, 31/8/99, Rafel G. B.»; 3 ej. «Cova dels, muricets» «Lérida, R-2 31-VIII-10 (MNCN». Depositados en las colecciones del Museu de Ciències Naturals de Barcelona y Museo Nacional de Ciencias Naturales de Madrid.
Longitud de 1,6 a 2,2 mm. Especie anoftálmica y áptera, con el cuerpo de contorno paralelo, cilíndrico, y de color testáceo rojizo. Antenas de once artejos con maza terminal de tres (Fig. 1); mandíbulas bidentadas; palpos labiales con el tercer artejo más grande que el segundo. Protórax tan ancho como largo, convexo, de márgenes laterales paralelos y la superficie con la puntuación igual a la de la cabeza. Escutelo no visible. Élitros convexos, con los ángulos humerales salientes, provistos de puntos poco marcados sin conformar estrías. Tarsos de cuatro artejos. Edeago según figura 3.
Dajoz (1969) describió A. brunneus chilensis con siete ejemplares procedentes Chile (provincia de Curicó) basándose en las diferencias obtenidas en las mediciones en comparación a diez ejemplares franceses. Dichas diferencias las hemos observado en gran parte en los ejemplares estudiados,
93 Butlletí ICHN 85 (3), 2021
FLORA ET FAUNA
FAMILIA SALPINGIDAE
1815 EN EL ÁREA IBEROBALEAR
GEA,
REVISIÓN DE LA
LEACH,
Figura 9. Habitus de Aglenus brunneus (Gyllenhal, 1813).
GEA, FLORA ET FAUNA
por lo que A. brunneus chilensis debe pasar a n. syn. de A. brunneus
Distribución
Pollock & Löbl (2008) dan a la especie una amplia distribución citándola de las regiones Afrotropical, Neártica y Paleártica, indicando su presencia en esta última en: Alemania, Austria, Croacia, Dinamarca, Egipto, Eslovaquia, España, Francia, Gran Bretaña, Grecia, Holanda, Hungría, Irlanda, Islas Azores, Islas Canarias, Islas Madeira, Italia, Líbano, Polonia, Portugal, República Checa, Rusia, Suecia, Suiza. Dajoz (1977) considera la especie subcosmopolita.
La distribución ibérica conocida incluye las siguientes localidades (Fig, 10):
España: Molinicos (Albacete), Beniarbeig, La Xara, Pego (Alicante), sin indicación (Badajoz), Barcelona ciudad, barrios de Sant Gervasi y Sarrià (Barcelona), Villamartín (Cádiz), Traiguera (Castelló), Pozuelo de Calatrava (Ciudad Real), Baena, Hornachuelos (Córdoba), Roses (Girona), Santisteban del Puerto, Villacarrillo (Jaén), Llimiana (Lleida), Totana (Murcia), Sierra de Aralar (Gipuzkoa), Caparroso (Navarra), Valls (Tarragona), Alzira, Llombai (Valencia) (Alonso Román & Bahillo de la Puebla, 2019; Ferrer & Quetglas, 2004; Fuente, 1928; Lencina et al., 2008; Otero et al., 2012; Pérez Fernández, 2007).
Portugal: Coimbra (Fuente, 1928).
Islas Baleares: Portopí (Mallorca) (Fuente, 1928), Menorca (Español, 1955).
10.
Biología
Tal como indican Barnouin & Zagatti (2017) la especie se puede considerar micetófaga endógena, aunque se desconoce su biología. Tiene propensión a la antropofília, se ha citado del estiércol de establos y apriscos, del guano de gallineros y palomares, de murciélagos en cuevas, en cultivos de hongos y en diferentes materias en descomposición (Barnouin, 2014; Barnouin & Zagatti, 2017; Dajoz, 1977). También fue localizada en el cadáver de un Myotis myotis (Borkhausen, 1797) (Ferrer & Quetglas, 2004). Dajoz (1977) la señala de los ni-
dos de Microtus arvalis Pallas, 1778 y Apodemus flavicollis (Melchior, 1834). En Tenerife (Islas Canarias) se obtuvieron un gran número de ejemplares con el tamizado de materia en descomposición.
Subfamila Salpinginae Leach, 1815
Clave de géneros
1 Cabeza más ancha que larga, sin rostro (Figs. 5-7) .......2
– Cabeza más larga que ancha, con rostro (Fig. 8)........... 4
2 Protórax con los márgenes laterales denticulados Lissodema – Protórax con los márgenes laterales no denticulados ....3
3 Epístoma en curva regular, nada saliente del contorno de la cabeza (Fig. 6) ........................................ Sphaeriestes – Epístoma saliente del contorno de la cabeza (Fig. 7)
Rabdocerus
4 Rostro largo o muy largo, siempre más largo que ancho; cuerpo mono o bicolor pero con la cabeza negruzca y el rostro rojizo (solo en S. aeneus es negro); intervalos elitrales impares con puntos en toda su longitud
........................................................................Salpingus
– Rostro corto y muy ancho; cuerpo bicolor, cabeza y protórax del mismo color; intervalos elitrales impares sólo con unos pocos puntos en la zona apical Vincenzellus
Género Lissodema Curtis, 1833
Lissodema Curtis, 1833. Entom. Mag., 1: 187
Stenolissodema Desbrochers des Loges, 1900. Frelon, 8: 6
Apinolissodema Pic, 1919. Mél. Exot.-Entom., 30: 3
Especie tipo: Lissodema heyana Curtis, 1833 (= Salpingus cursor Gyllenhal, 1813)
Clave de especies
1 Protórax igual o más largo que ancho (Fig. 11); antenas diferenciando maza terminal de tres artejos; márgenes laterales protonales no explanados y provistos de pequeños dientes en los dos tercios basales
L. denticolle (Gyllenhal) – Protórax transverso (Fig. 13); antenas con los cuatro últimos artejos engrosados, conformando una maza mal definida; márgenes laterales protonales explanados y denticulados........................................ L. lituratum (A. Costa)
En la separación específica del género se utiliza a menudo la coloración del cuerpo y la configuración elitral, caracteres poco válidos ya que junto a ejemplares de coloración típica, se observa una gran variabilidad en el número de manchas elitrales, hasta su desaparición total y también en su tonalidad más clara u oscura (Viñolas et al., 2018).
Lissodema denticolle (Gyllenhal, 1813) (Fig. 11)
Salpingus denticollis Gyllenhall, 1813. Ins. Suec., 1 (3): 715 Curculio quadripustulatus Marsham, 1802. Ent. brit., 1, Coleopt.: 297
94 Butlletí ICHN 85 (3), 2021
Figura
Mapa con la distribución de Aglenus brunneus
A. VIÑOLAS ET AL.
REVISIÓN DE LA FAMILIA SALPINGIDAE LEACH, 1815 EN EL ÁREA IBEROBALEAR

Rhinosimus quadriguttatum Le Peletier de Saint-Fargeau & Audinet-Serville, 1825. In: Encycl. méth. Hist. nat. Entomol.: 288
Rhinosimus quadrimaculatum Mulsant, 1830. Letres à Julie, 2: 302
Salpingus humerale A. Villa & G. B. Villa, 1833. Col. Europae: 35
Lissodema quadripustulatum var. gallicum Pic, 1933. Echange, 49: 2
Material estudiado
29 ej., etiquetados: 1 ej. «VI-2007, Parque Natural del Seño-río de Bértiz, Navarra, robledal de Q. robur, Recalde & San Martín leg.»; 1 ej. «21-VIII-2008, DG5625, el Vimeners, Riells i Viabrea, Girona, A. Viñolas, J. Muñoz & J. Soler leg.»; 1 ej. «19-VI-2010, DG5625, el Vimeners, Riells i Viabrea, Girona, A. Viñolas, J. Muñoz & J. Soler leg.»; 5 ej. «2-VII-2011, la Maquina Vella, 410 m, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.»; 1 ej. « 21-VII-2011, Puig de Conillers, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.»; 1 ej. «16/25-VI-2012, el Tillar, Vimbodí i Poblet, Tarragona, F. Benvenuti leg.»; 1 ej. «25-VI/4-VII-2012,
el Tillar, Vimbodí i Poblet, Tarragona, F. Benvenuti leg.»; 2 ej. «17-VII-2014, Prat Terrer, Gavar-rós, Guardiola de Berguedà, Barcelona, J. Muñoz & J. Soler leg.»; 1 ej. «31-VII2014, Baga de Gresolet, Gresolet, Saldes, Berguedà, Barcelona, 1430 m, J. Muñoz & J. Soler leg.»; 1 ej. «18-IV-2015, Font dels Tres Reis, EG1257, Torroella de Montgrí, Girona, A. Viñolas leg.»; 1 ej. «11-VI-2015, barranc de Castellfollit, Vimbodí i Poblet, Tarragona, E. Piera leg.»; 3 ej. «26-VI2015, barranc de Castellfollit, Vimbodí i Poblet, Tarragona, E. Piera leg.»; 1 ej. «04-VII-2015, Còrrec del Mas Reguinell, les Dunes, Torroella de Montgrí, Girona, J. Muñoz & J. Soler leg.»; 4 ej. «VI-2016, Valdorva, Navarra, Recalde & San Martín leg.»; 1 ej. «13-VII-2016, les Cent Fonts, l’Espluga de Francolí, Tarragona, E. Piera leg»; 2 ej. «16-VI-2017, Turó del Vilar, Blanes, Girona, P. Pons & E. Lucha leg.»; 1 ej. «16-VI-2018, Turó del Vilar, Blanes, Girona, P. Pons & E. Lucha leg.»; 1 ej. «30-VI-2018, Sot del Grèvol, Montseny, Fogars de Montclús, Barcelona, E. Feliubadaló leg.». Depositados en las colecciones de J. Muñoz, J. I. Recalde, J. Soler y A. Viñolas.
Material revisado
2 ej., etiquetados: 1 ej. «Montseny, 29-VII-42, Español leg.» «Lissodema quadripustulatum Mars., Español det.» «Lissodema lituratum (Costa, 1847), I. Pérez det. 2005» «Lissodema denticolle (Gyllenhal, 1813), A. Viñolas det. 2017» «838135. MZB»; 1 ej. «Planoles (GI), Rio el Rigard 1096 m, B42º19’09,3” E2º05’17,2”, 23-VI-2010, JM. Diéguez leg.» «Lissodema lituratum (A. Costa 1847), JM. Diéguez det. 2010» «Lissodema denticolle (Gyllenhal, 1813), A. Viñolas det. 2017» «2011-0027, MZB». Depositados en la colección del Museu de Ciències Naturals de Barcelona.
Longitud de 1,8 a 3,3 mm. Cuerpo convexo, brillante, testáceo rojizo oscuro, élitros más oscuros con cuatro manchas testáceas claras, muy variables en tamaño por fusión o ausencia, patas y los ocho primeros artejos de las antenas testáceos, la maza antenal oscura. Antenas de once artejos, con maza terminal bien definida de tres antenómeros; mandíbulas con cuatro dientes medio basales. Protórax convexo, ligeramente transverso, 1,05 veces más ancho que largo, con los márgenes laterales curvados regularmente en los dos tercios apicales y en línea entrante en el basal, con pequeños dientes setígeros en número de tres a cinco, irregularmente dispuestos; superficie con el punteado fuerte y denso, más hacia los márgenes laterales. Élitros convexos, de contorno subparalelo, largos, 1,9 veces más largos que anchos tomados conjuntamente, con su máxima anchura en el medio, húmeros redondeados, con estrías de puntos ligeramente embrolladas en la zona discal.
Distribución
Especie de amplia distribución europea (Pollock & Löbl, 2008; Barnouin & Zagatti, 2017) citada de: Alemania, Armenia, Austria, Bélgica, Bosnia, Bulgaria, Croacia, Dinamarca, Eslovaquia, España, Francia, Gran Bretaña, Holanda, Hungría, Irlanda, Italia, Polonia, República Checa, Rumania, Rusia europea, Suecia, Suiza y Cáucaso.
95 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
Figura 11. Habitus de Lissodema denticolle (Gyllenhal, 1813).
GEA, FLORA ET FAUNA
En el área peninsular sólo se conoce de España, citada de: Guardiola de Berguedà, Montseny, Saldes (Barcelona), Blanes, la Jonquera, Planoles, Riells i Viabrea, Torroella de Montgrí (Girona), Parque Natural del Señorío de Bértiz, Valdorva (Navarra), l’Espluga de Francolí, Vimbodí i Poblet (Tarragona) (Piera et al., 2017; Recalde Irurzun & San Martín, 2015; Viñolas et al., 2012, 2013, 2015, 2018) (Fig. 12).
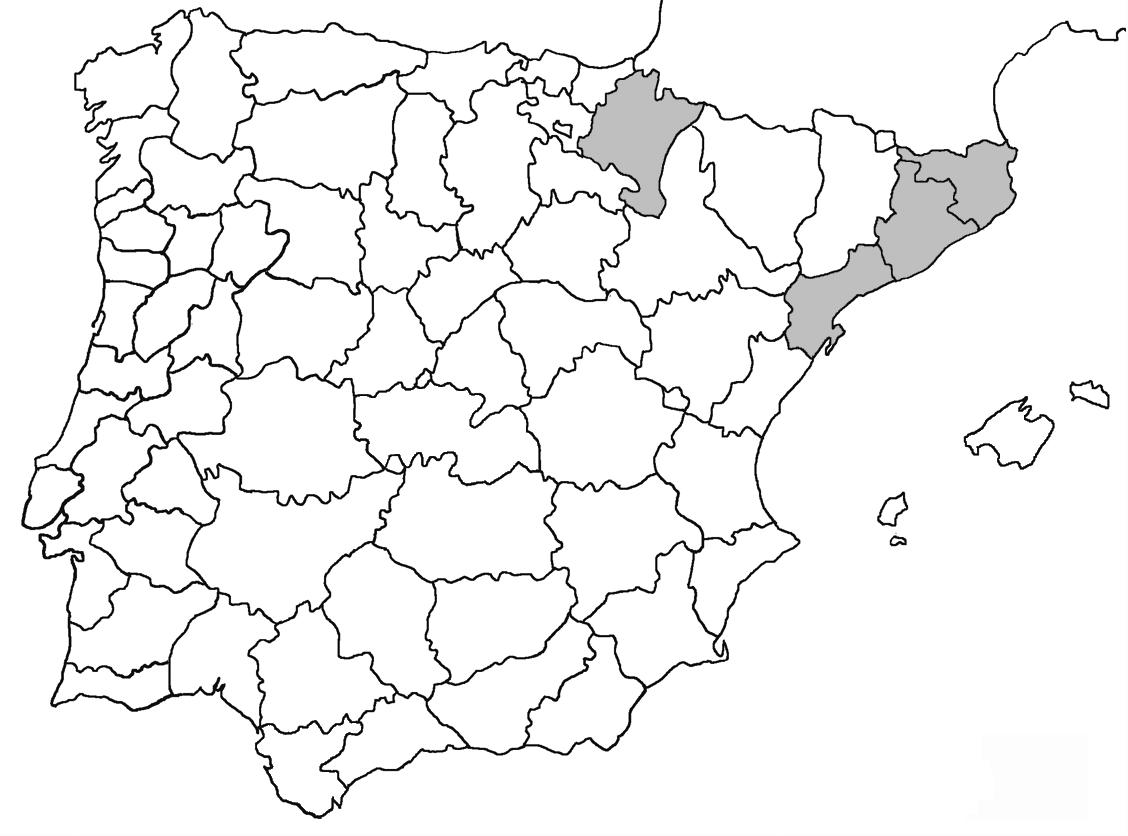
Biología
La especie se localiza en rodales de caducifolios, bajo la corteza de las ramas muertas. Al parecer tanto la larva como el adulto se alimentan de las larvas de Scolytinae, aunque su biología es poco conocida (Barnouin, 2014; Barnouin & Zagatti, 2017).
Lissodema lituratum (A. Costa, 1847) (Fig. 13)
Salpingus lituratus A. Costa, 1847. Ann. Accad. Asp. Nat. Napoli, (2) 1: 158
Lissodema cloueti Guillebeau, 1896. Bull. Soc. Ent. Fr.: 245
Material estudiado
8 ej., etiquetados: 1 ej. «1/30-IV-2011, Turó de Can Balasc, 31TDF2305, 297 m, Vallvidrera, Barcelona, J. Mederos leg.»; 1 ej. «1/28-XII-2018, Turó de Can Balasc, 31TDF2305, 297 m, Vallvidrera, Barcelona, J. Mederos leg.»; 6 ej. «21-II2021, Jou Vell, Peralada, Girona, J. Soler leg.«, Depositados en las colecciones de A. Viñolas, J. Muñoz y J. Soler.
Material revisado
2 ej., etiquetados: 1 ej. «MZB, Montserrat, 2-VII-09, Codina» «MZB, Lissodema lituratum» «Lissodema lituratum Costa, 1847), I. Pérez det. 2003» «83-7962, MZB»; 1 ej. «Montserrat, 2.VII.09, A. Codina» «Lissodema lituratum» «71-2233, MZB». Depositados en la colección del Museu de Ciències Naturals de Barcelona.
Longitud de 1,6 a 2,5 mm. Cuerpo brillante, de color testáceo rojizo oscuro, con zonas más claras o más oscuras, variables, fémures oscuros, tibias testáceas, así como los seis primeros artejos de las antenas. Antenas de once artejos, con

los cuatro últimos ligeramente engrosados; mandíbulas con cuatro dientes medio basales. Protórax convexo, transverso, 1,22 veces más ancho que largo, con la máxima anchura antes del medio; márgenes laterales de contorno curvado y explanado en los tres tercios apicales y fuertemente entrante en línea en el apical, la explanación con tres a cuatro dientes, muy variables en tamaño; superficie con punteado fuerte y denso, y con una línea vertical media lisa. Élitros convexos, 1,62 veces más largos que anchos tomados conjuntamente, con su máxima anchura en el medio; húmeros redondeados; con estrías bien indicadas de gruesos puntos.
Distribución
Pollock & Löbl (2008) la citan de Argelia, Francia, Italia y Túnez, ignorando la antigua cita peninsular de Fuente (1932), reflejada por Barnouin & Zagatti (2017).
En la Península Ibérica sólo se conoce de Barcelona (Montserrat, Vallvidrera) y Girona (Peralada) (Fuente, 1932; Viñolas et al., 2018) (Fig. 14).
Biología
Como la especie anterior las larvas y los adultos viven en la corteza de ramas muertas, se ha señalado de Clematis Dill. ex L, Crataegus Tourn. ex L., Parthenocissus Planch., Populus L., Quercus L., Salix L. Spartium L. y ahora de Ficus carica L. (ex-larva en Peralada). Se tienen pocos datos fiables sobre su biología (Barnouin, 2014; Barnouin & Zagatti, 2017).
96 Butlletí ICHN 85 (3), 2021
Figura 12. Mapa con la distribución de Lissodema denticolle
A. VIÑOLAS ET AL.
Figura 13. Habitus de Lissodema lituratum (A. Costa, 1847).
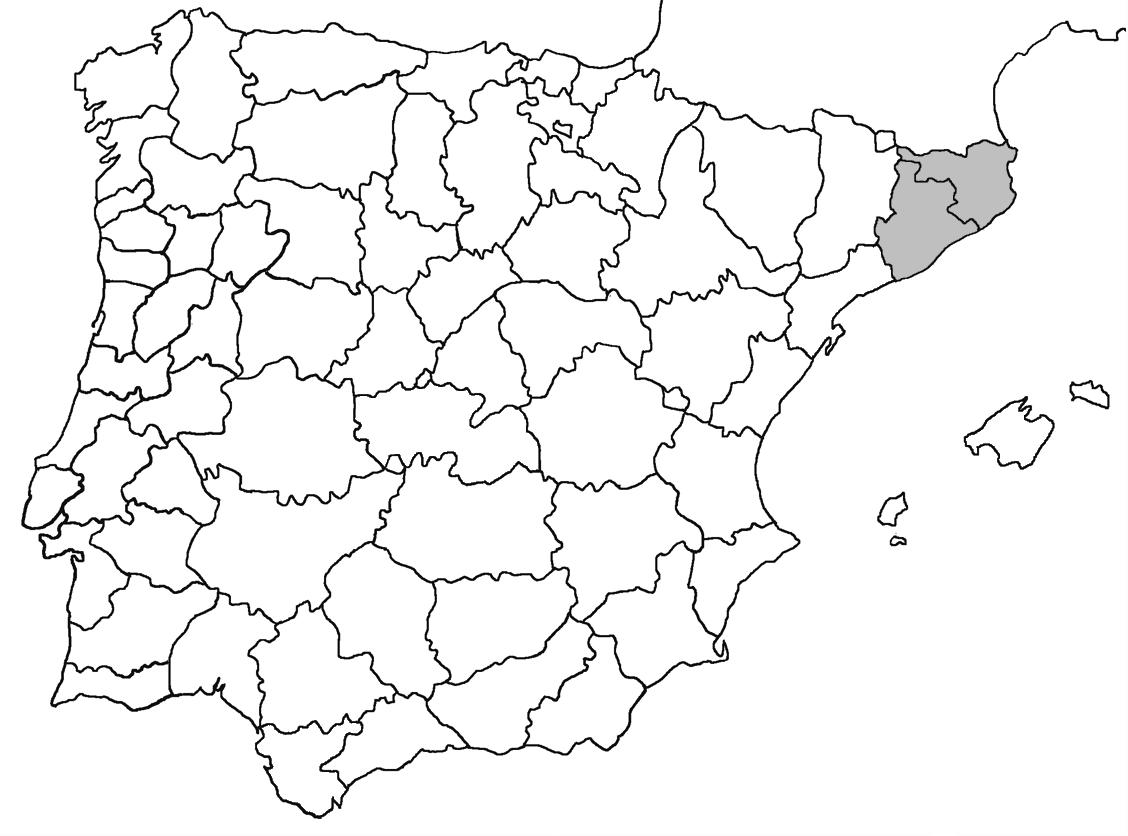
Género Rabocerus Mulsant, 1859
Rabocerus Mulsant, 1859. Hist. nat. col. Fr., Rostriferes: 43
Especie tipo: Salpingus foveolatus Ljungh, 1823.
Con dos especies conocidas del área paleártica, R. foveolatus (Ljungh, 1823) y R. gabrieli Gerhardt, 1901, en la Península Ibérica sólo se ha reportado recientemente la primera.
Rabocerus foveolatus (Ljungh, 1823) (Fig. 15)
Salpingus foveolatus Ljungh, 1823. Sven. Vet. Akad. Nya Handl., (3) 11: 269
Salpingus mutilatus Champion, 1886. Entom. Month. Mag., 23: 160
Rabocerus bishopi Sharp, 1909. Entom. Month. Mag., (2) 20: 245
Rabocerus championi Sharp, 1909. Entom. Month. Mag., (2) 20: 246
Material estudiado
4 ej., etiquetados: 1 ej. «IX-2008, Larra-Roncal, Navarra, Recalde & San Martin leg.»; 1 ej. «V-2013, Etxalar, Navarra, Recalde & San Martin leg.»: 1 ej. «17-IX-2017/1-X-2017, Pleta de Llom, El Serrat, 42°36’52”N 1°34’5”E, 2201,7 m, Flight Intercept S12IHFa67.11237, Andorra, CREAF & J. Bookwalter leg.»; 1 ej. «17-IX-2017/1-X-2017, cerca Parking de Riu de Sorteny, El Serrat, 42°37’14.65”N 1°33’52.21”E, 1963,7 m, Flight Intercept S12ILFb67.11248, Andorra, CREAF & J. Bookwalter leg.». Depositados en las colecciones de J. I. Recalde y J. Bookwalter.
Material revisado
2 ej., etiquetados: 1 ej. «Madrid, Mecklenburg, Fr. W. Konow leg. (MNCN_Ent 220176)»; 1 ej. «Estiria, Oberthur... col. Pérez Arcas (MNCN_Ent 220172)». Depositados en la colección del Museo Nacional de Ciencias Naturales de Madrid.
Longitud de 2,0 a 3,8 mm. Cuerpo convexo de color negro con reflejos bronceados, antenas, epístoma , palpos y patas castaño rojizas muy oscuras. Cabeza con el epístoma saliente

(Fig. 7); antenas de once artejos, del séptimo al onceavo regularmente engrosados, sin formar maza antenal; superficie densamente punteada. Protórax ligeramente convexo, transverso, 1,25 veces más ancho que largo; los márgenes laterales de contorno curvado en la mitad apical y en línea entrante en la basal; con dos impresiones basales nítidas y dos externas muy pequeñas poco o nada indicadas; superficie con denso y fuerte punteado. Élitros de contorno ligeramente oblongo; largos, 1,73 veces más largos que anchos tomados conjuntamente; húmeros redondeados y saliente hacia delante; superficie deprimida en la zona discal basal; estrías de puntos, confusas en la zona discal.
Distribución
De amplia distribución europea citada por Pollock & Löbl (2008) de: Alemania, Austria, Bélgica, Bielorrusia, Dinamarca, Eslovaquia, Estonia, Finlandia, Francia, Gran Bretaña, Hungría, Irlanda, Italia, Letonia, Lituania, Noruega, Polonia, República Checa, Rumania, Rusia, Suecia y Suiza. Ignorando la cita de España (La Rioja) de Pérez Moreno (2005).
La especie, hasta el momento, solo era conocida de dos localidades españolas: Lumbreras (La Rioja), Parque Natural del Señorío de Bertiz (Navarra) (Pérez Moreno, 2005; Pérez
97 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
Figura 14. Mapa con la distribución de Lissodema lituratum
Figura 15. Habitus de Rabocerus foveolatus (Ljungh, 1823).
REVISIÓN DE LA FAMILIA SALPINGIDAE LEACH, 1815 EN EL ÁREA IBEROBALEAR
GEA, FLORA ET FAUNA
Moreno & Moreno Grijalba, 2009; Recalde Irurzun & San Martín, 2015). También, al parecer de Madrid, con el ejemplar de dudosa procedencia depositado en la colección del Museo Nacional de Ciencias Naturales de Madrid (Fig. 16).
Los dos ejemplares de El Serrat son la primera cita de la especie para Andorra. Las localizaciones presentes permiten ampliar las dos únicas localidades ibéricas conocidas. Es clara su presencia en la zona pirenaica, mientras que las localidades del Sistema Ibérico sugiere la existencia de poblaciones relictas en este tipo de áreas montañosas del interior.
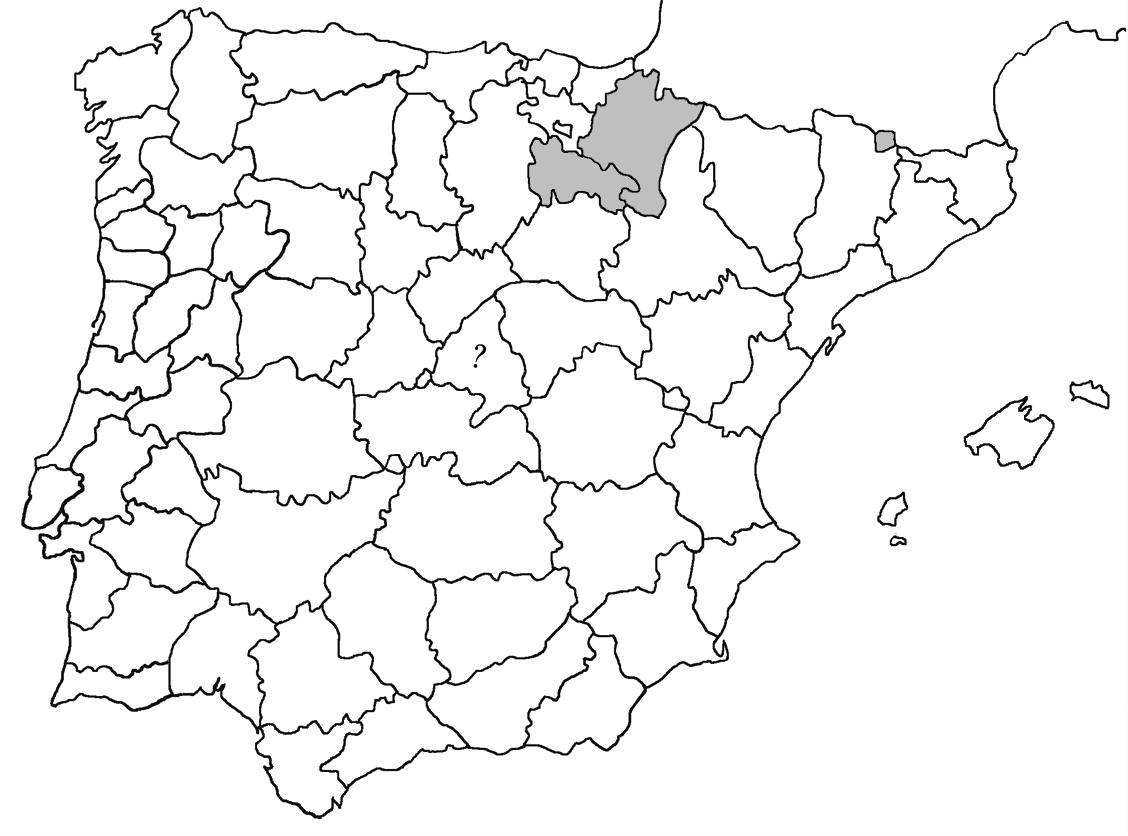
Biología
La especie se desarrolla y localiza debajo de la corteza de árboles muertos atacados por Scolytinae, se ha citado de abedul, arce, haya y roble (Barnouin & Zagatti, 2017). Barnouin (2014) también indica su presencia en resinosas. El ejemplar de Larra (Navarra) se han recolectado en una zona de Fagus sylvatica L. y Pinus uncinata Ramond ex DC y el de Etxalar (Navarra) en un área con F. sylvatica y Quercus robur L.
Género Salpingus Illiger, 1802
Salpingus Illiger, 1802. Mag. Insekt. 1: 150, 301
Anthribus Schellenberg, 1798. Helvet. Entomol.: 120
Rhinosimus Latreille, 1802. Hist. Nat. 3: 192
Rhynosimus A. Costa, 1847. Ann. Accad. Asp. Nat. Napoli, (2) 1: 109 (part. nom. nudum)
Rhinosimus (Cariderus) Mulsant, 1859. Hist. nat. col. Fr., Rostriferes: 46
Especie tipo: Curculio planirostris Fabricius, 1787 (= Curculio fulvirostris Fabricius, 1787)
Clave de especies
1 Rostro corto y ancho (Figs. 17, 19) ...............................2 – Rostro proporcionalmente estrecho y largo (Figs. 21, 23) .................................................................................3
2 Protórax de contorno cordiforme; antenas con maza mal definida de seis artejos S. aeneus
– Protórax con los márgenes de contorno en curva en el tercio apical y en línea entrante hacia la base; antenas con la maza terminal bien definida de cuatro artejos
S. fulvirostris
3 Cuerpo monocolor; antenas con maza terminal mal definida de seis artejos .............................................. S. tapirus – Cuerpo bicolor; antenas con maza terminal mal definida de cinco artejos S. ruficollis

Salpingus aeneus (A. G. Olivier, 1807) (Fig. 17)
Rhinosimus aeneus A. G. Olivier, 1807. Entom., Col., 5 (86): 455
Rhinosimus ornithorynchus Abeille de Perrin, 1874. Bull. Soc. Hist. Nat. Toulouse, 8: 30
Rhinosimus aeneus numidica Pic, 1904. Echange, 20 (229): 10
Cariderus megricus Iablokoff-Kahnzorian, 1956. Dokl. Akad. Nauk Armyansk., 22 (3): 135
Material estudiado
6 ej., etiquetados: 2 ej. «2-IV-2011, la Maquina Vella, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.»; 3 ej. «IV-2014, Iriso, Navarra, Recalde & San Martín leg.»; 1 ej.
98 Butlletí ICHN 85 (3), 2021
Figura 16. Mapa con la distribución de Rabocerus foveolatus
Figura 17. Habitus de Salpingus aeneus (A. G. Olivier, 1807).
A. VIÑOLAS ET AL.
«1-XII-2018, Pinar de Villanueva de Huerva, Zaragoza, E. Piera leg.». Depositados en la colecciones de J. Muñoz, J. Soler, J. I. Recalde.
Material revisado
1 ej., etiquetado: «Catalunya, Balanyà, Vilarubia leg.» «Rhinosimus aeneus Ol., Español det.» «Salpingus aeneus (Olivier, 1807), I. Pérez det. 2005» «83-8130 / MZB». Depositado en la colección del Museu de Ciències Naturals de Barcelona.
Longitud de 2,8 a 4,7 mm. Cuerpo convexo, brillante, de color negro ligeramente marrón con reflejos bronceados más o menos acusados, rostro más claro y rojizo, primeros artejos de las antenas, palpos y patas marrón rojizo, fémures y maza antenal oscurecidos. Cabeza con el rostro corto y ancho; antenas de once artejos, con maza mal definida de seis artejos; superficie fuertemente punteada. Protórax convexo, ligeramente transverso 1,08 veces más ancho que largo; de contorno cordiforme con su máxima anchura en el tercio apical; con las impresiones basales bien indicadas; superficie fuertemente punteada. Élitros convexos, largos, 1,85 más largos que anchos tomados conjuntamente, con la máxima anchura en la parte media; deprimidos en la región discal; estrías de puntos bien indicadas, intervalos impares con una serie de puntos en toda su longitud muy separados.
Distribución
Especie citada por Pollock & Löbl (2008) de: Alemania, Argelia, Austria, Bielorrusia, Francia, Grecia, Hungría, Italia, Letonia, Polonia y Suiza. No indicando su presencia en España aunque ya fue citada por primera vez de Jaén por Molino Olmedo (1997a), localización contemplada por Barnouin & Zagatti (2017), ampliando su distribución a Marruecos.
Actualmente se conoce al menos de las siguientes localidades peninsulares: El Tiemblo (Ávila), Balenyà (Barcelona), Parque Nacional de Cabañeros (Ciudad Real), la Jonquera (Girona), Cantalojas (Guadalajara), Despeñaperros, Venta de los Santos (Jaén), Iriso (Navarra), Campanarios de Azaba (Salamanca), El Espinar (Segovia), Villanueva de Huerva
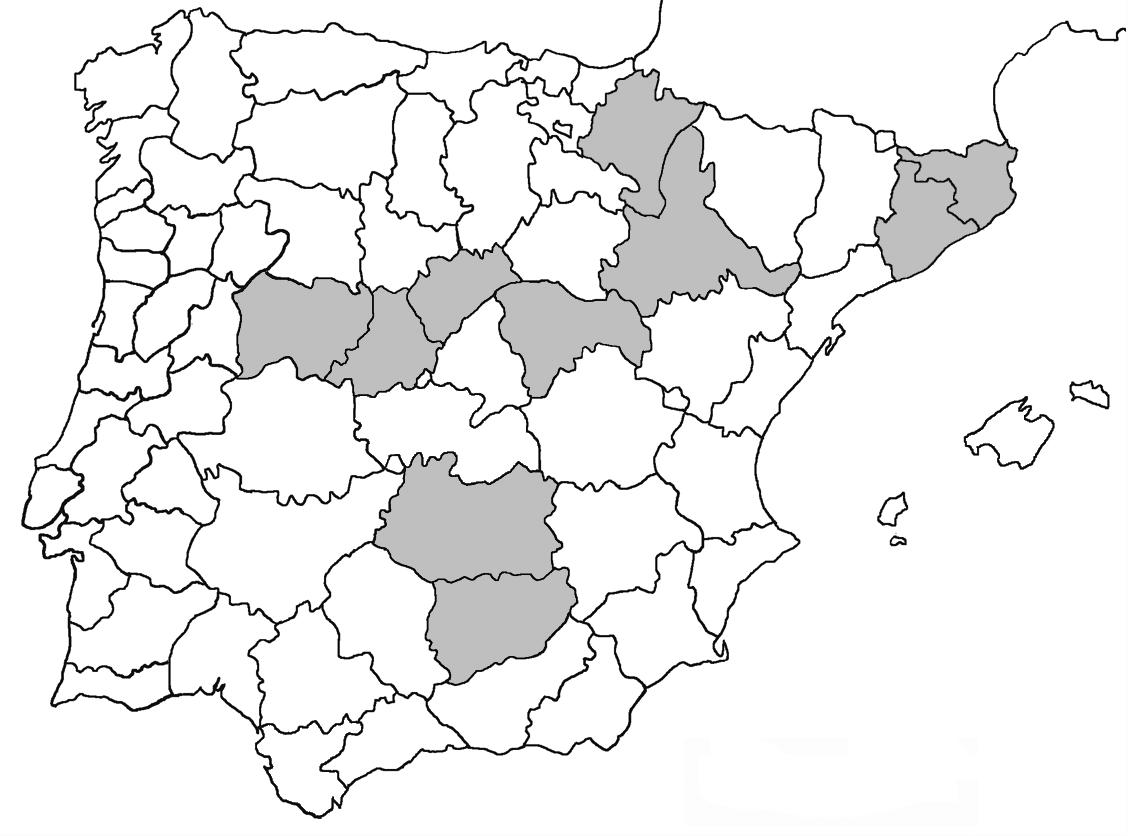
(Zaragoza) (Lencina et al., 2008; Micó et al., 2013; Molino Olmedo, 1997a; Ramírez-Hernández et al., 2014; Viñolas et al., 2019) (Fig. 18).
Biología
La especie se localiza en bosques de caducifolios, preferentemente de haya y roble, también se ha citado en madera descompuesta de quejidos, melojos y encinas (Molino Olmedo, 1997a; Barnouin & Zagatti, 2017). Los ejemplares de Navarra se han recolectado en un quejigal de Quercus faginea Lam. e híbridos.

Salpingus fulvirostris (Fabricius, 1787) (Fig. 19)
Curculio fulvirostris Fabricius, 1787. Mantissa Insect., 2: 381
Curculio planirostris Fabricius, 1787. Mantissa Insect., 1: 119
Rhynosimus spinolae A. Costa, 1847. Ann. Accad. Asp. Nat. Napoli, (2) 1: 109
Salpingus luteonitens Fairmaire, 1879. Rev. Mag. Zool. Pure Appliq., (3) 7: 206
Rhinosimus planirostris var. quadriimpressus Desbrocher des Loges, 1900. Frelon, 8: 22
99 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
Figura 18. Mapa con la distribución de Salpingus aeneus
EL ÁREA IBEROBALEAR
Figura 19. Habitus de Salpingus fulvirostris (Fabricius, 1787).
REVISIÓN DE LA FAMILIA SALPINGIDAE LEACH, 1815 EN
GEA, FLORA ET FAUNA
Material estudiado
96 ej., etiquetados: 2 ej. «03-IV-2008, Valdeinfierno, Los Barrios, Cádiz, A. Verdugo leg.»; 1 ej. «05-IV-2008, Valdeinfierno, Los Barrios, Cádiz, A. Verdugo leg.»; 3 ej. «03-VII-2009, els Vimeners, Riells i Viabrea, Girona, 31TDG5627, A. Viñolas, J. Muñoz & J. Soler leg.»; 1 ej. «25-VII-2009, les Vinyes, Fogars de Montclús, Barcelona, 31TDG5026, A. Viñolas, J. Muñoz & J. Soler leg.»; 1 ej. «01-VIII-2009, coll de Te, Arbúcies, Girona, 31TDG5626, A. Viñolas, J. Muñoz & J. Soler leg.»; 2 ej. «01-VIII-2009, els Vimeners, Riells i Viabrea, Girona, 31TDG5627, A. Viñolas, J. Muñoz & J. Soler leg.»; 3 ej. «29-V-2010, Sant Bernat, Montseny, Barcelona, 31TDG5026, A. Viñolas, J. Muñoz & J. Soler leg.»; 2 ej. «12-VI-2010, Sant Bernat, Montseny, Barcelona, 31TDG5026, A. Viñolas, J. Muñoz & J. Soler leg.»; 1 ej. «18-VII-2010, coll de Te, Arbúcies, Girona, 31TDG5626, A. Viñolas, J. Muñoz & J. Soler leg.»; 4 ej. «2-IV-2011, la Maquina Vella, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.»; 2 ej. «2-VII-2011, Xalamí, La Jonquera, Girona, J. Muñoz & J. Soler leg.»; 2 ej. «31-VII2014. baga de Gresolet, Saldes, Barcelona, J. Muñoz leg.»; 2 ej. «15-VI/25-VII-2012, El Tillar, Vimbodí i Poblet, Tarragona, F. Benvenuti leg.»; 2 ej. «Mas d’en Llac, Garriguella, Girona, J. Soler leg.»; 3 ej. «21-VI-2014, Santa Maria de Requesens, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.»; 1 ej. «31-VII-2014, corral de Rus, Castellar de n’Hug, Barcelona, A. Viñolas leg.»; 1 ej. «31-VII-2014, torrent de les Tortes, Castellar de n’Hug, Barcelona, J. Muñoz leg.»; 28-IX-2017, Pla dels Ocells, Montserrat, Collbató, Barcelona, J. Calaf leg.»; 11 ej. «14-VI-2018, Sot del Grèvol, Montseny, Fogars de Montclús, Barcelona, E. Feliubadaló leg.»; 2 ej. «16-VI-2018, Turó del Vilar, Blanes, Girona, P. Pons & E. Lucha leg.»; 40 ej. «30-VI-2018, Sot del Grèvol, Montseny, Fogars de Montclús, Barcelona, E. Feliubadaló leg.»; 1 ej. «26-V-2021, corral de la pedra detra, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.»; 1 ej. «26-V-2021, font del Ricard, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.»; 3 ej. «26-V-2021, Xalami, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.»; 1 ej. «30-V-2021, els Cavallets, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.»; 3 ej. «30-V-2021, Faig de la pedra, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.»; 2 ej. «30-V-2021, font del faig, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.». Depositados en las colecciones de J. Muñoz, J. Soler y A. Viñolas.
Material revisado
18 ej., etiquetados: 1 ej. «Balenyà (Torrellebreta), A. Vilarubia leg.» «83-7957 / MZB»; 1 ej. «21-VII-15, Lés, Vall d’Arán» «83-7956 / MZB»; 1 ej. «28-VI-47, Montseny, Pla de la Barraca» «83-7959 / MZB»; 2 ej. «VII-69, S. Juan Peña, Huesca, F. Español leg» «71-2473 / 71-2475 / MZB»; 1 ej. «VII-65, Alsina d’Aliña, Lleida, Vilella leg.» «71-2477 / MZB»; 2 ej. «26-IX-69, S. Juan Peña, 1222 m, Huesca, F. Español leg» «71-2472 / 71-2474 / MZB»; 1 ej. «23-XI-73, Sta. Maria de l’Estany, F. Español leg.» «83-8133 / MZB»; 1 ej. «VII/VIII-79, Canfranc (HU), Villalta leg.» «71-2476 / MZB»; 1 ej. «25-VII-87, Stª Fe Montseny, F. Español leg.» «71-2471 / MZB»; 1 ej. «2-VII-1982, Sta Fe del Montseny, Faig, T. Yélamos leg.» «83-7952 / MZB»; 4 ej. «2-VII-1982,
Montseny, faig, T. Yélamos leg.» «83-7954 / MZB»; 2 ej. «4-VII-82, Barcelona, Stª Fe del Montseny (haya), J. L. Moreno & L. Mestres leg.» «83-7955 / MZB». Depositados en la colección del Museu de Ciències Naturals de Barcelona.
Longitud de 2,4 a 3,8 mm. Cuerpo convexo, brillante, de color negro verdoso con reflejos bronceados, rostro, palpos, patas y antenas, excepto la maza más oscura, marrón amarillento. Cabeza con el rostro corto y ancho; antenas de once artejos, con maza terminal de cuatro artejos; superficie irregularmente punteada. Protórax convexo tan ancho como largo, con la máxima anchura en el tercio apical; márgenes de contorno en curva en el tercio apical y en línea entrante hacia la base en los dos tercios restantes; impresiones basales presentes; superficie con el punteado fuerte, irregularmente dispuesto y nada contiguo. Élitros convexos, 1,68 veces más largos que anchos tomados conjuntamente, con la máxima anchura en la parte media; húmeros redondeados; deprimidos en la región discal; estrías de puntos bien indicadas, intervalos impares con una serie de puntos en toda su longitud muy separados.
Comentario
Alonso Zarazaga (2014) indica que Curculio planirostris Fabricius, 1787, es homónimo primario de Curculio planirostris Piller & Mitterpacher, 1783, sinónimo de Tropideres albirostris (Schaller, 1783), dando validez a Curculio fulvirostris Fabricius, 1787, que había sido sinonimizado con C. planirostris al situar ambas especies en el género Anthribus Geoffroy, 1762.
Distribución
Especie conocida de: Alemania, Bélgica, Bielorrusia, Croacia, Dinamarca, España, Estonia, Finlandia, Francia, Gran Bretaña, Holanda, Hungría, Irlanda, Italia, Letonia, Lituania, Marruecos, Noruega, Polonia, Portugal, República Checa, Rumania, Rusia, Suecia y Túnez (Pollock & Löbl, 2008).
En la Península Ibérica se conoce de (Fig. 20): España: Villaverde de Guadalimar (Albacete), Lalastra (Araba), Balenyà, Barcelona, Castellar de n’Hug, Collbató, Fogars de Montclús, l’Estany, Montseny, Saldes (Barcelona), Karrantza Harana (Bizkaia), Algeciras, Los Barrios (Cádiz), Santader (Cantabria), Fuencaliente (Ciudad Real), Añarbe, Arbiun–Erroiarri, Endara, Errenteria, Oiartzum (Gipuzkoa), Arbúcies, Garrigella, la Jonquera, Riells i Viabrera (Girona), Cantalojas, Galve de Sorbe (Guadalajara), Canfranc, San Juan de la Peña (Huesca), Lumbreras, Villoslada de Cameros (La Rioja), l’Alzina d’Alinyà, Lés (Lleida), Somosierra (Madrid), Moratalla (Murcia), Garde, Irañeta, Parque Natural del Señorío de Bertiz (Navarra), Moscoso (Pontevedra), Riofrío de Riaza (Segovia), Vimbodí i Poblet (Tarragona), Moncayo (Zaragoza) (Alonso Román & Bahillo de la Puebla, 2019; Bahillo de la Puebla & Alonso Román, 2018; Champion & Chapman, 1904; Cuní i Martorell & Martorell i Peña, 1876; Diéguez Fernández, 2015; Fuente, 1932; Molino Olmedo, 1997b; Outerelo & Gamarra, 2011; Pagola Carte et al., 2007; Lencina et al., 2008; Pérez Moreno, 2005; Pé-
100 Butlletí ICHN 85 (3), 2021
A. VIÑOLAS ET AL.
rez Moreno & Moreno Grijalba, 2009; Piera et al., 2017; Recalde Irurzun & San Martín, 2015, 2016; Recalde Irurzun et al., 2017; Viñolas et al., 2013, 2014, 2015).
Portugal: Buçaco (Aveiro), Castro de Carvalhelhos (Vila Real) (Fuente, 1932; Grosso Silva, 2002).
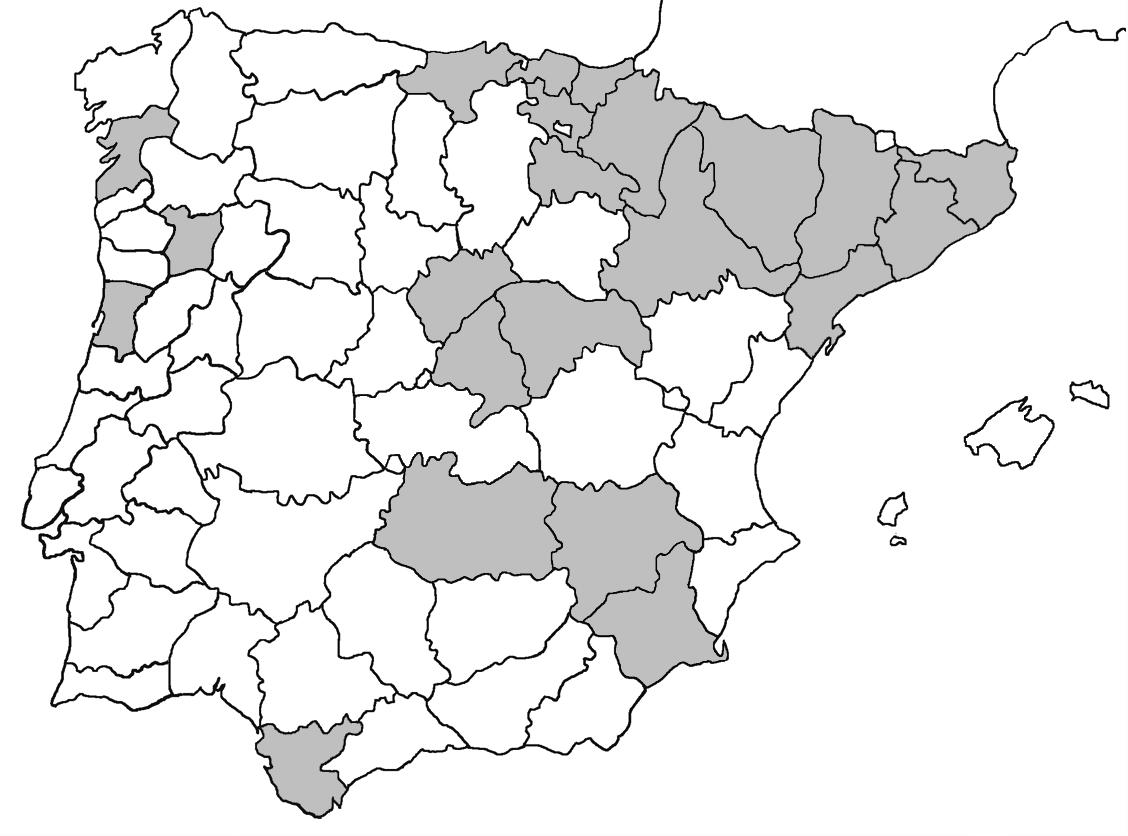
Biología
La especie se localiza en ramas muertas de diferentes grosores de caducifolios, predando Scolytinae y otros pequeños xilófagos (Barnouin & Zagatti, 2017).
Salpingus ruficollis (Linnaeus, 1760) (Fig. 21)
Curculio ruficollis Linnaeus, 1760. Fauna Suecica: 177
Curculio rostratus DeGeer, 1775. Mém. hist. Insect., 5: 252
Curculio roboris Fabricius, 1787. Mantissa Insect., 1: 100
Rhinosimus coeruleocephalus Latreille, 1804. Hist. Nat. 11: 28
Rhinosimus ruficollis var. viridicollis Pic, 1892. Echange, 8 (85): 32
Material estudiado
17 ej., etiquetados: 2 ej. «VII-2004, Vera de Bidasoa, Navarra, en trampa de ventana en Quercus robur, Recalde & San Martín leg.»; 1 ej. «13-VIII-2009, coll de Té, Arbúcies, Girona, 31TDG5627, A. Viñolas, J. Muñoz & J. Soler leg.»; 2 ej. «13-VIII-2009, coll de Té, Arbúcies, Girona, 31TDG5627, A. Viñolas, J. Muñoz & J. Soler leg.»; 2 ej. «15-V-2011, Rec de Mirapols, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.»; 1 ej. «16-XII-2011, resclosa de Requesens, 427 m, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.»; 1 ej. «31-VII-2014, baga de Gresolet, Saldes, Barcelona, J. Muñoz leg.»; 3 ej. «14-VI-2018, Sot del Grèvol, 1200 m, Fogars de Montclús, Barcelona, E. Feliubadaló leg.»; 5 ej. «30-VI-2018, Sot del Grèvol, 1200 m, Fogars de Montclús, Barcelona, E. Feliubadaló leg.». Depositados en la colecciones de J. Muñoz, J. I. Recalde y A. Viñolas.
Material revisado
10 ej., etiquetados: 1 ej. «Ordesa, Huesca, 1200 m, C. Bolivar leg.» (MNCN); 2 ej. «Valle de Ordesa» «83-7950 /

MZB»; 2 ej. «La Fajatona, Montseny» «83-7958 / MZB»; 1 ej. «19-VIII-1941, Sant Segimond, Montseny occ., Barcelona, J. Mateu leg.» «83-8131 / MZB»; 2 ej. «VIII-43, Panticosa (Osca), Muntada leg.» «83-7951 / MZB»; 2 ej. «VII1961, Roncesvalles, Navarra, Hispania, M. González leg.» (MNCN); 1 ej. «VII-1969, San Juan de la Peña, Huesca, F. Español leg.» «71-2478 / MZB»; 1 ej. «VII-69, Montseny, S. Segimond, Español leg.» «83-8132 / MZB»; 1 ej. «29-VII83, Vall de Toran, Faig, T. Yélamos leg.» «83-7953 / MZB». Depositados en la colecciones del Museo Nacional Historia Natural de Madrid y en el Museu de Ciències Naturals de Barcelona.
Longitud de 2,8 a 5,2. Cuerpo convexo, brillante, con la cabeza, protórax y extremidades marrón amarillento, la cabeza presenta una zona negra hasta el nivel de los ojos, élitros azules o verdes metalizados. Cabeza con el rostro estrecho y largo; antenas de once artejos con una maza mal definida de cinco artejos; superficie punteada. Protórax convexo, tan largo como ancho, de contorno cordiforme; impresiones basales bien indicadas; superficie con el punteado irregular, menos denso en la zona discal. Élitros convexos, 1,67 veces más largos que anchos tomados conjuntamente, con su máxima anchura en la parte media; húmeros redondeados; estrías de finos puntos muy bien definidas, intervalos impares con una serie de puntos en toda su longitud muy separados.
101 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
Figura 20. Mapa con la distribución de Salpingus fulvirostris
FAMILIA SALPINGIDAE LEACH, 1815 EN EL ÁREA IBEROBALEAR
Figura 21. Habitus de Salpingus ruficollis (Linnaeus, 1760).
REVISIÓN DE LA
GEA, FLORA ET FAUNA
Distribución
Especie de amplia distribución europea citada por Pollock & Löbl (2008) de: Alemania, Austria, Bielorrusia, Croacia, Dinamarca, Eslovaquia, Eslovenia, España, Estonia, Finlandia, Francia, Holanda, Hungría, Irlanda, Italia, Letonia, Lituania, Noruega, Polonia, República Checa, Rumania, Rusia, Suecia y Suiza.
En el área peninsular sólo se conoce, de momento, de España, citada de: Montes de Vitoria (Araba), Montseny, Saldes (Barcelona), Oiartzum (Gipuzkoa), Arbúcies, la Jonquera, Viladrau (Girona), Cantalojas (Guadalajara), Panticosa, San Juan de la Peña, Valle de Ordesa (Huesca), Lumbreras, Villoslada de Cameros (La Rioja), Sant Joan de Toran (Lleida), Bera de Bidasoa, Irañeta, Parque Natural del Señorío de Bertiz, Roncesvalles (Navarra), Moncayo (Zaragoza) (Alonso Román & Bahillo de la Puebla, 2019; Champion & Chapman, 1904; Lencina et al., 2008; Pagola Carte et al., 2007; Pérez Moreno, 2005; Pérez Moreno & Moreno Grijalba, 2009; Recalde Irurzun & San Martín, 2015; Recalde Irurzun et al., 2017; Viñolas et al., 2012, 2014) (Fig. 22).
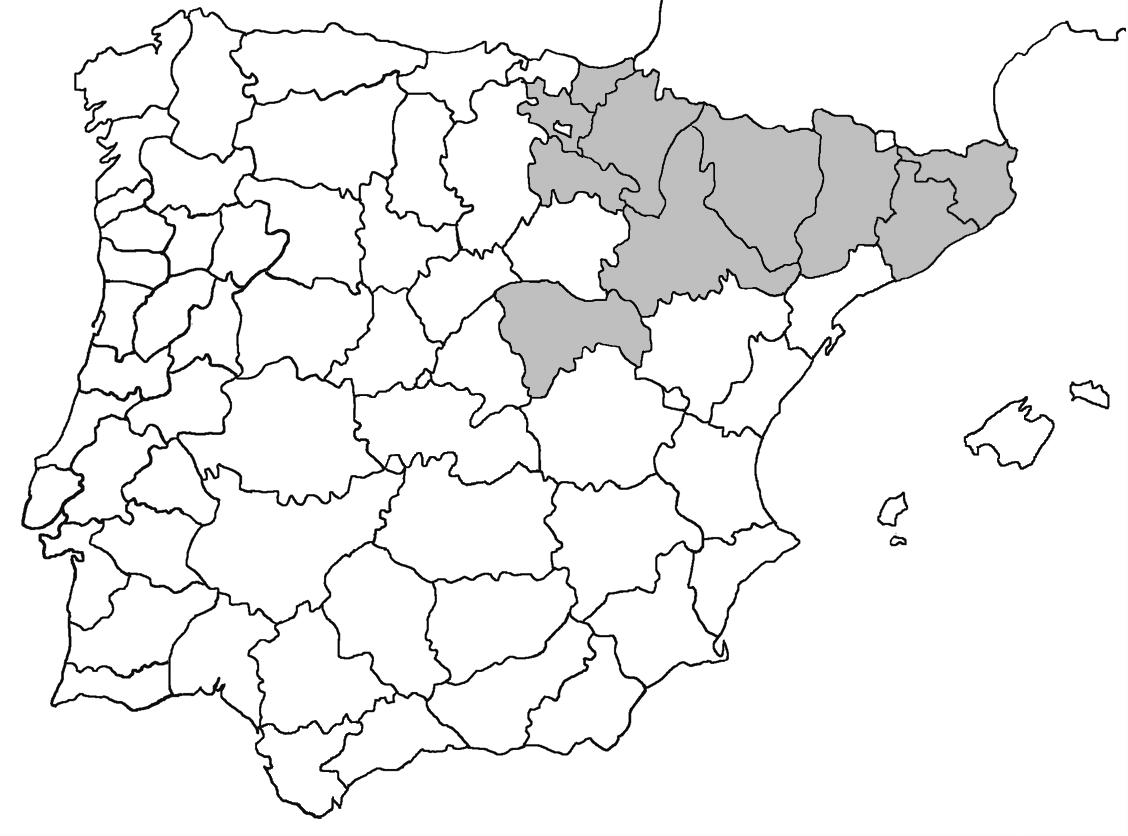
Biología
La especie se localiza en un gran número de caducifolios y resinosas bajo la corteza o en ramas muertas. Al parecer es un predador de pequeños xilófagos (Barnouin & Zagatti, 2017).
Salpingus tapirus (Abeille de Perrin, 1874) (Fig. 23)
Rhinosimus tapirus Abeille de Perrin, 1874. Bull. Soc. Hist.
Nat. Toulouse, 8: 29
Rhinosimus (Cadiredus) angustirostris Pic, 1898. Frelon., 6: 35
Salpingus elephas Chobaut, 1903. Bull. Soc. Ent. Fr.: 143
Rhinosimus (Cadiredus) tapiroides Reitter, 1912. Entom.
Blätter, 8: 196
Material estudiado
29 ej., etiquetados: 2 ej. «15-V-2008, Lesundegi, Olazti, Navarra, Recalde & San Martin leg.»; 14 ej. «IV-2014, Iriso, Navarra, Recalde & San Martin leg.»; 3 ej. «26-V-2021, Xalamí, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.»; 5 ej.

«30-V-2021, Faig de la Pedra, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.»; 2 ej. «30-V-2021, font del Faig, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.»; 1 ej. «30-V-2021, Corral de la Pedra Detra, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.»; 2 ej. «10-VI-2021, Faig de la Pedra, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.». Depositados en las colecciones de J. Muñoz, J. Soler, A. Viñolas y J. I. Recalde Irurzun.
Longitud de 3,1 a 4,7 mm. Cuerpo convexo, brillante, de color marrón negruzco a rojizo, el rostro más claro marrón amarillento, funículo antenal, palpos y patas marrón rojizo muy claro. Cabeza con el rostro largo y estrecho; antenas de once artejos con una maza mal definida de 6 artejos; superficie punteada, con una línea media impunteada en el rostro. Protórax convexo, ligeramente más largo que ancho; de contorno cordiforme, con la máxima anchura en el cuarto apical; impresiones basales bien indicadas; superficie con fuerte e irregular punteado. Élitros convexos, 1,76 veces más largos que anchos tomados conjuntamente; con la máxima anchura en la parte media; húmeros redondeados; deprimidos en la
102 Butlletí ICHN 85 (3), 2021
Figura 22. Mapa con la distribución de Salpingus ruficollis
A. VIÑOLAS ET AL.
Figura 23. Habitus de Salpingus tapirus (Abeille de Perrin, 1874).
REVISIÓN DE LA FAMILIA SALPINGIDAE LEACH, 1815 EN EL ÁREA IBEROBALEAR
región discal; estrías de puntos bien indicadas, intervalos impares con una serie de puntos muy separados.
Distribución
Pollock & Löbl (2008) sólo la citan de: Argelia, Italia y Túnez. Barnouin & Zagatti (2017) amplían su área citándola de España, Francia, Cerdeña y Sicilia.
Especie con muy pocos registros ibéricos, sólo se conoce del Parque Nacional de Cabañeros (Ciudad Real), la Jonquera (Girona), Villoslada de Cameros, Muro en Cameros, Robledal de Turgueiza (La Rioja), Garde, Irañeta, Iriso, Olazti (Navarra), Cantalojas (Guadalajara), Rebollar (Salamanca) (Lencina et al., 2008; Miscó et al., 2013; Ramilo et al., 2017; Recalde Irurzun & San Martín, 2016; Recalde Irurzun et al., 2017; Pérez-Moreno et al., 2020). Los especímenes recolectados en el Paratge Natural d’Interès Nacional de l’Albera, Girona, amplían notablemente el área de distribución de la especie (Fig. 24).
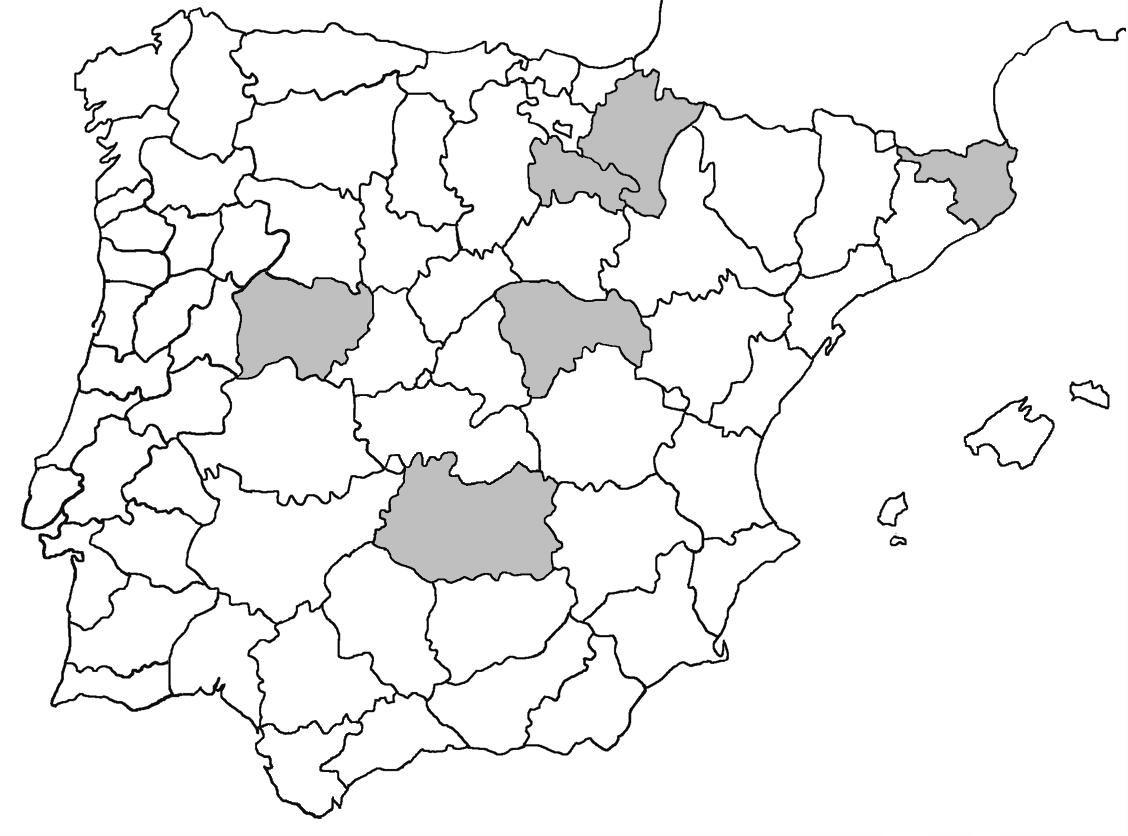
Biología
Preferentemente se localiza en robledales maduros, también se ha recolectado en bosque de ribera y curiosamente en abetos, mientras que los ejemplares de Girona se han recolectado en hayedos mediante polytrap. Los ejemplares de Navarra se han recolectado en un quejigal de Q. faginea e híbridos, y en Quercus petraea (Matt.), Liebl. Se desconoce su biología (Barnouin & Zagatti, 2017).
Género Sphaeriestes Stephens, 1831
Sphaeriestes Stephens, 1831. Illustr. Brit. Entom., 4: 218
Salpingus Gyllenhal, 1810. Ins. Suec., 1 (2): 640
Salpingellus Reitter, 1911. Fauna Germ., 3: 415
Trichocolposinus Seidlitz, 1916. Deutsch. Entom. Zeitsch.: 489
Sphaeriesthes Schenkling, 1922. Coleopt. Catal.: 194
Arabocerus Iablokoff-Khnzorian, 1985. Deutsch. Entom. Zeitsch., 32: 2016
Especie tipo: Dermestes ater Paykull, 1798 (= Salpingus stockmanni Biström, 1977)
Clave de especies
1 Cuerpo negro brillante, ápices del mismo color; maza antenal bien definida de tres artejos; protórax muy densamente punteado S. stockmanni
Cuerpo marrón rojizo o negruzco rojizo, ápices más claros; maza antenal mal definida de cuatro cinco artejos; protórax con punteado bien indicado, pero no muy denso .............................................................................2
2 Epipleuras elitrales largas alcanzando la sutura apical; maza antenal mal definida de cinco artejos S. castaneus
– Epipleuras elitrales cortas no alcanzando la sutura apical e interrumpidas a nivel de las metacoxas; maza antenal mal definida de cuatro artejos ........................................3
3 Cuerpo de color marrón rojizo muy oscuro a negruzco, ápices más claros; antenas con los artejos 8 a 10 más largos que anchos S. reyi – Cuerpo marrón rojizo, ápices más claros; antenas con los artejos 8 a 10 tan anchos o más que largos .... S. exsanguis
Sphaeriestes castaneus (Panzer, 1796) (Fig. 25)
Notoxus castaneus Panzer, 1796. Faun. Ins. Deutsch. Ins., 31: 16
Salpingus achilleae Bonelli, 1812. Mem. R. Soc. Agr., Torino, 9: 175
Salpingus piceae Germar, 1824. Faun. Insect. Europ., 10: 9
Sphaeriestes immaculatus Stephens, 1831. Illustr. Brit. Entom., 4: 218
Salpingus castaneus var. brunnescens Pic, 1892. Echange, 8 (85): 32
Material estudiado
19 ej., etiquetados: 3 ej. «VIII-1997, la Garriga, Barcelona, A. Viñolas leg.»; 1 ej. «09-VIII-2009, la Bedollera, Cerc, Lleida, CG7787, A. Viñolas leg.»; 1 ej. «baga de Gresolet, Saldes, Barcelona, J. Muñoz leg.»; 2 ej. «02-VII-2009, prat Terres, Guardiola de Berguedà, Barcelona, A. Viñolas leg.»; 2 ej. «2-VIII-2009, Sot de Penyacans, Montseny, Fogars de Montclús, Barcelona, A. Viñolas, J. Muñoz & J. Soler leg.»; 2 ej. «15-VI/25-VII-2012, El Tillar, Vimbodí i Poblet, Tarragona, F. Benvenuti leg.»; 1 ej. «2014, Iriso, Navarra, Recalde & San Martín leg.»; 4 ej. «V/VI-2015, Mace, Isaba, Navarra, Recalde & San Martín leg.»; 5 ej. «20-V-2017, Larraza, Isaba, Navarra, Recalde & San Martín leg.»; 1 ej. «8/26-VI2018, Comis Vell, Vallnord Arcalis, Ordino, Andorra, CREAF leg.». Depositados en las colecciones de J. Muñoz, A. Viñolas y J. I. Recalde.
Material revisado
4 ej., etiquetados: 1 ej. «Françe, Royer» «83-7960 / MZB»; 1 ej. «10-XI-1981, Guardiola, Barna, F. Español leg.» «838139 / MZB»; 1 ej. «6-XII-87, Mascà, Ports Tortosa, X. Vázquez leg.» «bajo corteza de Pinus sylvestris» «83-8137 / MZB»; 1 ej. «23-VI-2011, Coll del Cantó (Ll) 106, km 260 (N-260) 1708 m, N42º22’13.7” E1º14’14.3”, JM. Diéguez leg.» «2011-1336 / MZB». Depositados en la colección
Museu de Ciències Naturals de Barcelona.
103 Butlletí ICHN 85 (3), 2021
FLORA ET FAUNA
GEA,
–
del
Figura 24. Mapa con la distribución de Salpingus tapirus
GEA, FLORA ET FAUNA

Longitud de 2,0 a 3,9 mm. Cuerpo convexo, de color marrón rojizo con los apéndices más claros. Cabeza con el clípeo trapezoidal; superficie con punteado irregular y poco denso; antenas de once artejos, con los cinco últimos engrosados. Protórax convexo, ligeramente transverso, 1,19 veces más ancho que largo, con la máxima anchura en el tercio apical; los márgenes de contorno en curva en el tercio apical y el resto en línea entrante hacia la base; impresiones basales poco indicadas, superficie con el mismo punteado que la cabeza. Élitros convexos, 1,68 veces más largos que anchos tomados conjuntamente, con la máxima anchura en la mitad apical; húmeros redondeados; epipleuras largas alcanzando la sutura apical; superficie con el mismo punteado que el resto del cuerpo, irregularmente dispuesto y sin definir estrías. Macho con los protarsos dilatados.
Distribución
Conocida en Europa de: Alemania, Austria, Bélgica, Bosnia, Bielorrusia, Croacia, Dinamarca, Eslovaquia, Eslovenia,
España, Estonia, Finlandia, Francia, Gran Bretaña, Holanda, Hungría, Irlanda, Italia, Letonia, Lituania, Noruega, Polonia, República Checa, Rusia, Suecia y Suiza. Pollock & Löbl, 2008 no la citan de España aunque la primera cita ibérica es de Pérez Moreno (2005), igual sucede con la distribución de Barnouin & Zagatti (2017).
En el área de estudio sólo se conoce de:
Andorra: Ordino.
España: Fogars de Montclús, la Garriga, Guardiola de Berguedà, Saldes (Barcelona), Galve de Sorbe, Siles, Somolinos, Zaorejas (Guadalajara), Alàs i Cerc, Coll del Cantó (Lleida), El Escorial (Madrid), Moratalla, Totana (Murcia), Garde, Iriso, Isaba (Navarra), San Rafael (Segovia), Soria (Soria), Alfara de Carles, Vimbodí i Poblet (Tarragona), Albarracín (Teruel) (Diéguez Férnandez, 2012; Lencina et al., 2008; Pérez Moreno, 2005; Piera et al., 2017; Recalde Irurzun & San Martín, 2016; Viñolas et al., 2013, 2014, 2015) (Fig. 26).
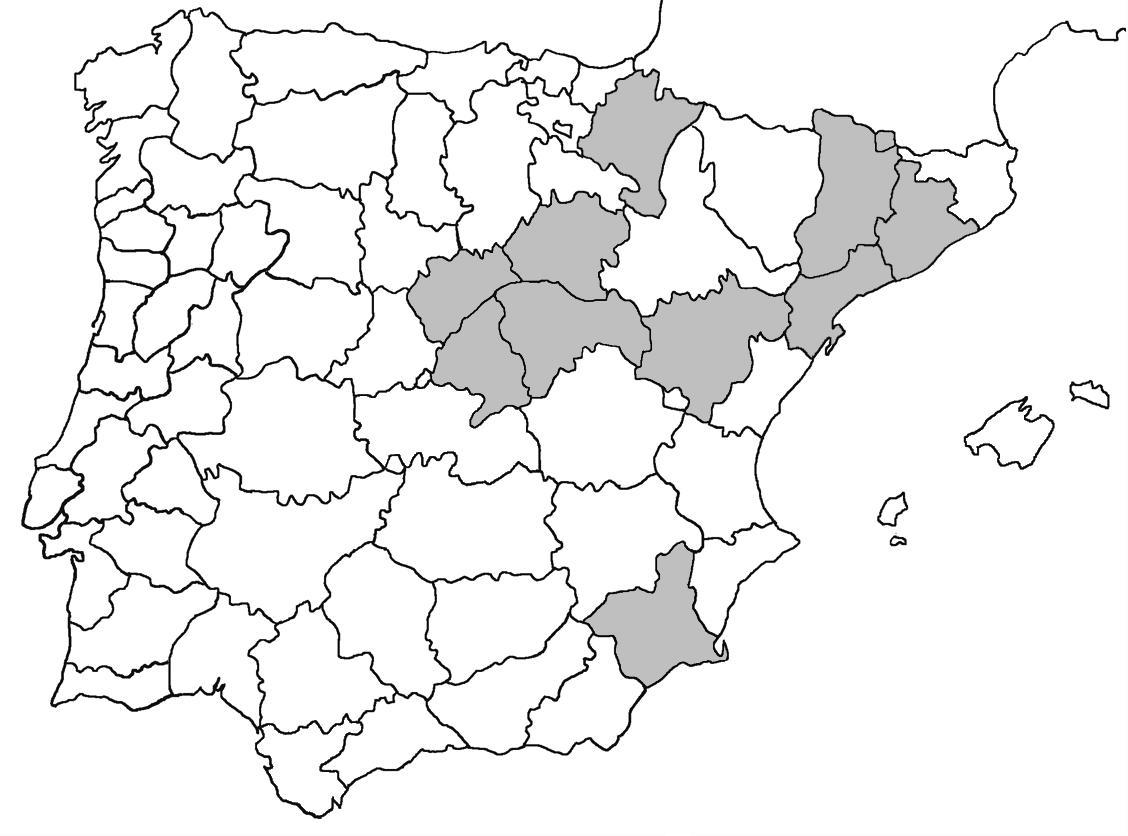
Biología
Especie estrechamente relacionada con las resinosas de los géneros Pinus L. Abies Mill. atacados por Scolytinae, también se localiza en árboles muertos (Barnouin, 2014; Barnouin & Zagatti, 2017). Los ejemplares de Navarra se han recolectado en Pinus sylvestris L. y en quejigal de Q. faginea e híbridos.
Sphaeriestes exsanguis (Abeille de Perrin, 1870) (Fig. 27) Salpingus exsanguis Abeille de Perrin, 1870. Abeille, Mém. Ent., 7: 89
Material estudiado
4 ej., etiquetados: 1 ej. «IX-2010, Rada, Navarra, Recalde & San Martín leg.»; 1 ej. «28-IX-2011, coll del Falcó, 275 m, Portbou, Girona, J. Muñoz & J. Soler leg.»; 1 ej. «V-2012, Mélida, Navarra, Recalde & San Martín leg.»; 1 ej. «VIII2013, Traibuenas, Navarra, Recalde & San Martin leg.». Depositados en las colección de J. Soler y J. I. Recalde.
104 Butlletí ICHN 85 (3), 2021
Figura 25. Habitus de Sphaeriestes castaneus (Panzer, 1796). Figura 26. Mapa con la distribución de Sphaeriestes castaneus
A. VIÑOLAS ET AL.
REVISIÓN DE LA FAMILIA SALPINGIDAE LEACH, 1815 EN EL ÁREA IBEROBALEAR

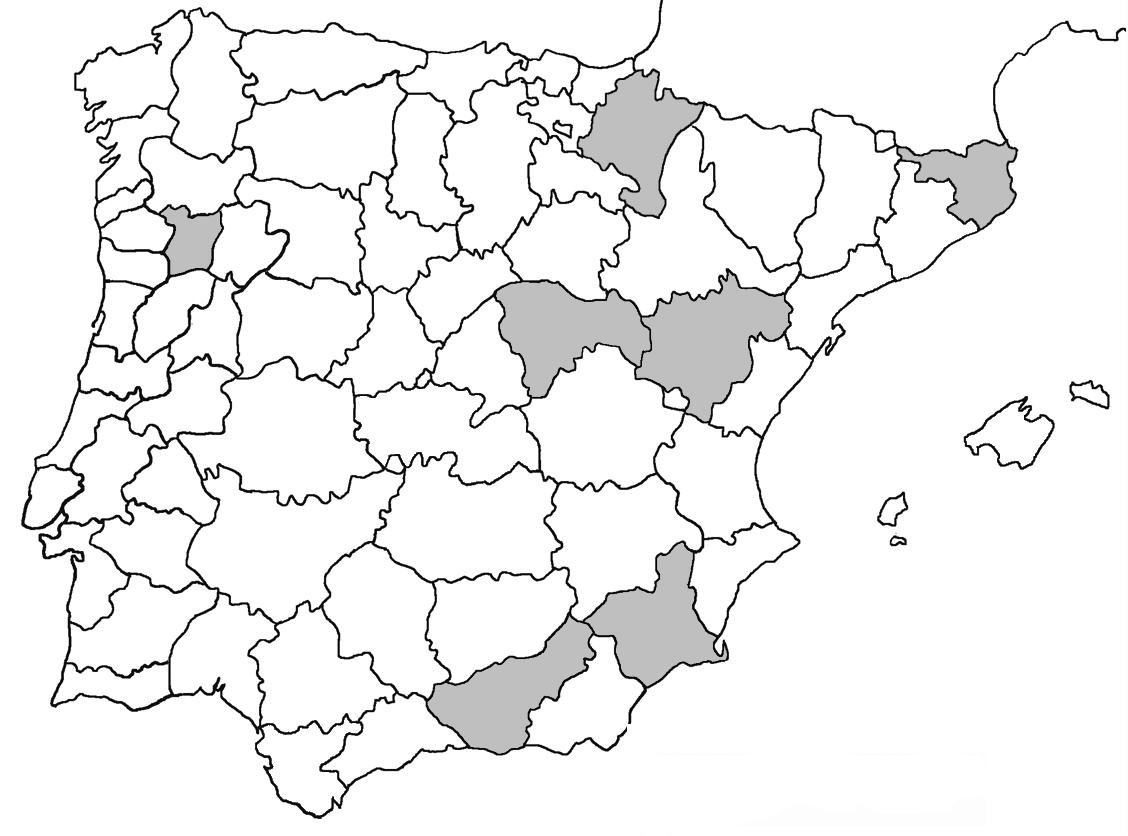
España: Portbou (Girona), Baza (Granada), Villanueva de Alcorón (Guadalajara), Jumilla, Moratalla, Yecla (Murcia), Mélida, Rada, Traibuenas (Navarra), Camarena de la Sierra (Teruel) (Lencina et al., 2008; Viñolas et al., 2012).
Portugal: São Martinho de Antas (Vila Real) (Fuente, 1932).
Biología
La especie se ha citado y relacionado con Pinus halepensis Mill., y Pinus sylvestris. El ejemplar de Girona se ha recolectado sobre Pinus nigra salzmannii (Dunal) Franco y los de Navarra sobre P. halepensis. Se desarrolla bajo la corteza y en las ramas muertas predando Scolytinae (Barnouin, 2014; Barnouin & Zagatti, 2017).
Sphaeriestes reyi (Abeille de Perrin, 1874) (Fig. 29)
Longitud de 2,3 a 2,8 mm. Cuerpo convexo, de color marrón rojizo con las extremidades más claras. Cabeza con el clípeo trapezoidal; antenas de once artejos, con los artejos ocho a diez más anchos que largos, con maza mal definida de cuatro artejos; superficie con el punteado bien indicado y bastante denso. Protórax convexo, ligeramente transverso, 1,15 veces más ancho que largo, con la máxima anchura en la mitad apical; márgenes laterales de contorno en curva entrante hacia la base; impresiones basales bien indicadas; superficie con el mismo punteado que la cabeza. Élitros convexos, 1,76 veces más largos que anchos tomados conjuntamente, con la máxima anchura en la parte media; húmeros redondeados; con estrías bien impresas de gruesos puntos; epipleuras estrechadas progresivamente hasta el nivel de las metacoxas. Machos con los protarsos dilatados.
Distribución
Pollock & Löbl (2008) la citan sólo de Francia, ignorando el antiguo registro de Portugal de Correa de Barros (Fuente, 1932). Por contra Barnouin (2014) y Barnouin & Zagatti (2017) la citan de la Francia meridional, Córcega, Madeira y Península Ibérica.
En la Península Ibérica se ha citado de (Fig. 28):
Salpingus reyi Abeille de Perrin, 1874. Bull. Soc. Hist. Nat. Toulouse, 8: 27
Sphaeriestes ater Stephens, 1831. Illustr. Brit. Entom., 4: 218
Sphaeriestes aeneus Stephens, 1835. In: Illustr. Brit. Entom., 5: 421
Sphaeriestes palpalis Baudi di Selve, 1877. Col. coll. Mus. Zool. Torino, 1: 49
Salpingus tetramerus Rey, 1892. Echange, 8 (95): 138
Sphaeriestes (Salpingellus) reyi var. fowleri Seidlitz, 1916.
Deutsch. Entom. Zeitsch.: 467
Material estudiado
6 ej., etiquetados: 2 ej. «13-X-2017, Can Maçana, Montserrat, el Bruc, Barcelona, J. Calaf leg.»; 2 ej. «20-VIII-2018, Matastacs, Montserrat, el Bruc, Barcelona, J. Calaf leg.»; 2 ej. «14-XI-2018, Sant Pau de la Guàrdia, Montserrat, el Bruc, Barcelona, J. Calaf leg.». Depositados en la colección de J. Muñoz.
Material revisado
21 ej., etiquetados: 1 ej. «21-VII-68, Tiana, Español» «712482 / MZB»; 20 ej. «24-VIII-69, La Conreria, Tiana, Barcelona, roble quemado, F. Español & Dufort leg.» «Salpingus reyi Ab. F. Español det.» «71-2478, 71-2480, 71-2481 /
105 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
Figura 27. Habitus de Sphaeriestes exsanguis (Abeille de Perrin, 1870).
Figura 28. Mapa con la distribución de Sphaeriestes exsanguis.
GEA, FLORA ET FAUNA

MZB». Depositados en la colección del Museu de Ciències Naturals de Barcelona.
Longitud de 2,0 a 3,6 mm. Cuerpo convexo, brillante, de color marrón rojizo muy oscuro a negruzco, artejos del funículo antenal, palpos y patas más claros, los fémures están ligeramente oscurecidos. Cabeza con el clípeo trapezoidal; antenas de once artejos, del octavo al décimo más largos que anchos, los cuatro últimos dilatados formando maza; superficie con punteado fuerte, no contiguo; Protórax convexo, ligeramente transverso 1,09 veces más ancho que largo; márgenes laterales de contorno curvado en el tercio apical y los dos basales ligeramente cóncavos y entrantes hacia la base, con la máxima anchura en el tercio apical; impresiones basales poco indicadas; superficie con el punteado fuerte y denso. Élitros convexos, 1,89 veces más largos que anchos tomados conjuntamente; húmeros redondeados; epipleuras presentes hasta el nivel de metacoxas; superficie con las estrías de gruesos puntos bien indicadas. Macho con los protarsos ligeramente dilatados.
Distribución
Conocida de: Alemania, Chipre, Creta, Dinamarca, España, Finlandia, Francia, Gran Bretaña, Holanda, Irlanda, Italia, Noruega, Polonia, Siria y Suecia (Pollock & Löbl, 2008).
De la Península Ibérica la podemos citar de la Conreria: Tiana, el Bruc (Barcelona), Villarroya (La Rioja), Palaciosmil (León), Rebollar (Salamanca) (Español, 1971; Etxebeste et al., 2013; Pérez Moreno et al., 2020; Ramilo et al., 2017) (Fig. 30).
Fuente (1932) pone en duda la cita de Cuní i Martorell & Martorell i Peña (1876) de Dermestes ater Paykull, 1798 (= S. stockmanni (Biström, 1977)) de los montes cercanos a Barcelona por posible confusión de dichos autores con S. ater Stephens, 1831 (= S. reyi (Abeille de Perrin, 1874)) presente en la zona.
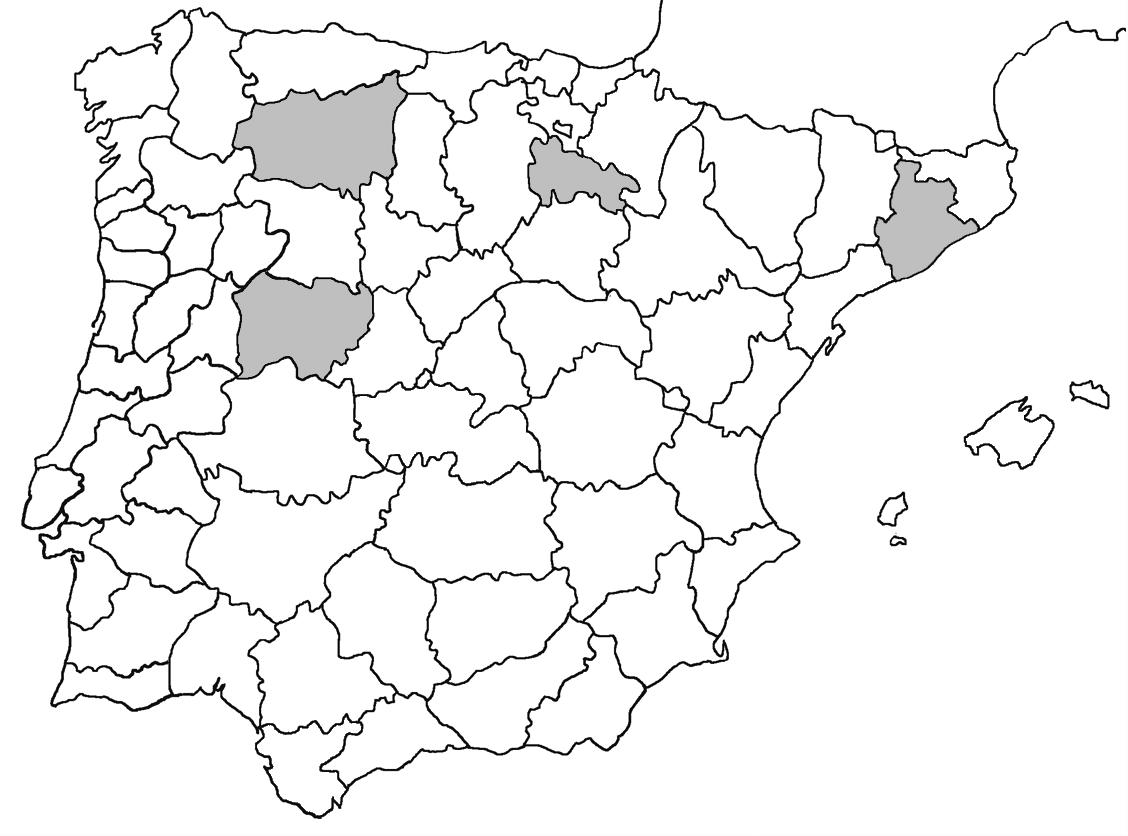
Biología
Se ha citado de abedul, álamo, sauce, pino, roble y diversos frutales. Tiene una especial afinidad con los bosques quemados (Barnouin & Zagatti, 2017). Los ejemplares de Tiana (La Conreria) se capturaron bajo corteza de roble quemado.
Sphaeriestes stockmanni (Biström, 1977) (Fig. 31)
Salpingus stockmanni Biström, 1977. Notulae Entomol., 57: 18
Dermestes ater Paykull, 1798. Faun. Suec., Insect., 1 : 298
Material estudiado
1 ej., etiquetado: «V-2010, Garralda, Navarra, robledal, J. I. Recalde leg.» «Sphaeriestes, stockmanni, Recalde det. 2013». Depositado en la colección de J. I. Recalde.
Longitud de 2,5 a 3,7 mm. Cuerpo convexo, de color negro brillante, funículo antenal, tibias y tarsos ligeramente más claros, algo rojizos. Cabeza con el clípeo trapezoidal; antenas de once artejos, con maza terminal de tres; superficie con punteado denso. Protórax convexo, ligeramente transverso, 1,12 veces más ancho que largo; márgenes de contorno curvado entrante hacia la base y escotados en dicho punto, con su máxima anchura en el tercio apical; fosas basales poco indicadas, en numerosas ocasiones borradas; superficie con el punteado fuerte, irregularmente dispuesto y contiguo
106 Butlletí ICHN 85 (3), 2021
Figura 29. Habitus de Sphaeriestes reyi (Abeille de Perrin, 1874).
Figura 30. Mapa con la distribución de Sphaeriestes reyi
A. VIÑOLAS ET AL.
REVISIÓN DE LA FAMILIA SALPINGIDAE LEACH, 1815 EN EL ÁREA IBEROBALEAR

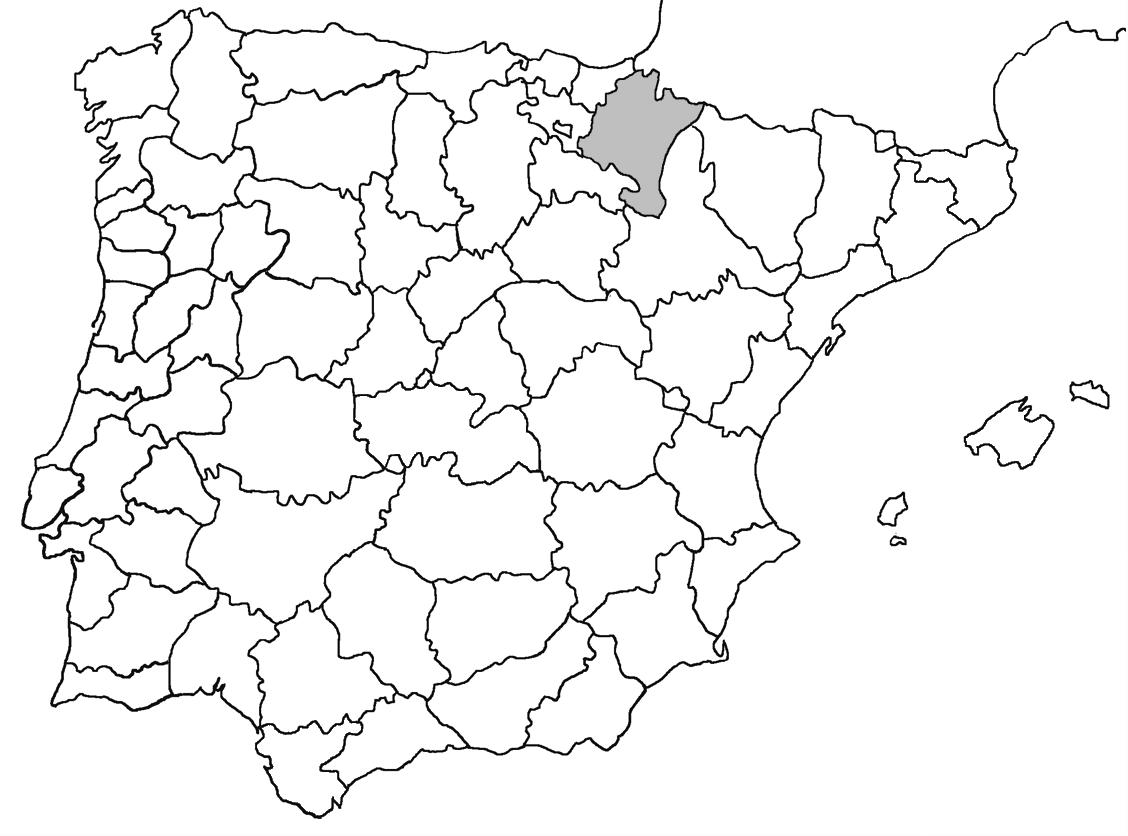
Biología
Se localiza bajo la corteza y en ramas muertas de caducifolios y resinosas, con una clara preferencia por estas últimas. Se ha recolectado en bosques quemados de Pinus y Quercus (Barnouin & Zagatti, 2017).
Género Vincenzellus Reitter, 1911
Vincenzellus Reitter, 1911. Fauna Germanica: 418
Rhynosimus A. Costa, 1847. Ann. Accad. Asp. Nat. Napoli, (2) 1: 107 (part. nom. nudum)
Especie tipo: Rhinosimus viridipennis Latreille, 1804 (= Anthribus ruficollis Panzer, 1794)
Vincenzellus ruficollis (Panzer, 1794) (Fig. 33)
Anthribus ruficollis Panzer, 1794. Faun. Ins. Deutsch. Ins., 24: 19
en diferentes zonas. Élitros convexos, 1,76 veces más largos que anchos tomados conjuntamente, con su máxima anchura en la parte media; húmeros redondeados, nada salientes hacia delante; epipleuras borradas a nivel de las metacoxas; con estrías de puntos bien indicadas. Machos con los protarsos ligeramente dilatados.
Distribución
Con una muy amplia distribución se conoce de: Alemania, Austria, Bielorrusia, Dinamarca, Eslovaquia, Estonia, Finlandia, Francia, Gran Bretaña, Holanda, Hungría, Irlanda, Italia, Letonia, Noruega, Polonia, República Checa, Rusia (europea y asiática), Suecia, Suiza y Ucrania, citada también de la región Neártica (Pollock & Löbl, 2008).
Especie citada recientemente del área peninsular de Garralda (Navarra) (Viñolas et al., 2018) (Fig. 32).
Como se ha mencionado anteriormente, fue citado de los montes próximos a Barcelona por Cuní i Martorell & Martorell i Peña (1876) como Dermestes ater Paykull, 1798, registro cuestionado por Fuente (1932) por probable confusión con S. ater Stephens, 1831 (= S. reyi (Abeille de Perrin, 1874).
Anthribus roboris Fabricius, 1798. Suppl. entom. system.: 161
Rhinosimus viridipennis Latreille, 1804. Hist. Nat. 11: 26
Rhynosimus genei A. Costa, 1847. Ann. Accad. Asp. Nat. Napoli, (2) 1: 107
Rhinosimus ruficeps Bose, 1858. Entom. Zeitung, 19: 96
Material estudiado
4 ej., etiquetados: 1 ej. «V-2002, Lizaso, Navarra, trampa de ventana en Quercus robur, Recalde & San Martín leg.», 2 ej. «15-V-2011, Rec de Mirapolls, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.»; 1 ej. «20-V-2012, les Pipes, la Jonquera, l’Albera, Girona, J. Muñoz & J. Soler leg.». Depositados en las colecciones de J. Muñoz, J. I. Recalde y J. Soler.
Longitud de 2,5 a 4,1 mm. Cuerpo convexo, cabeza y protórax rojizos, élitros azul metálico, patas marrón amarillento y las antenas marrón rojizo con la maza antenal más oscura. Cabeza con el rostro corto y muy ancho; antenas de once artejos con maza terminal, mal defina, de cinco artejos; superficie con el punteado fuerte e irregular. Protórax convexo, no transverso 1,07 veces más largo que ancho; márgenes de contorno cordiforme, con su máxima anchura en el tercio apical;
107 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
Figura 31. Habitus de Sphaeriestes stockmanni (Biström, 1977).
Figura 30. Mapa con la distribución de Sphaeriestes stockmanni
GEA,

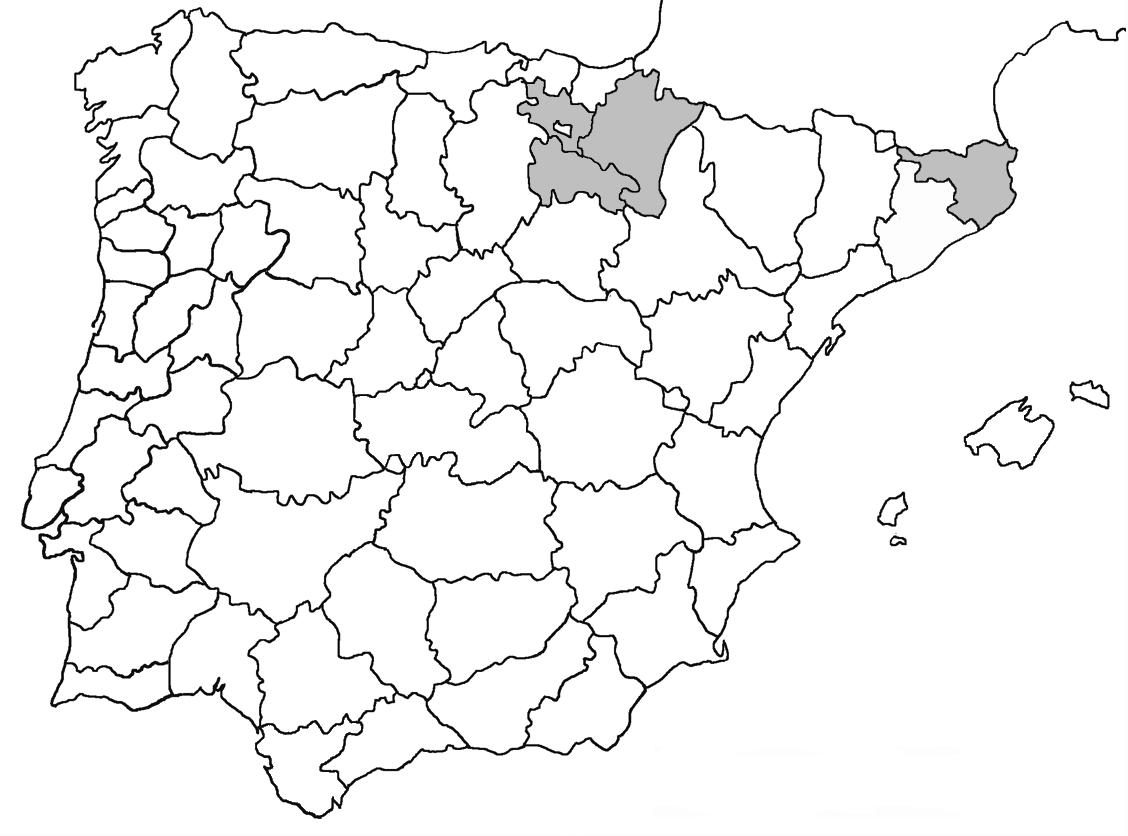
Biología
La especie se localiza en la madera podrida de diferentes especies de caducifolios, tales como hayas, robles y sauces, en donde se alimenta, al parecer, de los hongos presentes en la descomposición de la misma (Barnouin & Zagatti, 2017).
Agradecimientos
A Glòria Masó y Berta Caballero, conservadoras del Museu de Ciències Naturals de Barcelona, las facilidades ofrecidas para la revisión de los ejemplares de Salpingidae depositados en la colección de la entidad. A Mercedes París, del Museo Nacional de Ciencias Naturales de Madrid, el habernos permitido revisar los ejemplares de la familia depositados en la colección de la entidad.
impresiones basales fuertemente indicadas; superficie fuerte e irregularmente punteada. Élitros convexos, muy anchos, 1,57 veces más largos que anchos tomados conjuntamente, con su máxima anchura en la parte media; húmeros redondeados, nada salientes hacia delante; estrías de puntos netas y bien indicadas; intervalos impares con unos pocos puntos en la zona apical. Machos con las antenas más gruesas.
Distribución
Pollock & Löbl (2008) indican su presencia en: Alemania, Austria, Bélgica, Bosnia, Bulgaria, Croacia, Dinamarca, Eslovaquia, España, Francia, Holanda, Hungría, Irlanda, Italia, Polonia, Rumania, República Checa, Serbia, Suecia y Ucrania. En le Península sólo se ha localizado en el norte de España: Montes de Vitoria (Araba), Cataluña (Girona), Lumbreras, Villoslada de Cameros (La Rioja), Garde, Irañeta, Lizaso, Parque Natural del Señorío de Bertiz (Navarra) (Alonso Román & Bahillo de la Puebla, 2019; Pérez Moreno, 2005; Pérez Moreno & Moreno Grijalba, 2009; Recalde Irurzun & San Martín, 2015, 2016; Recalde Irurzun et al., 2017) (Fig. 34).
A J. Bookwalter, Girona, el habernos permitido estudiar parte de los ejemplares recolectados en Andorra para su tesis. A Joaquim Soler, de Garrigella, a Eduard Piera, de l’Arboçar, a Antonio San Martín, de Pamplona, a Jordi Calaf, técnico del Parc de la Muntanya de Montserrat y a Bartomeu Borras, Pau Federico y Marc Fernández, director y técnicos respectivamente del Paratge Natural d’Interès Nacional de l’Albera, la ayuda y esfuerzos realizados en las recolecciones sistemáticas efectuadas en los diferentes parajes protegidos.
Referencias
Alonso Román, I. & Bahillo de la Puebla, P. 2019. Catálogo de los coleópteros de la Comunidad Autónoma del País Vasco. Heteropterus, Revista de Entomología, 19 (1): 1-266.
Alonso Zarazaga. M. A. 2014. On the identity of some weevil species described by Johann Christian Fabricius (1745–1808) in the Museum of Zoology of Copenhagen (Coleoptera, Cucujoidea, Curculionoidea, Tenebrionoidea). ZooKeys, 451: 61-91.
Bahillo de la Puebla, P. & Alonso Román, I. 2018. Nuevos registros de coleópteros del País Vasco (Insecta: Coleoptera). Heteropterus Revista de Entomología, 18 (1): 71-80.
Barnouin, T. 2014. Salpingidae. P. 556-557. In. Catalogue des Coléoptères de France. Supplément au Tome XXIII. Association Roussillonnaise d’Entomologie. Perpignan. 1052 p.
108 Butlletí ICHN 85 (3), 2021
FLORA ET FAUNA
Figura 33. Habitus de Vincenzellus ruficollis (Panzer, 1794).
A. VIÑOLAS ET AL.
Figura 34. Mapa con la distribución de Vincenzellus ruficollis
REVISIÓN DE LA FAMILIA SALPINGIDAE LEACH, 1815 EN EL ÁREA IBEROBALEAR
Barnouin, T. & Zagatti, P. 2017. Les Salpingidae de la faune de France. (Coleoptera Tenebrionoidea). L’Entomologiste, 73 (6): 353-386.
Champion, G. C. & Chapman, T. A. 1904. V. An Entomological Excursion to Moncayo, N. Spain. Transactions of the Royal Entomological Society of London, 52: 81-102.
Cuní I Martorell, M. & Martorell i Peña, M. 1876. Catálogo metódico y razonado de los coleópteros observados en Cataluña Tomas Gorchs y Cia. Barcelona. 360 p.
Dajoz, R. 1969. Nouveaux Colydiidae anophthalmes du Chili (Col.). Bulletin de la Société Entomologique de France, 74 (9-10): 230-234.
Dajoz, R. 1977. Coléoptéres Colydiidae et Anommatidae Paléartiques. Faune de l’Europe et du bassin méditerranéen, 8. Masson. Paris. 272 p.
Diéguez Férnandez, J. M. 2012. Coleópteros nuevos o interesantes para la fauna catalana (Insecta: Coleoptera). Heteropterus, Revista de Entomología, 12 (1): 123-128.
Diéguez Fernández, J. M. 2015. Registros interesantes de coleópteros para España y Marruecos (Insecta: Coleoptera). Arquivos Entomolóxicos, 14: 27-31.
Español, F. 1955. Colidíidos endógenos del levante español (Col. Cucujoidea). Publicaciones del Instituto de Biología Aplicada, 21: 101-106.
Español, F. 1971. Sobre algunos coleópteros asociados a Bostríquidos, Líctidos y Anóbidos. Boletín del Servicio de Plagas Forestales, 14 (27): 17-21.
Etxebeste Larrañaga, I., Lencia Gutierrez, J. I. & Pajares Alonso, J. A. 2013.Respuesta de Ips sexdentatus (Col. Curculionidae, Scolytinae) y de coleópteros saproxílicos asociados a la variación en la composición feromonal. Actas del 6º Congreso forestal Español. Montes: servicio y desarrollo rural 6CFE01-382: 2-14.
Ferrer, J. de & Quetglas, J. 2004. Sobre la presencia de Paromalus (Isomalus) luderti Marseul 1862 (Coleoptera, Histeridae), en la provincia de Cádiz (España), y nuevo hábitat para el mismo. Boletín de la Sociedad Entomológica Aragonesa, 35: 282.
Fuente, J. M. de la 1928. Catálogo sistemático-geográfico de los Coleópteros observados en la Península Ibérica, Pirineos propiamente dichos y Baleares (Continuación). Boletín de la Sociedad Entomológica de España, 11: 74-89.
Fuente, J. M. de la 1932. Catálogo sistemático-geográfico de los Coleópteros observados en la Península Ibérica, Pirineos propiamente dichos y Baleares (Continuación). Boletín de la Sociedad Entomológica de España, 15: 75-90.
Grosso-Silva, J. M. 2002. Registros interessantes de coleópteros (Insecta, Coleoptera) para Portugal (3ª nota). Primeiro registro ibérico de Pediacus dermestoides (Fabricius, 1792) (Cucujidae). Boletín de la Sociedad Entomológica Aragonesa, 31: 49-54.
Lencina, J. L., Baena, M., Gallego, D. & Andújar, C. 2008. Un nuevo representante y nuevos registros de la familia Salpingidae Leach, 1815 (Coleoptera) en la Península Ibérica. Boletín de la Sociedad Entomológica Aragonesa, 43: 421-423.
Micó, E., Marcos García, M. A. & Galante, E. (Eds.). 2013. Los insectos saproxílicos del Parque Nacional de Cabañeros Organismo Autónomo del Parques Nacionales, Ministerio de Agricultura, Alimentación y Medio Ambiente, Madrid. 139 p.
Molino Olmedo, F. 1997a. Coleópteros nuevos para la Península Ibérica y España. Boletín de la Asociación española de Entomología, 21 (1-2): 107.
Molino Olmedo, F. 1997b. Algunos coleópteros nuevos para Andalucía. Zoologica baetica, 8: 239-241.
Otero, J. C., García, A. & López, M. J. 2012. Sobre algunas especies de Cryptophagidae, Latridiidae y Salpingidae (Coleoptera) capturados en medios subterráneos de España. Boletín de la Asociación española de Entomología, 36 (1-2): 137-144.
Outerelo, R. & Gamarra, P. 2011. Salpingus planirostris (Fabricius,
1787) (Coleoptera: Salpingidae), primera cita para Galicia (N.O. Península Ibérica). Arquivos Entomologicos 5: 105-108.
Pagola-Carte, S., Zabalegui, I., Recalde Irurzún J. I., San Martín Moreno, A. F., Bahillo de la Puebla, P. & Petitpierre. E. 2007. Algunos coleópteros interesantes (Insecta: Coleoptera) del Parque Natural de Aiako Hatria (Gipuzkoa, norte de la Península Ibérica). Heteropterus, Revista de Entomología, 7 (1): 77-90.
Pérez Fernàndez, A. 2007. Algunas notas sobre invertebrados capturados en cuevas de Jaén. Espeleo, Bioespeleo, 19: 27-28.
Pérez Moreno, I. 2005. Aportación al conocimiento de la familia Salpingidae (Coleoptera) en la Península Ibérica. Boletín de la Sociedad Entomológica Aragonesa, 37: 141-147.
Pérez Moreno, I. & Moreno Grijalba, F. 2009. Los coleópteros saproxílicos del Parque Natural de Sierra de Cebollera (La Rioja). Ciencias de la Tierra, 28. Gobierno de La Rioja. Instituto de Estudios Riojanos. Logroño. 180 p.
Pérez-Moreno, I., Recalde.Irurzun, J. I. & San Martin Moreno, A. 2020. Coleópteros saproxílicos notables (Coleoptera) de las dehesas riojanas del Sistema Ibérico septentrional. Boletín de la Sociedad Entomológica Aragonesa, 66: 107-122.
Piera, E., Viñolas, A., Soler, J., Buqueras, X. & Muñoz-Batet, J. 2017. Els coleòpters saproxílics de les pinedes del Bosc de Poblet. El Podall (2016): 321-352.
Pollock D. A. & Löbl, I. 2008. Salpingidae. In: Löbl, I., Smetana, A. (Ed.) Catalogue of Palaearctic Coleoptera. Vol. 5. Apollo Books, Stenstrup, 417-421.
Ramilo, P. E., Galante, E. & Micó, E. 2017. Intra-annual patterns of saproxylic beetle assemblages inhabiting Mediterranean oak forests. Journal of Insect Conservation, 21: 607-620.
Ramírez Hernández, A., Micó, E. & Galante, E. 2014. Temporal variation in saproxylic beetle assemblages in a Mediterranean ecosystem. Journal of Insect Conservation. DOI10.1007/s10841-014-9706-9
Recalde Irurzun, J. I. & San Martín Moreno, A. F. 2015. Aproximación a la fauna de escarabajos (Coleoptera) del Parque Natural del Señorío de Bertiz (Navarra). Heteropterus, Revista de Entomología, 15 (1): 43-57.
Recalde Irurzun, J. I. & San Martín Moreno, A. F. 2016. Escarabajos saproxílicos (Coleoptera) de dos bosques pirenaicos de Navarra. Heteropterus, Revista de Entomología, 16 (1): 53-69.
Recalde Irurzun, J. I., San Martín Moreno, A. F. & Bizkai, J. A. 2017. Los coleópteros saproxílicos (Coleoptera) de un pasto arbolado del norte de la Península Ibérica. Boletín de la Sociedad Entomológica Aragonesa, 61: 85-98.
Spilman, T. J. 1952. The male genitalia of the Nearctic Salpingidae. The Coleopterists Bulletin, 6 (1): 9-13.
Viñolas, A., Muñoz-Batet, J. & Soler, J. 2012. Nuevos registros y nuevas localizaciones de coleópteros para la Península Ibérica y en especial del Paratge Natural de l’Albera, Girona (Coleoptera). Elytron. 25: 3-63.
Viñolas, A., Muñoz-Batet, J. & Soler, J. 2015. Els coleòpters saproxílics de la casa forestal del Tillar, serra de Prades, Tarragona. IV Jornades sobre el bosc de Poblet i les muntanyes de Prades: 361-370.
Viñolas, A., Muñoz, J., Mencuccini, M. & Benvenuti, F. 2013. Nuevos datos sobre Rushia parreyssi (Mulsant, 1856), Melandrydae Leach, 1815 y otros coleópteros interesantes de la sierra de Prades, Tarragona (Coleoptera). Orsis, 27: 29-51.
Viñolas, A., Muñoz-Batet, J., Bentanachs, J. & Masó, G. 2014. Catálogo de los coleópteros del Parque Natural del Cadí-Moixeró, Cataluña, Península Ibérica. Coleopterological Monograph, 5. 155 p.
Viñolas, A., Piera, E. & Muñoz-Batet, J. 2019. Nuevas citas de coleópteros para Aragón, Península Ibérica (Coleoptera). Butlletí de la Societat Catalana d’Història Natural, 83: 215-217.
Viñolas, A., Recalde-Irurzun, J. I. & Muñoz-Batet, J. 2018. Noves aportacions al coneixement de la fauna coleopterològica de la península Ibèrica i illes Canàries. Nota 3a, amb revisió dels Lissodema Curtis, 1833 ibèrics (Coleoptera, Salpingidae, Lissodeminae). Butlletí de la Societat Catalana d’Història Natural, 82: 41-52.
109 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
Butlletí de la Institució Catalana d’Història Natural, 85 (3): 111-113. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition)
NOTA BREU
Erigeron blakei Cabrera (Asteraceae), an alien species confirmed for the Iberian Peninsula
Erigeron blakei Cabrera (Asteraceae), espècie aŀlòctona confirmada a la península Ibèrica
Lluís Vilar* & Llorenç Sáez**, ***
* Unitat de Botànica, Laboratori d’Anàlisi i Gestió del Paisatge, Facultat de Ciències, Campus de Montilivi, Universitat de Girona. C/ M. Aurèlia Campmany, 69, 17003, Girona
**Systematics and Evolution of Vascular Plants (UAB) – Associated Unit to CSIC. Dept. Biologia Animal, Vegetal i Ecologia, Facultat de Biociències, Universitat Autònoma de Barcelona, ES08193 Bellaterra, Barcelona
***Societat d’Història Natural de les Illes Balears, c/ Margarida Xirgu 16, ES07003 Palma de Mallorca, Illes Balears
Correspondence author: Lluís Vilar. A/e: lluis.vilar@udg.edu
Rebut: 23.08.2021. Acceptat: 02.09.2021. Publicat: 30.09.2021
Erigeron blakei Cabrera [≡Conyza blakei (Cabrera) Cabrera]
Gironès: Vilablareix, gardened areas and waysides, 31TDG8244, 99 m, 8-VII-1999, L. Vilar (HGI 16281); ibidem, 10-VII-2021, L. Vilar (HGI 24227). Figure 1.

Eight alien Erigeron L. species formerly included in Conyza Less. are present in Europe and the Mediterranean basin according to Euro+Med PlantBase (Euro+Med, 2006-2021): Erigeron bilbaoanus (J. Rémy) Cabrera; E. blakei Cabrera; E. bonariensis L.; E. canadensis L.; E. daveauanus (Sennen) Greuter; E. floribundus (Kunth) Sch. Bip.; E. primulifolius (Lam.) Greuter and E. sumatrensis Retz. However, in recent taxonomic treatments two of the species accepted in Euro+Med PlantBase are usually considered synonymous with other taxa or have an uncertain taxonomic value. This is the case of E. bilbaoanus, included in the synonymy of E. floribundus [≡Conyza floribunda Kunth] (Stace, 2010; Pyke, 2020; Liendo et al., 2021) and E. daveauanus, morphologically close to E. sumatrensis [≡Conyza sumatrensis (Retz.) E. Walker] (Pyke, 2020) or included in the synonymy of the latter species (Stace, 2010; Nesom, 2018; Liendo et al., 2021).
In Europe, these alien species of Erigeron formerly included in Conyza usually occur in strongly disturbed habitats (waste places, roadsides, railways, gardens and cultivated fields) but also in more or less disturbed areas of native vegetation such as sand dunes or grasslands (Thébaud & Abbott, 1995; Liendo et al., 2021; Verloove, 2021).
Erigeron blakei [Conyza blakei (Cabrera) Cabrera] is a species native to South America, from South Brazil to North Argentina (POWO, 2021). It is naturalized and established in Southern France (Tison & de Foucault, 2014; Tison et al., 2014; Euro+Med, 2006-2021). The presence of this species in other European countries has been ruled out or is uncertain (Liendo et al., 2021). Erigeron blakei was found for the first time in Europe in the late 1960s (Jovet & Vilmorin, 1975) and started to spread rapidly in southeastern France in late 20th-century (Kerguelen et al. 1987), invading mainly old fields and vineyards (Thébaud & Abbott, 1995).
According to Laínz (2002) and Pyke (2020) E. blakei was reported in error for the Iberian Peninsula by several authors
111 Butlletí ICHN 85 (3), 2021
BREU
NOTA
DOI: 10.2436.20.1502.01.80
Figure 1. Erigeron blakei Cabrera (HGI 24227): Habit of a flowering plant (left); lower stem leaf with lobes (above right); detail of the secondary inflorescence (bottom right).
(Malagarriga, 1965; Casasayas, 1989; Bolòs & Vigo, 1996). These reports are mainly due to confusion with E. floribundus (Kunth) Schultz-Bip. On the other hand, E. blakei was neither included in Flora iberica (Morales, 2019) nor in a recent revision of the alien Erigeron species in central-northern Spain (Liendo et al., 2021), although the latter authors suggested that its present (or future) occurrence in northern Spain cannot be ruled out.
Erigeron floribundus and E. blakei are closely related species (Thébaud & Abbott, 1995; Tison & Foucault, 2014; Tison et al. 2014; Nesom, 2018). Both taxa are annual or short-lived perennial, have reduced vestiture of the stems and leaves (subglabrous to sparsely hirsute), lower stem leaves dentate or narrowly lobed, small capitulae (c. 3-4 mm long), glabrous or almost glabrous involucral bracts, inner (tubular) florets mostly 5-lobbed and ligules absent or rudimentary, not exceeding involucre. Plants of E. floribundus are distinct in their relatively broad leaves (rachis usually > 3 mm wide, see Tison & de Foucault, 2014 and Tison et al., 2014), middle and upper stem leaves lanceolate, with visible secondary venation and the secondary inflorescences are broadly paniculate, whereas those of E. blakei have narrow lower stem leaves (rachis up to 3 mm wide and lobes up to 5 mm long in our specimens) middle and upper stem leaves linear, with a single vein visible and the secondary inflorescences are cylindrical or subcylindrical. Erigeron blakei has also an appearance similar to E. canadensis, a rather variable species with several infraspecific taxa described (Sell & Murrell, 2006; Pyke, 2020; Verloove, 2021). Both species share cylindrical or subcylindrical secondary inflorescences (conical in E. canadensis var. glabratus A. Gray, according to Pyke, 2020) and small capitulae. However, E. canadensis has not hirsute stems, inner (tubular) florets mostly 4-lobbed (4 or 5-lobbed in E. canadensis var. glabratus A. Gray, according to Pyke, 2020) and ligules always present, distinctly exceeding involucre. Furthermore, our observations indicate that the plants here referred to E. blakei, can be short-lived perennial, whereas E. canadensis is annual (Thébaud & Abbott, 1995; Diez de Ulzurrun et al., 2018). Main morphological characters differentiating Erigeron blakei, E. canadensis and E. floribundus are summarised in Table 1.
Spontaneous plants of Erigeron blakei were recently found in 2021 at the edge of garden in an urban area in Girona province (HGI 24227). The population consists of few
plants occupying an area of about 10 square meters. Interestingly, our recent revision of the materials of Erigeron formerly included in Conyza deposited at HGI, has revealed that E. blakei was already collected in the same locality in 1999 (HGI 16281). In addition, one of us (LV) remembers seeing this plant more than 35 years in the same locality. Therefore, E. blakei could be considered locally naturalized.
The presence of E. blakei in northeastern Iberian Peninsula is not surprising, since it is naturalized in Southern France (Tison et al., 2014) and should be expected in disturbed areas in northeastern Catalonia.
Acknowledgements
The authors are grateful to Patricia Diez de Ulzurrun, Guy Nesom, John L. Strother and Filip Verloove, for providing crucial information for the identification of the species.
References
Bolòs, O. & Vigo, J. 1996. Flora dels Països Catalans, vol. 3. Ed. Barcino, Barcelona. 1230 p.
Casasayas, T. 1989. La flora aŀlòctona de Catalunya. Tesis Doctoral, Universitat de Barcelona, Barcelona. 880 p.
Diez de Ulzurrun, P., Acedo, M. B., Garvano, M. E., Gianelli, V. & Ispizúa, V. 2018. Caracterización morfológica de Conyza blakei, Conyza bonariensis var. bonariensis, Conyza sumatrensis var. sumatrensis y Conyza lorentzii en el sudeste bonaerense (Argentina). Boletín de la Sociedad Argentina de Botánica, 53: 359-373.
Kérguelen, M., Bosc, G. & Lambinon, J. 1987. Données taxonomiques, nomenclaturales et chorologiques pour une révision de la flore de France. Lejeunia, 120: 1-264.
Euro+Med. 2006-2021. Euro+Med PlantBase - the information resource for Euro-Mediterranean plant diversity. http://ww2. bgbm.org/EuroPlusMed [accessed 10 Aug 2021].
Laínz, M. 2002. Conyza blakei (Cabrera) Cabrera (Compositae), especie Peninsular? Anales del Jardín Botánico de Madrid, 59: 352-353.
Liendo, D., García-Mijangos, I., Biurrun, I., & Campos, J. A. 2021. Annual weedy species of Erigeron in the northern Iberian Peninsula: a review. Mediterranean Botany, 42: e67649. https:// doi.org/10.5209/mbot.67649
Malagarriga, T. 1965. Flora analítica de Barcelona. La Salle Bonanova, Barcelona. 271 p.
Table 1. Morphological comparison among Erigeron blakei, E. canadensis and E. floribundus.
E. blakei
Lower stem leaves
E. canadensis
Dentate or narrowly lobed; rachis Entire to dentate
E. floribundus
Dentate or narrowly lobed; rachis usually ≤ 3 mm wide usually > 3 mm wide
Upper stem leaves Linear, < 1.5 mm wide
Linear to oblanceolate, usually Linear to lanceolate, > 2 mm wide > 2 mm wide
Secondary inflorescences Cylindrical or subcylindrical Subcylindrical to conical Broadly paniculate
Ligules Rudimentary, or absent, not ligules always present, distinctly Rudimentary, or absent, not exceeding involucre, <0.4 mm long exceeding involucre, usually exceeding involucre, < 0.4 mm long 0.5-1.5 mm long
Inner tubular florets mostly 5- lobbed mostly 4- lobbed mostly 5- lobbed
112 Butlletí ICHN 85 (3), 2021 NOTA BREU L. VILAR & L. SÁEZ
Morales, R. 2019. Conyza Less. In: Benedí, C., Buira, A., Rico, E., Crespo, M.B., Quintanar, A. & Aedo, C. (Eds.). Flora iberica vol. XVI (III). Pp. 2022–2027. Real Jardín Botánico, CSIC, Madrid.
Nesom, G. 2018. Erigeron floribundus and E. sumatrensis (Asteraceae) in the USA and Mexico. Phytoneuron, 2018–27: 1–19.
POWO 2021. Plants of the World Online. http://www. plantsoftheworldonline.org [Accessed 30 July 2021]
Pyke, S. 2020. Conyza (Asteraceae): una valoración crítica basada en las poblaciones de Cataluña, España. Collectanea Botanica (Barcelona), 39: e005.
Sell, P. & Murrell, G. 2006. Flora of Great Britain and Ireland. Vol. 4 Campanulaceae – Asteraceae. Cambridge University Press, Cambridge. 624 p.
Stace, C. A. 2010. New flora of the British Isles, third ed. Cambridge University Press, Cambridge. 1232 p.
Thébaud, C. & Abbott, R.J. 1995. Characterization of invasive Conyza species (Asteraceae) in Europe: quantitative trait and isozyme analysis. American Journal of Botany, 82: 360-368.
Tison, J.-M. & de Foucault, B. (Eds.). 2014. Flora Gallica: Flore de France. Soc. Bot. France. Biotope Éd., Mèze. 1195 p.
Tison, J.-M.; Jauzein, P. & Michaud, H. 2014. Flore de la France méditerranéenne continentale. Naturalia. Turriers. 2078 pp.
Verloove, F. 2021. Erigeron L. On: Manual of the Alien Plants of Belgium. Botanic Garden Meise, Belgium. At: alienplantsbelgium. be. [Accessed 16 Aug 2021]
113 Butlletí ICHN 85 (3), 2021
BREU
NOTA
ERIGERON BLAKEI CABRERA (ASTERACEAE), AN ALIEN SPACIES CONFIRMED FOR THE IBERIAN PENINSULA
Butlletí de la Institució Catalana d’Història Natural, 85 (3): 115-117. 2021
NOTA BREU
ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition)
Nova població de l’espècie amenaçada Asphodelus ramosus subsp. ramosus (Asphodelaceae) a Catalunya
A new population of the threatened species Asphodelus ramosus subsp. ramosus (Asphodelaceae) in Catalonia
Genís Puig* & Llorenç Sáez*
* Sistemàtica i Evolució de Plantes Vasculars (UAB) –Unitat Associada al CSIC. Departament de Biologia Animal, Vegetal i Ecologia, Facultat de Biociències. Universitat Autònoma de Barcelona. 08193 Bellaterra, Barcelona.
Autor per a la correspondència: Genís Puig. A/e: genispuigsurroca@gmail.com
Rebut: 10.07.2021. Acceptat: 02.09.2021. Publicat: 30.09.2021
Asphodelus ramosus L. subsp. ramosus
Baix Empordà: Torroella de Montgrí, Puig de la Fonollera, 31TEG1551, 15 m, brolles sobre gresos calcaris i conglomerats eocènics, 9-V-2021, G. Puig Surroca (herb. pers.) (Figs, 1, 2).

Asphodelus ramosus L. subsp. ramosus és un endemisme mediterrani molt rar a la península Ibèrica (Díaz Lifante, 2013) la presència del qual, a la Catalunya autònoma, no ha estat confirmada fins temps molt recents (Aymerich & Sáez, 2021). Pocs anys abans, els mateixos autors (Aymerich & Sáez, 2015) fan un recull de les dades relatives a la presència d’aquesta espècie a Catalunya. De fet, fins al moment actual només es coneixia una població que es troba entre el Puig

115 Butlletí ICHN 85 (3), 2021
BREU
NOTA
DOI: 10.2436.20.1502.01.81
Figura 1. Detalls d’Asphodelus ramosus subsp. ramosus del Puig de la Fonollera. branca de la inflorèscencia (esquerra); detall de la inflorescència (dreta, a dalt); fruits (dreta, a baix).
Figura 2. Exemplars d’Asphodelus ramosus subsp. ramosus del Puig de la Fonollera.
de Cervera i el litoral de les Freses (municipi de Portbou, Alt Empordà), molt a prop de la línia fronterera. Encara que Aymerich & Sáez (2015) no van poder trobar evidències de la presència d’A. ramosus a la península del cap de Creus, -àrea d’on existeixen citacions bibliogràfiques versemblants (Cadevall, 1933, sub A. microcarpus Viv.) sobre la base de les quals, probablement, Díaz Lifante (2013) va acceptar la presència d’aquesta espècie a Catalunya-, sembla molt versemblant la presència d’aquest tàxon en aquesta darrera zona de l’Alt Empordà.
En unes prospeccions realitzades al maig de 2021 a la zona de la Fonollera, vàrem trobar uns exemplars d’un Asphodelus de fulles planes i carenades (de fins a 35 mm d’amplada), però que, a diferència d’A. cerasiferus J Gay (una espècie que és coneguda del massís del Montgrí i les Medes), aquests presentaven uns fruits de mida notablement petita, el que va atreure immediatament la nostra atenció. Un estudi més detallat de les mostres obtingudes, de les fotografies i una revisió de la bibliografia, va posar de manifest que es tractava d’A. ramosus subsp. ramosus. La localitat aquí aportada confirmaria la seva presència, a Catalunya, en zones més meridionals del que s’havia documentat fins al moment.
Les plantes de la localitat del Puig de la Fonollera presenten les característiques morfològiques pròpies de la sect. Asphodelus (Díaz Lifante & Valdés 1996; Díaz Lifante, 2013) i, més concretament, del grup d’A. ramosus: uns fruits relativament petits (Figura 1), la part basal dels estams que s’estreny de forma abrupta al seu àpex, tija molt ramificada, bràctees bru-clares o blanquinoses, almenys al marge, i floració simultània. Les espècies incloses en aquest grup serien, a més del tàxon que ens ocupa, A. aestivus Brot. i A. serotinus Wolley-Dod.
Segons Díaz Lifante (2013), un dels caràcters diagnòstics d’A. ramosus davant les dues darreres espècies és la presència de pedicels de 0,8-1,3 mm de diàmetre a la fructificació, mentre que A. aestivus i A. lusitanicus Cout. (aquesta darrera és una altra espècie de la sect. Asphodelus relacionada amb A. ramosus) presenten uns pedicels de 0,5-0,7 mm de diàmetre. L’estudi dels nostres exemplars posa de manifest que el diàmetre dels pedicels és de 0,9-1,2 mm (mesurats a la meitat de la seva longitud), i per tant aquestes mides entren dins el rang assignat per a A. ramosus.
La mida de les càpsules de les mostres estudiades és 5-7 x 4,5-5,7 mm i, per tant, aquestes entrarien dins del rang de variació de la subsp. ramosus (Díaz Lifante & Valdés 1996; Díaz Lifante, 2013), ja que la subsp. distalis Z. Díaz & Valdés, un tàxon endèmic del SW ibèric, NW del Marroc i Canàries (Díaz Lifante & Valdés 1996; Díaz Lifante, 2013), presenta uns fruits notablement més grossos.
La població d’Asphodelus ramosus subsp. ramosus del Puig de la Fonollera constava, l’any 2021, de 389 individus -243 dels quals reproductors- distribuïts en 5 petits rodals que ocupaven 970 m² (Figura 2). El tàxon es troba tant al planell superior com en el vessant SW del puig. En el primer cas, es fa sobre sòls esquelètics on predominen els pradells terofítics (rodal 1) i en petites clarianes de brolles arbrades amb Pistacia lentiscus, Rhamnus alaternus i Olea europaea (rodal 2). En el segon cas, es troba en herbassars relativament humits molt més oberts i amb peus esparsos de Ligustrum vulgare, Quercus pubescens i Rubus ulmifolius (i convivint amb Asphodelus fistulosus) (rodals 3 i 4) i al marge de boscos de Pinus halepensis (rodal 5).
La població està emparada per tres figures de protecció: la Xarxa Natura 2000, el Parc Natural del Montgrí, les Illes Medes i el Baix Ter (PNMMBT) i la Reserva natural parcial dels aiguamolls del Baix Ter (GOV/112/2006, de 5 de setembre; Llei 15/2010, del 28 de maig).
El principal factor de risc per a la població és el progressiu tancament natural de la vegetació. Entre 1956 i 2020, la superfície dels espais oberts del Puig de la Fonollera va passar de 26.616 m² a 8.918 m² (variació calculada a partir d’ortofotografies de l’Institut Cartogràfic i Geològic de Catalunya; http:// www.icc.cat/vissir3), la qual cosa implica una reducció del 66,5 %. Aquest factor afecta sobretot el rodal 2, situat en els darrers claps mínimament oberts d’una brolla arbrada. També podria explicar la situació crítica del rodal 5, format per tan sols 14 peus (Taula 1) ubicats en una estreta franja entre un bosc molt dens de Pinus halepensis i un camí, si bé aquesta podria ser en realitat el producte d’una colonització recent. Pel que fa a la resta de rodals, també podrien veure’s greument amenaçats pel tancament de la vegetació, que molt probablement en els propers anys anirà constrenyent l’extensió de l’hàbitat potencial.
Per altra banda, una freqüentació antròpica eventualment excessiva podria arribar a constituir un segon factor de risc. Aquesta, però, no sembla haver tingut fins ara un impacte significatiu sobre l’espècie (ni tan sols en el rodal 1, situat entre uns antics búnquers molt visitats) més enllà de la destrucció d’escapus, observada directament al camp en 4 exemplars del rodal 4.
En el cas particular del rodal 5, caldria considerar també les amenaces derivades de la seva ubicació al marge d’un camí. En altres trams d’aquest mateix camí es va observar, el 18/04/2021, que les vores havien estat ruixades amb herbicida. La competència amb espècies ruderals més ben adaptades a aquest tipus d’ambients, doncs, podria fer-lo acabar desapareixent. D’altra banda, però, la baixa cobertura llenyosa associada a la franja del marge més immediata al camí resulta favorable per aquest rodal.
116 Butlletí ICHN 85 (3), 2021 NOTA BREU G. PUIG & L. SAEZ
Rodal 1 Rodal 2 Rodal 3 Rodal 4 Rodal 5 Nre. d’individus reproductors 60 72 7 98 6 Nre. d’individus no reproductors 37 66 1 34 8 Àrea d’ocupació (m²) 318 255 42 257 98
Taula 1. Dades poblacionals per als diversos rodals d’Asphodelus ramosus subsp. ramosus a la localitat del Puig de la Fonollera.
El desbrossament indiscriminat, en el marc de les tasques de prevenció d’incendis, no constituiria un veritable factor de risc. El 05/06/2021 es va constatar com aquest factor havia causat la destrucció total del rodal 1 (en la part aèria dels seus exemplars). El període d’activitat fotosintètica i generació de reserves, doncs, es va veure bastant escurçat. Tanmateix, al tractar-se d’un geòfit i haver arribat ja a produir càpsules madures, probablement l’afectació va ser lleu. I, en qualsevol cas, l’eliminació de tot l’estrat arbustiu va generar un ambient molt més obert i, per tant, molt més favorable per a l’espècie. És per això que aquest factor no constitueix realment una amenaça i cal considerar-lo fins i tot beneficiós.
Malgrat la descoberta d’aquesta nova població aquí documentada, no es contempla un canvi de categoria IUCN (2012) respecte a la recent qualificació d’Asphodelus ramosus subsp. ramosus com a EN a nivell de Catalunya (Aymerich & Sáez, 2021), ja que únicament es coneixen dues localitats, i una d’elles (la localitzada a Portbou) està sotmesa a fluctuacions en el nombre d’efectius reproductors (uns 660 segons les observacions del primer autor realitzades el 04/07/2021) a causa dels incendis. Sí que es pot descartar, però, l’efecte rescat, ja que la nova localitat implica que no tota la població administrativament catalana està vinculada a poblacions extradministratives. Per tant, la seva categoria i els criteris que la justificarien serien EN B2ac(i,ii,iv).
Agraïments
Volem agrair a Zoila María Díaz Lifante la confirmació de la identitat del tàxon i a Pere Aymerich alguns comentaris relatius a la categoria de risc.
Bibliografia
Aymerich, P. & Sáez, L. 2015. Comentaris i precisions previs a la Checklist de la flora de Catalunya (nord-est de la península Ibèrica). Orsis, 29: 1-68.
Aymerich, P. & Sáez, L. 2021. Llista Vermella de la flora vascular de Catalunya. Actualització any 2020. Monografies de la Institució Catalana d’Història Natural, 2: 1-100.
Díaz Lifante, Z. 2013. Asphodelus L. In: Rico, E.; Crespo, M.B.; Quintanar, A.; Herrero, A.; Aedo. C. (eds.) Flora iberica. Vol. XX. Liliaceae-Agavaceae: 276-308. Real Jardín Botánico, CSIC. Madrid.
Díaz Lifante, Z. & B. Valdés 1996. Revisión del género Asphodelus L. (Asphodelaceae) en el Mediterráneo Occidental. Boissiera, 52: 1-189.
IUCN 2012. Red List Categories and Criteria: Version 3.1. Second edition. Gland, Switzerland and Cambridge. portals. iucn.org/ library/node/10315
117 Butlletí ICHN 85 (3), 2021 NOTA BREU NOVA POBLACIÓ DE L'ESPÈCIE AMENAÇADA ASPHODELUS RAMOSUS SUBSP. RAMOSUS A CATALUNYA
Sobre la presència d’Hydropunctaria scabra a la península Ibèrica, i notes sobre altres líquens aquàtics de Les Planes de Son i mata de València
Cristian Romero* & Pere Navarro-Rosinés*
* Secció de Botànica i Micologia. Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals (BEECA). Institut de Recerca de la Biodiversitat (IRBio). Facultat de Biologia. Universitat de Barcelona, Diagonal, 643. 08028 Barcelona.
Autor per a la correspondència: Pere Navarro-Rosinés. A/e: pnavarro@ub.edu
Rebut: 20.05.2021; Acceptat: 02.09.2021; Publicat: 30.09.2021
Resum
S’aporten diverses observacions de líquens hidròfils per a Catalunya, que procedeixen totes de la mateixa localitat dels Pirineus, situada al municipi de l’Alt Àneu (Pallars Sobirà, prov. de Lleida). Aquestes observacions constitueix una addició al catàleg dels líquens de l’àrea que es denominà «Les Planes de Son i mata de València», publicat prèviament en un estudi més ampli i integral de la zona. Es pot destacar la presència d’Hydropunctaria scabra, una espècie que, segons la informació disponible, és citada per primera vegada a la península Ibèrica. Per aquest tàxon s’inclou una descripció detallada, acompanyada d’iconografia fotogràfica i dels comentaris morfològics i corològics oportuns. Verrucaria pachyderma és també una espècie amb interès corològic per Catalunya, ja que anteriorment era només coneguda d’una localitat de la Vall d’Aran.
Paraules clau: Hydropunctaria, Verrucaria, líquens aquàtics, Catalunya, Espanya.
Abstract
About the presence of Hydropunctaria scabra in the Iberian Peninsula, and notes of other aquatic lichens of Les Planes de Son i Mata de Valencia
Several observations of different hydrophilic lichens in Catalonia are commented, all of them come from the same Pyrenean locality, in the municipality of Alt Àneu (Pallars Sobirà, Lleida province). These observations constitute an addition to the lichen checklist of the area called «Les Planes de Son i mata de València», previously published in a larger and more comprehensive study of the area. We would like to highlight the presence of Hydropunctaria scabra, a species, according to the available information, mentioned for the first time in the Iberian Peninsula. A detailed description is included for this taxon, accompanied by photographic iconography and the appropriate morphological and chorological comments. Verrucaria pachyderma is also a species of chorological interest in Catalonia, as it was only known from a single locality in the Aran Valley.
Key words: Hydropunctaria, Verrucaria, hydrophilic lichens, Catalonia, Spain.
Introducció
Durant l’estudi dels líquens de les Planes de Son i la mata de València es recoŀlectaren, en una mateixa localitat, diferents líquens que creixien sobre substrats rocosos inundats o adjacents al curs d’aigua d’un rierol i, per tant, sotmesos a inundacions temporals. Aquests exemplars, tant per la impossibilitat de reconèixer les espècies directament al camp com per la dificultat d’estudi i de identificació al laboratori, degut a l’escassa bibliografia disponible sobre líquens aquàtics en els anys de l’estudi de la zona (anys 2006 i 2007), no pogueren ser identificats amb certesa, i no van ser inclosos en el catàleg dels líquens del territori (Llop et al., 2010).
Posteriorment a la publicació d’aquest treball sobre els líquens de les Planes de Son, el coneixement dels líquens
aquàtics ha avançat considerablement amb la publicació de nous treballs específics d’aquests ambients (Thüs & Schultz, 2009; Coste, 2011; Orange, 2012, 2013). Una revisió d’aquests exemplars ens ha permès ara identificar moltes de les espècies. Amb la finalitat de completar el catàleg del territori en qüestió, i també per la novetat o interès corològic que presenta alguna de les espècies, considerem oportú ferne menció en aquest treball.
Entre els líquens aquàtics presents al territori de Les Planes de Son i mata de València, destaca la presència d’Hydropunctaria scabra, una espècie per a la qual no es tenia constància de la seva presència a la península Ibèrica.
El gènere Hydropunctaria va ser descrit com a resultat de l’estudi molecular de la família Verrucariaceae (Gueidan et al., 2009), per a segregar del gènere Verrucaria algunes
119 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA GEA, FLORA ET FAUNA
Butlletí de la Institució Catalana d’Història Natural, 85 (3): 119-124. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition) DOI: 10.2436.20.1502.01.82
GEA,
espècies aquàtiques que formen un clade ben diferenciat, que agrupa la majoria d’espècies aquàtiques típiques del litoral marí, però en el que hi trobem també dues espècies d’aigua dolça, H. rheitrophila i H. scabra. Les espècies d’Hydropunctaria es poden reconèixer macroscòpicament, per les concrecions negre carbó que tenen en el taŀlus i que donen un aspecte papiŀlós a la superfície del liquen, un caràcter que no es presenta en la resta d’espècies de Verrucaria s.l. Hydropunctaria rheitrophila, al contrari d’H. scabra, és una espècie que ja ha estat citada prèviament per a la península Ibèrica (Pereira, 1992; Valcárcel & López de Silanes, 2011).
Material i mètode
Per a l’estudi dels diferents exemplars al microscopi òptic s’han fet seccions a mà alçada del taŀlus i dels ascomes, que s’han muntat en aigua o, per tal de millorar el contrast, en lactofenol-blau de cotó (LBC). Totes les mesures s’han pres en preparacions muntades en aigua. En l’expressió de les dimensions de les ascòspores s’inclou la llargada, l’amplada i la relació llargada/amplada, i es menciona per cada cas el valor mitjà en cursiva, enquadrat dels valors mínims i màxims que resulten de descartar el 10 % dels valors més alts i el 10 % dels més baixos, mentre que els valors extrems absoluts observats s’indiquen entre parèntesis. El nombre total d’ascòspores mesurades s’indica entre claudàtors [n]. Les fotografies de l’aspecte macroscòpic s’han pres amb una càmera Olympus® Tough 5 acollada al ocular de l’estereomicroscopi, i s’han integrat digitalment de forma automàtica amb la càmera o, en algun cas, amb ajuda del programa HeliconFocus® (Helicon Soft Inc.). Les fotografies dels caràcters microscòpics s’han pres igualment amb la mateixa càmera acollada al microscopi òptic, però en aquest cas sense procedir a la integració de les imatges.
Resultats
Hydropunctaria scabra (Vězda) C. Keller, Gueidan & Thüs (2009), Taxon 58: 194.
Verrucaria scabra Vězda (1970), Folia Geobot. Phytotax. 5(3-4): 308-309
Material estudiat
Catalunya, Lleida, Pallars Sobirà, Alt Àneu, riu de Cabanyeres, revolt de la pista passat el Plan d’Estaro, 42º36’21,48’’N, 1º03’51,72’’E, 1980 m, 20-VI-2007, P. Navarro-Rosinés (BCN-lich. 21879; 21880; 21881)
Descripció morfològica (Fig. 1a-e, i Fig. 2b)
Taŀlus crustaci, epilític, continu o només lleugerament fissurat, relativament prim, de 100-130 µm de gruix, de color verd molt fosc a negrós, que esdevé subgelatinós al hidratarse. La superfície taŀlina és escabrosa, recoberta d’abundants papiŀles punctiformes de color negre i d’uns 25-30 µm de diàmetre. En secció, el taŀlus en estructura està format per fileres d’hifes amb les cèŀlules orientades verticalment en tot el seu gruix, amb un còrtex paraplectenquimàtic, que no es
diferencia pràcticament en estructura de les hifes de la capa algal, i que presenta la paret de les cèŀlules de les capes més superficials del còrtex lleugerament pigmentades de verd. Capa algal amb les algues disposades també formant columnes verticals entre les hifes del fong. Meduŀla poc definida i no clarament visible, substituïda per una capa basal color bru fosc o negra, constituïda per cèŀlules isodiamètriques, arrodonides o lleugerament poligonals, de 3-6,5 µm de diàmetre. Per sobre d’aquesta capa basal, penetrant tant a zona hialina de la meduŀla com a la capa algal, s’observen abundants granulacions disperses, densament pigmentades, de color bru negrós, amb tonalitat similar a la de l’involucre que rodeja els peritecis. Aquestes granulacions pigmentades són les que donen forma a les papiŀles visibles en la superfície del taŀlus, especialment quant aquest està deshidratat.
Peritecis enfonsats en el taŀlus, de 180-230 µm de diàmetre, amb l’exciple o paret de l’ascoma incolor o de tonalitat bru clara, d’aproximadament 12-15 µm de gruix, que està envoltat externament per un involucre pigmentat, de color entre bru fosc i negre, que s’eixampla i se separa progressivament de l’exciple fins arribar al nivell de la base dels periteci, on acaba per unir-se amb la capa taŀlina basal pigmentada. Normalment, tot el conjunt, ascoma pròpiament dit més involucre, té un diàmetre de 0,3 a 0,6 mm, i queda en gran part recobert pel taŀlus, que es superposa a l’involucre, i dona lloc a engruiximents més o menys aparents en la superfície del liquen, on normalment els ascomes són només visibles per una zona ostiolar lleugerament concavada; però excepcionalment en algun cas, quan els ascomes estan situats a les parts més primes del taŀlus, l’involucre pot també estar exposat en part, al no quedar completament recobert pel taŀlus.
Ascs claviformes, de 52-64 × 20-25 µm, octosporats. Ascòspores simples, hialines, oblongo-eŀlipsoidals, de (14,5) 15-16,0-17 (18,5) × 7-8,0-9 µm, i amb una relació llargada/ amplada esporal de (1,8) 1,9-2,0-2,1 [n = 30].
Observacions i comentaris
Hydropunctaria scabra és una espècie que va ser descrita originalment com a Verrucaria scabra per Vězda (1970), de les muntanyes dels Carpats, i que no va ser inclosa en les claus d’identificació dels líquens d’Europa occidental de Clauzade & Roux (1985), les claus més utilitzades pels autors mediterranis, però sí que va ser tractada en algunes claus d’identificació incloses en treballs d’abast més local, com per exemple en les claus d’identificació dels líquens del sud-oest d’Alemanya (Wirth, 1980). En general podem considerar que, en molts territoris, H. scabra ha pogut passar desapercebuda en un concepte ampli d’H. rheitrophila, un tàxon aparentment més abundat i de distribució coneguda més àmplia. Ha estat necessari esperar a treballs més recents, per trobar una bona delimitació i comprensió d’Hydropunctaria scabra. Entre els treballs que han facilitat el reconeixement d’aquesta espècie, hi trobem l’estudi molecular de la família Verrucariaceae (Gueidan et al., 2009), treball on es descriu el gènere Hydropunctaria, i el treball de recopilació dels líquens aquàtics d’Europa central (Thüs & Schultz, 2009), un treball que inclou claus actualitzades per a la identificació dels líquens aquàtics d’aigua dolça, i que ha estat de gran aju-
120 Butlletí ICHN 85 (3), 2021
FLORA ET FAUNA C. ROMERO & P. NAVARRO-ROSINÉS
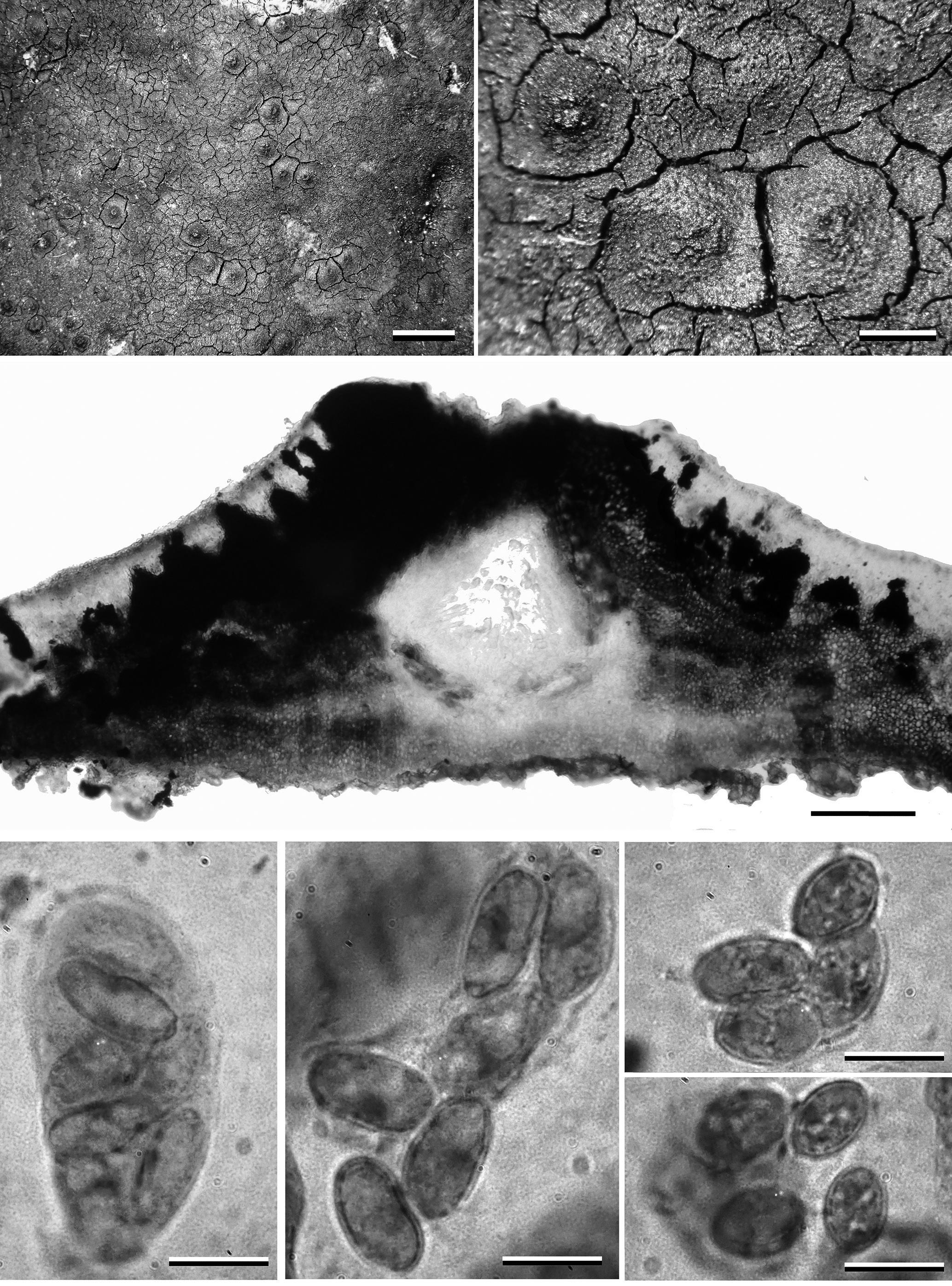
121 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
SOBRE LA PRESÈNCIA D'HYDROPUNCTARIA SCABRA A LA PENÍNSULA IBÈRICA
a b c d e f g
Figura 1. Hydropunctaria scabra (a-e): a) hàbit; b) detall ampliat del taŀlus on s’observa la zona ostiolar dels peritecis i l’aspecte papiŀlós de la superfície del taŀlus; c) secció d’un periteci, amb involucre, i d’una part del taŀlus, on s’hi observa la capa basal pigmentada i les concrecions de color negre carbó; d) asc octosporat; e) ascòspores. Hydropunctaria rheitrophila: (f-g) ascòspores. Escales: a= 2 mm, b= 0,5 mm. c= 100 µm, d-h= 10 µm.
Figura 2. Ascòspores de: a) Hydropunctaria scabra; b) H. rheitrophila. A la mateixa escala. per a comparació de la forma i la mida relativa entre les dues espècies. Escala = 10 µm.
da per a identificar de forma fiable la majoria de les espècies mencionades en el nostre treball.
Per descartar la possibilitat de què H. scabra hagi pogut passar desapercebuda en un concepte ampli d’H. rheitrophila en els estudis previs referits als líquens aquàtics de la Península Ibèrica (Pereira, 1992; Valcárcel & López de Silanes, 2011), hem revisat els exemplars d’aquesta última espècie estudiats en el seu moment per Pereira (1992) i actualment depositats en l’Herbari BCN (CeDocBiv, UB). Entre aquests exemplars revisats d’H. rheitrophila, que es mencionen més endavant en l’apartat de material estudiat, hi trobem tant exemplars precedents de Catalunya com d’altres localitats ibèriques. La revisió ens ha permès confirmar que tots aquests exemplars havien estat correctament identificats i pertanyen a H. rheitrophila.
Hydropunctaria rheitrophila i H. scabra són dues espècies d’aspecte macroscòpic molt semblant. Però, H. rheitrophila es caracteritza i diferencia d’H. scabra per tenir el taŀlus de color entre bru fosc i gairebé negrós, sense tonalitats verdoses, i per presentar la superfície del taŀlus més clivellada; tampoc presenta la capa basal negrosa que trobem a H. scabra, i el nombre de papiŀles visibles en el taŀlus és també molt més reduït. En relació a les característiques dels ascomes, al comparar les dues espècies, veiem que H. rheitrophila te peritecis d’estructura similar, però d’un diàmetre total, incloent l’involucre, inferior, de només 0,2-0,3 mm (en front dels 0,3-0,6 mm d’H. scabra). Els ascs són octosporats, amplament claviformes, d’aproximadament 40-50 × 14-19 µm, i les ascòspores simples, hialines, oblongo-arrodonides, de 11-12,5-14 (14,5) x (5,5) 6-7,0-7,5 (8) µm, amb una relació llargada-amplada de (1,4) 1,7-1,9-2 (2,1) [n = 58], unes dimensions que són clarament inferiors a les d’H. scabra (Fig. 2a i 3), i que constitueixen un bon caràcter diferencial entre les dues espècies. Totes les característiques esmentades per H. rheitrophila, coincideixen amb les indicades per altres autors (Smith et al., 2009; Thüs & Schultz, 2009; Orange, 2013).
Figura 3. Comparació mitjançant diagrames de caixa de les dimensions esporals d’Hydropunctaria rheitrophila i H. scabra. (A= amplada, L= llargada). Dimensions calculades en base als exemplars estudiats (H. rheitrophila: n = 58, i H. scabra: n = 30). La caixa està delimitada els valors compresos entre els percentils 10 i 90, el límit dels decils extrems s’indiquen fora de la caixa, la mitjana es representa per una línia horitzontal dintre la caixa. b a
Ecologia i distribució
Hydropunctaria scabra és una espècie saxícola fortament hidròfila, més o menys heliòfila i no nitròfila, que creix sobre roques silíciques inundades en zones de muntanya (Roux et coll., 2020). A la nova localitat aportada dels Pirineus catalans, ha estat recoŀlectada sobre roques completament submergides, en un rierol situat a l’estatge subalpí, a una altitud de 1980 m, sota l’ombra parcial d’una pineda. En aquesta localitat de les Planes de Son, H. scabra creix acompanyada de Verrucaria aquatilis, V. funckii i V. pachyderma. Aquestes mateixes espècies també son mencionades com acompanyats típiques d’H. scabra per Shivarov et al. (2017), en un tre-
122 Butlletí ICHN 85 (3), 2021
FLORA ET FAUNA
GEA,
C. ROMERO & P. NAVARRO-ROSINÉS
ball on es fa primera menció d’aquesta espècie per Bulgària. La distribució prèviament coneguda a Europa d’H. scabra es recollida per Shivarov et al. (2017), i es disposen de dades de: Àustria, Alemanya, Bulgària, Eslovàquia, França, Gran Bretanya, Islàndia, Montenegro, Polònia i Suïssa. Als Pirineus francesos hi tenim per a H. scabra diferents citacions relativament recents, en concret dels departaments dels Alts Pirineus (Coste, 2011) i dels Pirineus orientals (Roux et al., 2011), autors aquests últims que consideren la seva aportació com la primera menció d’aquesta espècie per França. En el departament dels Pirineus orientals, que inclou la Catalunya del Nord, H. scabra ha estat citada de diferents localitats de les reserves naturals de Prats de Molló, Jújols i Noedes, a les comarques del Conflent i del Capcir. Hydropunctaria scabra és considerada en el catàleg dels líquens de França com una espècie potencialment amenaçada (Roux et coll., 2020). Exemplars d’Hydropunctaria rheitrophila estudiats per comparació
Catalunya, Girona, Baixa Cerdanya, Meranges, emissari de l’Estany de Malniu, 2000-2200 m d’alt., granit, en roques completament submergides en llocs poc iŀluminats, 9-XI1985, I. Pereira (BCN-lich. 5549).– Girona, Baixa Cerdanya, Meranges, Coll superior, Puig Pedrós (pujada), 2300 m d’alt., granit, 13-X-1991, X. Llimona (BCN-lich. s. núm.).–Lleida, Vall d’Aran, Vielha, Güells del Joeu, 1400 m d’alt., granit i esquistos metamòrfics, en roques submergides en llocs poc iŀluminats, on creix juntament amb Hildenbrandia i Fissidens grandifrons, 7-VII-1989, I. Pereira (BCN-lich. 5550 i 5551).
andalusia, Granada, Trevélez, Sierra Nevada, Cañada de Siete Lagunas, 2800-2900 m d’alt., esquists metamòrfics, en roques a la vora d’un rierol, a 5 cm per sota el nivell de l’aigua, junt a Verrucaria funckii i V. aquatilis, 4-VII-1985, I. Pereira (BCN-lich. s. núm.).
Altres líquens aquàtics presents a Les Planes de Son i mata de València
A la mateixa localitat d’Hydropunctaria scabra, acompanyant aquesta espècie o sobre blocs diferents, es recoŀlectaren també altres espècies de líquens aquàtics que tampoc van ser citades en el treball dels líquens de les Planes de Son (Llop et al., 2010), però que són ja conegudes tant per a la península Ibérica com per a Catalunya. Per tal de completar el catàleg dels líquens de les Planes de Son, mencionem a continuació aquestes espècies aquàtiques i comentem la seva distribució tant al principat com al departament del Pirineus Orientals (Catalunya del Nord), on moltes d’aquestes espècies han estat també recentment citades de diferents localitats dels Pirineus per Roux et al. (2011).
Hydropunctaria rheitrophila (Zschacke) C. Keller, Gueidan & Thüs
Exemplars estudiats: BCN-lich. 21885a (junt amb Polyblastia sp. no identificada); 21962.
Els exemplars d’aquesta espècie recoŀlectats en aquesta localitat de les Planes de Son, estan poc desenvolupats i ocu-
pen petites extensions sobre les roques estudiades, pel que consideren que corresponen a taŀlus incipients. A Catalunya, H. rheitrophila és un tàxon que es coneix dels Pirineus (Baixa Cerdanya, Ripollès i Vall d’Aran) i també de diferents localitats del Montseny (Pereira, 1992).
Staurothele fissa (Taylor) Zwackh
Exemplars estudiats: BCN-lich. 21883.
És una espècie saxícola, preferentment silicícola, tot i que pot arribar també a créixer sobre substrats carbonatats, i que apareix en ambients que varien entre hidròfils i higròfils, tant sobre roques submergides com en roques emergents dels costats dels cursos d’aigua. A Catalunya ha estat citada per Pereira (1992) de diferents comarques dels Pirineus de Lleida (Pallars Jussà, Pallars Sobirà i Vall d’Aran), i de Girona (Ripollès), i també hi ha citacions per algunes comarques de Barcelona (Osona i Vallès Oriental). A Catalunya del Nord ha estat citada per Roux et al. (2011) de les comarques del Conflent i del Capcir, aportació que aquest autors consideren la primera menció d’aquesta espècie pel departament dels Pirineus Orientals.
Verrucaria aquatilis Mudd.
Exemplars estudiats: BCN-lich. 21875 (junt amb Verrucaria hydrela); 21876; 21877; 21878; 21881 (junt amb Hydropunctaria scabra).
Pel seu hàbitat, V. aquatilis és una espècie saxícola i silicícola, fortament hidròfila, que creix sobre roques submergides. És una espècie que no es coneixia prèviament del Pallars Sobirà, però sí de Catalunya. Hi ha citacions de les comarques dels Pirineus de Lleida (Vall d’Aran), i de Girona (Baixa Cerdanya i Alt Empordà). Tenim també citacions de Tarragona (Serra de Prades) i de Barcelona (Berguedà, la Selva i Vallès Oriental). Totes aquestes citacions prèvies han estat aportades per Pereira (1992). A Catalunya del Nord ha estat citada per Roux et al. (2011) de les comarques del Conflent i del Capcir. Al catàleg dels líquens de França, és considerada com una espècie potencialment amenaçada (Roux et coll., 2020).
Verrucaria funckii (Spreng.) Zahlbr.
Exemplars estudiats: BCN-lich. 21881 (junt amb Hydropunctaria scabra).
Espècie saxícola i silicícola, fortament hidròfila, que creix en superfícies planes i submergides de les roques. Totes les citacions prèvies de V. funckii per Catalunya es localitzen al nord del territori, a les comarques pirinenques, i han estat aportades per Pereira (1992). En concret hi ha dades de les comarques de Lleida (Vall d’Aran i Pallars Sobirà) i de Girona (Ripollès i Baixa Cerdanya). Roux et al. (2011) la citen també de Catalunya del Nord, a les comarques del Conflent i del Capcir. Al catàleg dels líquens de França, es considerada com una espècie poc comuna i potencialment amenaçada (Roux et coll., 2020).
Verrucaria hydrela Ach.
Exemplars estudiats: BCN-lich. 21875; 21883 (junt amb Staurothele fissa); 21884 (junt amb Polyblastia sp.).
123 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
LA PRESÈNCIA D'HYDROPUNCTARIA SCABRA A LA PENÍNSULA IBÈRICA
SOBRE
GEA, FLORA ET FAUNA
Cal considerar V. hydrela com una espècie saxícola, clarament hidròfila, que creix sobre roques silíciques que s’inunden periòdicament, en altituds molt àmplies, que van des d’ambients mesomediterranis fins pràcticament subalpins. A Catalunya a estat citada principalment de diferents localitats del Pirineus i del Montseny, però hi ha dades també de localitats de baixa altitud de la serra de Collserola. Es coneix de les comarques de Lleida (Vall d’Aran), de Girona (Alt Empordà) i de Barcelona (Berguedà, la Selva, Osona, Vallès Occidental i Oriental) (Hladun, 1985; Pereira 1992, Alvaro et al., 2009). A Catalunya del Nord ha estat citada també per Roux et al (2011) en diferents localitats de les comarques del Conflent i del Capcir.
Verrucaria margacea (Wahlenb.) Wahlenb.
Exemplars estudiats: BCN-lich. 21884 (junt amb Polyblastia sp.); 21963.
És una espècie saxícola, tant de substrats silícics com més o menys carbonatats, clarament hidròfila, que creix sobre roques permanentment submergides. Aquesta espècie, no coneguda prèviament del Pallars Sobirà, té una distribució irregular a Catalunya. Hi ha dades d’altres comarques pirinenques, en concret del Pallars Jussà, de la Vall d’Aran, de la Baixa Cerdanya i del Berguedà (Pereira, 1992). A estat també citada també per Roux et al. (2011) de la Catalunya del Nord, a les comarques del Conflent i del Capcir, on la mencionen per primera vegada en el departament dels Pirineus Orientals.
Verrucaria pachyderma Arnold
Exemplars estudiats: BCN-lich. 21881 (junt amb Hydropunctaria scabra); 21882.
És també una espècie hidròfila, pròpia de roques silíciques periòdicament submergides, granítiques en el nostre cas. Cal remarcar que la nostra és la segona localitat de V. pachyderma coneguda per a Catalunya, on només es coneixia prèviament d’una localitat de la Vall d’Aran, d’un rierol situat prop del Pla de Beret (Pereira, 1992). Al vessant nord del Pirineus ha estat citada per Roux et al. (2011), de les comarques del Conflent i del Capcir, autors que consideren que la seva menció és també la primera pel departament dels Pirineus Orientals.
Bibliografia
Álvaro, M. I., Barbero, M., Llop, E. & Gómez-Bolea, A. 2009. Riquesa briològica i liquenològica del Parc de Collserola (Catalunya). Estacions d’especial interès. Acta Botanica Barcinonensia, 52: 5-44.
Clauzade, G. & Roux, C. 1985. Likenoj de okcidenta Europo. Ilustrita determinlibro. Bulletin de la Société Botanique du Centre-Ouest, Nouvelle Série - Numéro Spécial: 7, Royan (France). 892 p.
Coste, C. 2011. Écologie et fuonctionnement des communautés lichéniques saxicoles-hydrophiles. Tesi doctoral. Université de Toulouse, França. 139 p.
Gueidan, C., Savic, S., Thüs, H., Roux, C., Keller, C., Tibell, L., Prieto, M., Heissmarsson, S., Breuss, O., Orange, A., Fröberg, L., Wynns, A. A., Navarro-Rosinés, P., Krzewicka, B., Pykälä, J., Grube, M. & Lutzoni, F. 2009. Generic classification of the Verrucariaceae (Ascomycota) base on molecular and morphological evidence: recent progress and remaining challenges. Taxon, 58 (1): 184-208.
Hladun, N. L. 1985. Aportació a la flora, morfologia i vegetació dels líquens de la part alta del Montseny. Institut d’Estudis Catalans. Arxius de la secció de ciències, LXXX. 204 p.
Llop, E., Barbero, M., Chesa, M. J., Fernández-Brime, S., Figueras, G., Gómez-Bolea, A., Hladun, N. L., Muñiz, D. & NavarroRosinés, P. 2010. Els líquens i els fongs liquenícoles de Les Planes de Son i mata de València. In: Germain, J. (cur.). Els sistemes naturals de Les Planes de Son i la mata de València. Treballs de la Institució Catalana d’Història Natural, 16: 189-227.
Orange, A. 2012. Semi-cryptic marine species of Hydropunctaria (Verrucariaceae, lichenized Ascomycota) from north-west Europe. The Lichenologist, 44 (3): 299-320.
Orange, A. 2013. British and other pyrenocarpous lichens. National Musem Wales. Treball inèdit, 217 p. Disponible en: https:// museum.wales/media/13849/Orange-A-2013-British-and-otherpyrenocarpous-lichens.pdf
Pereira, I. 1992. Flora, vegetación y ecología de los líquenes acuáticos de España. Tesis Doctoral. Universitat de Barcelona. 342 p.
Roux, C. et coll. 2020. Catalogue des lichens et champignons lichénicoles de France métropolitaine. 3e édition revue et augmentée (2020). Ed. Association française de lichénologie (AFL), Fontainebleau, França. 1769 p.
Roux, C., Masson D., Bricaud, O., Coste, C. & Poumarat, S. 2011. Flore et végétation des lichens et champignons lichénicoles de quatre réserves naturelles des Pyrénées-Orientales (France). Bulletin de la Société Linnéenne de Provence, nº special 14: 3-152.
Shivarov, V. V., Thüs, H. & Denchev, C. M. 2017. First records of two freshwater lichens, Hydropunctaria scabra and Verrucaria alpicola, from Bulgaria. Mycobiota, 7: 1-5.
Smith, C. W., Aptroot, A., Coppins, B. J., Fletcher, A., Gilbert, O. L., James, P. W. & Wolseley, P. A. (eds). 2009. The Lichens of Great Britain and Ireland 2nd edition. Ed. British Lichen Society, London. 1046 p.
Thüs, H. & Schultz, M. 2009. Freshwater Flora of Central Europe. Vol. 21/1, Fungi, Part 1, Lichens. Spektrum, Heidelberg. 217 p.
Valcárcel, C. P. & López de Silanes, M. E. 2011. Especies acuáticas del género Verrucaria s.l. (Ascomycetes liquenizados) en la península Ibérica. Nova Acta Científica Compostelana (Bioloxía), 19: 5-20.
Vězda, A. 1970. Neue und wenig bekannte Flechten in der Tschechoslowakei. I. Folia Geobotanica et Phytotaxonomica 5(3-4):307-337
Wirth W. 1980. Flechtenflora. Ökologische Kennzeichnung und Bestimmung der Flechten Südwestdeutschlands und angrenzender Gebiete. Ulmer, Stuttgar. 552 p.
124 Butlletí ICHN 85 (3), 2021
C. ROMERO & P. NAVARRO-ROSINÉS
Butlletí de la Institució Catalana d’Història Natural, 85 (3): 125-126. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition)
NOTA BREU
NOTA BREU
Noves dades poblacionals sobre Daphne alpina (Thymelaeaceae) a Catalunya
New populational data about Daphne alpina (Thymelaeaceae) in Catalonia
Pere Aymerich* & Albert Tarragó**
* C. Barcelona, 29. 08600 Berga.
** C. del Sol, 5. 25280 Solsona.
Autor para la correspondència: Pere Aymerich. A/e: pere_aymerich@yahoo.es
Rebut: 05.09.2021. Acceptat: 07.09.2021. Publicat: 30.09.2021
Daphne alpina L. (Timeleàcies) és un arbust (Figs. 1 i 2) que viu a les muntanyes del sud d’Europa, especialment a la seva part oriental, i que té als Pirineus un extrem oest d’àrea. En aquesta serralada es coneix d’un nombre baix de localitats, al voltant de deu, que es concentren bàsicament al massís de les Corberes i en l’àmbit del Cadí-Moixeró. A conseqüència de la seva gran raresa, al Llibre Vermell de la flora vascular de Catalunya (Sáez et al., 2010) va ser qualificada com a espècie amenaçada, amb la categoria EN (En perill). Pel mateix motiu ha estat protegida legalment, com a resultat de la seva incorporació als catàlegs legals d’espècies amenaçades català (Decret 172/2008) i espanyol (Ordre AAA/1771/2015 de modificació del Reial Decret 139/2011), en tots dos casos amb la categoria de màxima protecció (En perill d’extinció); cal dir que, a escala estatal, només es coneix de Catalunya.
A Catalunya es va descobrir per primera vegada els anys 1970, a la vall de Pi, Cerdanya (Gruber, 1977). Aquesta va ser durant molt de temps l’única localitat coneguda, i la qualificació com a EN a Sáez et al., (2010) es fonamentava en les poques dades disponibles sobre aquella població, un sol cens de 71 individus, dels quals 65 reproductors (Aymerich & Sáez, 2001). En la darrera dècada s’han anat trobant altres

localitats a la Cerdanya (Aymerich, 2014, 2021) i una a l’Alt Urgell (Guardiola et al., 2016), i en l’actualitat sumen un total de cinc (Fig. 3). Tot i això, en l’actualització recent de la Llista Vermella de la flora catalana (Aymerich & Sáez, 2021) s’ha mantingut per aquesta espècie la mateixa categoria (EN) i el mateix criteri (D: població de menys de 250 individus madurs).
Atès que es tracta d’una espècie protegida d’interès conservacionista alt i que les dades publicades sobre la mida poblacional de les diverses localitats són molt escasses, s’ha considerat convenient sintetitzar en aquesta nota les informacions poblacionals més actualitzades, que són les que justifiquen el manteniment de la categoria EN a la Llista Vermella. Es tracta de dades obtingudes en censos promoguts pel Parc Natural del Cadí-Moixeró (localitats de la Cerdanya) o per la Reserva Nacional de Caça del Boumort (serra de Prada, Alt Urgell). Aquestes dades es mostren a la Taula 1. Com es pot observar a la taula, sumant totes les localitats el nombre d’individus reproductors és de 132, ben per sota del límit de 250 establert per a la categoria EN. Cal precisar que és probable que el nombre real sigui una mica més alt, però en tot cas, clarament inferior a aquesta xifra límit. Això és degut a dos factors: a) La població de la serra de Prada es

125 Butlletí ICHN 85 (3), 2021
DOI: 10.2436.20.1502.01.83
Figura 1. Individu de D. alpina no reproductor en una tartera estabilitzada, serra de Prada (A. Tarragó)
Figura 2. Daphne alpina: Detall de fulles i flors, vall de Pi (P. Aymerich)
Taula 1. Censos més recents disponibles de cada localitat.
Localitat Serrat de les Pedrusques Roca d’Urús Canals Vall de Pi Serra de Prada TOTAL Any
va censar quan les plantes estaven fructificades, de manera que no es pot excloure que no es comptés algun individu reproductor que no va arribar a desenvolupar fruits; i b) Les poblacions del serrat de les Pedrusques i de la Roca d’Urús es fan en cingles molt intricats, de prospecció difícil fins i tot utilitzant òptiques per observar a distància, motiu pel qual és molt probable que el nombre de plantes existent sigui més alt que el reflectit pels censos.
L’única localitat de la qual es tenen censos clarament separats en el temps, i que permetin avaluar la dinàmica poblacional, és la de la vall de Pi. En aquesta població, en el període 1999-2009, el nombre de reproductors es va mantenir en general estable (65 i 69 respectivament) i el nombre de no reproductors va augmentar (6 i 16). En conjunt, la tendència va ser positiva, amb un increment del 19,7 %. Tot i algunes limitacions metodològiques que afecten la detecció dels individus no reproductors, aquesta tendència positiva es considera real, ja que es va detectar en els quatre sectors diferenciats en l’àmbit del cens.
En part dels censos de les poblacions de D. alpina s’han inclòs dades referides a la fructificació, perquè ja en les primeres avaluacions que es van fer a la vall de Pi (Aymerich & Sáez, 2001) es va considerar sorprenent que, malgrat la taxa de fructificació alta i la gran disponibilitat d’hàbitats potencials, el reclutament fos molt baix. En aquesta localitat, en anys posteriors s’ha observat que tot i que hi ha anys en què la fructificació efectiva és alta (ex. 2009 o 2015), en altres
(ex. 2014) és pràcticament nul·la, ja que els fruits no maduren ni desenvolupen llavors viables. A la localitat de la serra de Prada, l’any 2017 es va quantificar la fructificació, amb un resultat d’un total de 935 fruits i una mitjana de 30,1 fruits per planta, que és relativament baixa.
Referències
Aymerich, P. 2014. Notes florístiques de les conques altes dels rius
Segre i Llobregat (II). Orsis, 28: 7-47.
Aymerich, P. 2021. Notes florístiques de les conques altes dels rius
Segre i Llobregat (V). Butlletí de la Institució Catalana d’Història Natural, 85: 41-53.
Aymerich, P. & Sáez, L. 2001. Dades sobre l’estatus d’algunes plantes endèmiques, amenaçades o rares a Catalunya (nord-est de la península Ibèrica). Orsis, 16: 47-70.
Aymerich, P. & Sáez, L. 2021. Llista Vermella de la flora vascular de Catalunya. Actualització 2020. Monografies de la Institució Catalana d’Història Natural, 2. 100 p.
Gruber, M. 1977. Contribution à la flore des Pyrénées ariègeoises et catalanes: 2e note. Bulletin Societé Botanique de France, 124: 93-104.
Guardiola, M., Petit, A., Molero, J. & Sáez, L. 2016. Aportacions al coneixement de la flora vascular del massís de Boumort i serres veïnes. Orsis, 30: 67-100.
Sáez, L., Aymerich, P. & Blanché, C. 2010. Llibre Vermell de les plantes vasculars endèmiques i amenaçades de Catalunya. Argania Editio. Barcelona. 811 p.
126 Butlletí ICHN 85 (3), 2021 NOTA BREU P. AYMERICH & A. TARRAGÓ
cens
2021 2014-2018 2009 2017 Individus reproductors 27 4 1 69 31 132 Individus no reproductors 15 7 0 16 43 81 Suma individus 42 11 1 85 74 213
2017-2018
Figura 3. Distribució actualment coneguda de D. alpina a Catalunya.
Butlletí de la Institució Catalana d’Història Natural, 85 (3): 127-129. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print
NOTA BREU
Primer hospedador conocido de Aegilips clarimontis Kieffer, 1907 (Figitidae: Anacharitinae)
First known host of Aegilips clarimontis Kieffer, 1907 (Figitidae: Anacharitinae)
Juli Pujade-Villar*, Juan M. Vanegas-Rico** & Noel Mata-Casanova**
* Universitat de Barcelona. Facultat de Biologia. Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals. Avda. Diagonal 645. 08028 Barcelona, Cataluña. A/e: jpujade@ub.edu, feofitotu@gmail.com
** Universidad Nacional Autónoma de México. FES-I, Control de plagas. Unidad de morfología y función. Los Reyes Iztacala. 54090 Tlalnepantla, Estado de México, México. A/e: entomologo.mexicano@gmail.com
Autor para la correspondencia: Noel Mata-Casanova. A/e: feofitotu@gmail.com
Rebut: 01.09.2021. Acceptat: 13.09.2021. Publicat: 30.09.2021
La subfamilia Anacharitinae puede distinguirse con facilidad de los otros Figitidae por tres sinapomorfias principales (Ros-Farré et al., 2000): 1) cabeza de forma triangular en visión anterior (aunque en algunos ejemplares una expansión secundaria del área malar lo hace más cuadrada), 2) placa pronotal redondeada y continua, y 3) mandíbulas pequeñas, ampliamente superpuestas. Biológicamente, también son distintos al resto de subfamilias pues son los únicos que atacan larvas de Neuroptera.
Los Anacharitinae incluyen nueve géneros (Mata-Casanova et al., 2019): Acanthaegilips Ashmead, 1897; Acantahegilopsis Pujade-Villar, 2013; Aegilips Haliday, 1835; Anacharis Dalman, 1823; Calofigites Kieffer, 1909; Hexacharis Kieffer, 1907; Proanacharis Kovalev, 1996; Solenofigites Díaz, 1979 y Xyalaspis Hartig, 1843. Seis géneros están presentes en México (Mata-Casanova et al., 2013) de los cuales tres son endémicos de la región neotropical: Acanthaegilips, Calofigites y Solenofigites
La biología de la mayoría de especies de Anacharitinae es desconocida. Según Ronquist (1999) se han citado atacando larvas depredadoras de las familias Hemerobiidae y Chrysopidae; no obstante, su presencia en esta última familia de Neuroptera es dudosa, ya que toda la bibliografía encontrada menciona sólo a Hemerobiidae (Díaz, 1979; New, 1979; Kierych, 1984; Miller & Lambdin, 1985; Ferguson, 1986; Cave & Miller, 1987; Mata-Casanova et al., 2014, Mata-Casanova, 2018), y ninguna refiere a hospederos de la familia Crhysopidae.
Los Hemerobiidae se caracterizan ser depredadores voraces tanto en el estado larval como en estado adulto de diferentes artrópodos de cuerpo blando como: áfidos y otros homópteros, hemípteros fitófagos, larvas de lepidópteros, por lo que son utilizadas en programas de control de plagas en la agricultura (New, 1975).
Los huéspedes conocidos de Hemerobiidae atacados por Anacharitinae son: Drepanacra Tillyard, 1916 (Drepanacrinae), Hemerobius Linnaeus, 1758 y Wesmaelius Krüger,
1922 (Hemerobiinae), Micromus Rambur, 1842 (Microminae), Notiobiella Banks, 1909 (Notiobiellinae) y Sympherobius Banks, 1904 (Sympherobiinae).
Aegilips agrupa in total de 22 especies (Mata-Casanova, 2018), de las cuales sólo de las siguientes es conocido algún hospedador siempre perteneciente a los género Wesmaelius o Sympherobius: A. atricornis Ferguson, 1985 especie de distribución Holártica e Hindomalaya (W. betulinus (Ström, 1788)) (Ferguson, 1985), A. capensis de distribución Afrotropical (un Hemenobiidae no determinado) (Mata-Casanova et al., 2017), A. chilensis Bréthes, 1918 de distribución andina (una especie no determinada de Sympherobius) (Loiacono & Díaz, 1977), y A. nitidulus de distribución paleártica (W. betulinus (Strøm, 1788) y W. subnebulosus (Stephens, 1836)) (Ferguson, 1985).
En un estudio sobre los organismos que atacan el nopal (Opuntia ficus-indica (L.) Mill., 1768) realizado por el segundo de los autores, se colectaron larvas de neuróptero que estaban depredando a Dactylopius opuntiae (Cockerell, 1896) (Hemiptera Coccoidea) en Colpos (Texcoco, México) de las cuales el 15.ix.2013 emergió una hembra de A. clarimontis Los adultos obtenidos del neuróptero (Fig. 1) corresponden a Sympherobius nr angustus (Banks, 1904).
En México se conocen dos especies depredadoras de D. opuntiae en nopal: Sympherobius barberi (Banks, 1903) y S angustus (Banks, 1904) (Vanegas-Rico et al., 2010), ambas corroboradas por J. Oswald (Department of Entomology, Texas A & M University, EEUU), a pesar de que Montserrat et al. (2018) indica que S. barberi se asocia Opuntia Mill.y S. angustus en Picea A.Dietr.
En cultivos de nopal de Tlalnepantla (Morelos), durante 2008- 2012, más del 99 % de los hemeróbidos que atacan cochinillas del nopal correspondían a Sympherobius barberi (Vanegas-Rico, datos no publicados). En contraste, en cultivos experimentales de Colpos, (Estado de México), durante el periodo 2010-2014, el neuróptero colectado en el nopal presentaba características del grupo «angustus», ya que las
127 Butlletí ICHN 85 (3), 2021
BREU
NOTA
edition)
10.2436.20.1502.01.84
DOI:

características coincidían con las de los ejemplares determinados por Jhon Olwald como S. angustus para Tlalnepantla. No obstante existía la duda en si eran S. angustus o S. bifasciatus Los ejemplares obtenidos se revisaron nuevamente en 2019, llegando a la conclusión de que se acercaban más a S. angustus. Posteriormente, se envió material al especialista J. Oswald, sin obtener confirmación. Por otro lado, Montserrat (2008) menciona que S. angustus es una especie de morfología muy variable habiendo estudiado numerosos ejemplares con una gran variabilidad en su coloración y venación alar, solapándose con otras especies conocidas del grupo ‘angustus’ (Sympherobius angustus, S. quadricuspis, S. bifasciatus, S. californicus y S. constrictus). En cualquier caso, el material examinado no corresponde a S. barberi, y lo denominamos Sympherobius nr angustus por lo mencionado anteriormente.
Esta cita representa no solo el primer huésped conocido de A. clarimontis sino también la segunda cita Aegilips en Sympherobius y la primera especie de hospedador para una especie de Aegilips de América.
Atendiendo a los escasos resultados biológicos de Aegilips, las especies americanas atacan hasta el momento el género Sympherobius y las especies paleárticas al género Wesmaelius
Bibliografía
Cave, R. D. Miller, G. L. 1987. Notes on Anacharis melanoneura (Hymenoptera: Figitidae) and Charitopes mellicornis (Hyme-
noptera: Ichneumonidae) parasitizing Micromus posticus (Neuroptera: Hemerobiidae). Entomological News, 98: 211-216.
Díaz, N. B. 1979. Neotropical Parasitic Hymenoptera. Revista de la Sociedad Entomológica Argentina, 38 (1-4): 21-28.
Fergusson, N. D. M. 1985. British species of the parasitic cynipidwasp genus Aegilips (Hymenoptera: Cynipoidea, Anacharitinae). Journal of Natural History, 19 (4): 811-818.
Fergusson, N. D. M. 1986. Charipidae, Ibaliidae and Figitidae (Hymenoptera: Cynipoidea). Handbooks for Identification of British Insects, Vol. 8, Part 1c: 29 p.
Kierych, E. 1984. Notes on the genus Prosynapsis D. T. et Kieff. (Synapsis Först) with a list of Anacharis Dalm. species occurring in Poland (Hymenoptera, Cynipoidea, Anacharitidae). Annales Zoologici, 37 (11): 335-339.
Loiacono, M. S. & Díaz, N. B. 1977. Anotaciones sobre himenópteros procotrupoideos y cinipoideos argentinos (Hymenoptera: Proctotrupoidea, Cynipoidea). Neotrópica, 23 (69): 95-102.
Mata-Casanova, N., Selfa, J. & Pujade-Villar, J. 2013. Anacharitinos neotropicales (Hymenoptera: Figitidae) con especial atención a las especies mexicanas. Entomología mexicana, 12 (2): 1482-1487.
Mata-Casanova, N., Selfa, J., Arcaya, E. & Pujade-Villar, J. 2014. First host record for Acanthaegilips (Hymenoptera: Figitidae: Anacharitinae) and description of a new species from Venezuela. Florida Entomologist, 97: 461-464.
Mata-Casanova, N., Selfa, J. & Pujade-Villar, J. 2017. Revision of the current knowledge of genus Aegilips Haliday, 1835 (Hymenoptera: Figitidae: Anacharitinae) in the Afrotropical region, with description of three new species. African Entomology, 25(1): 62-71. http://dx.doi.org/10.4001/003.025.0062
Mata-Casanova, N., Selfa, J., Gallardo, F. E. & Pujade-Villar, J. 2019. Aegilips chilensis (Hymenoptera: Cynipoidea: Figitidae:
128 Butlletí ICHN 85 (3), 2021 NOTA BREU J. PUJADE-VILLAR ET AL
Figura 1. Adulto Hemerobiidae (Sympherobius nr angustus) emergido a partir de larvas de neuróptero aisladas que depredaban la colonia de Dactylopius opuntiae (Coccoidea) en Colpos sobre nopal.
Anacharitinae): redescription and biogeographic considerations. Neotropical Entomology, 48 (1-4):1-5. DOI: 10.1007/s13744019-00674-9
Mata-Casanova, N. 2018. Taxonomical revision of subfamily Anacharitinae (Hymenoptera: Cynipoidea: Figitidae). Ph. D. Thesis, Department of Zoology, University of Barcelona, Barcelona, 535 p. [Disponible en: http://hdl.handle.net/2445/123302]
Miller, G. L. & Lambdin, P. L. 1985. Observations on Anacharis melanoneura (Hymenoptera: Figitidae), a parasite of Hemerobius stigma (Neuroptera: Hemerobiidae). Entomological News, Philadelphia, 96: 93-97.
Montserrat, V. J., 2008. Nuevos datos sobre algunas especies de Hemeróbidos (Insecta, Neuroptera, Hemerobiidae). Graellsia, 64 (2): 233-253.
New, T.R., 1975. The biology of Chrysopidae and Hemerobiidae (Neuroptera), with reference to their usage as biocontrol agents: a
review. Ecological Entomology, 127 (2): 115-140.
New, T. R. 1979. An Australian species of Xyalaspis Hartig (Hymenoptera: Figitidae). Journal of the Australian Entomological Society, 18: 177-180. https://doi.org/10.1111/j.13652311.1975.tb00561.x|
Ronquist, F. 1999. Phylogeny, classification and evolution of the Cynipoidea. Zoologica Scripta, 28 (1–2): 139–164.
Ros-Farré, P., Ronquist, F. & Pujade-Villar, J. 2000. Redescription of Acanthaegilips Ashmead 1987, with characterization of the Anacharitinae and Aspiceratinae (Hymenoptera: Cynipoidea: Figitidae). Zoological Journal of the Linnean Society, 129: 467-488.
Vanegas-Rico, J. M., Lomeli-Flores, J. R., Rodríguez-Leyva, E., Mora-Aguilera, G., & Valdez, J. M. 2010. Enemigos naturales de Dactylopius opuntiae (Cockerell) en Opuntia ficus-indica (L.) Miller en el centro de México. Acta zoológica mexicana, 26 (2): 415-433.
129 Butlletí ICHN 85 (3), 2021
BREU PRIMER HOSPEDADOR CONOCIDO DE AEGILIPS CLARIMONTIS KIEFFER, 1907 (FIGITIDAE: ANACHARITINAE)
NOTA
Nuevos datos sistemáticos y de distribución de Xyletinus (Calypterus) bucephalus s. lat. (Coleoptera: Ptinidae: Xyletininae)
Amador Viñolas* & Josep Muñoz-Batet*
* Museu de Ciències Naturals de Barcelona. Laboratori de Natura. Coŀlecció d’artròpodes. Passeig Picasso, s/n. 08003 Barcelona.
Autor para la correspondencia: Amador Viñolas. A/e: av.rodama@gmail.com
Rebut: 25.08.2021; Acceptat: 14.09.2021; Publicat: 30.09.2021
Resumen
Xyletinus (Calypterus) bucephalus s. lat. es una especie de Xyletininae con una distribución paleártica muy amplia y estrechamente ligada a zonas semiáridas y excrementos de ganado ovino, caprino y equino. Especie que, como todas las que tienen un área de distribución tan extensa, presenta una cierta variabilidad individual, que no tiene ninguna validez especifica. Dicha variabilidad ha posibilitado la descripción de una subespecie y de dos nuevas especies. Tras el estudio de 208 especímenes de prácticamente toda el área de distribución se ha podido constatar que no hay diferencias especificas válidas para su separación y que el edeago con una pieza muy bien caracterizada en el saco interno, no varia en todos los ejemplares examinados por lo que se pasa a sinonímia de Xyletinus (Calypterus) bucephalus los siguientes taxones: Xyletinus (Calypterus) bucephalus theanus Reitter, 1901 n. syn., Xyletinus (Calypterus) bucephalus omanus Toskina, 1998 n. syn., Xyletinus (Calypterus) materliki Toskina, 2002 n. syn. y Xyletinus (Calypterus) lepidus Toskina, 2006 nov. syn. Se representa el habitus de varios de los ejemplares estudiados, sus edeagos y los dibujados por diferentes autores así como el mapa de distribución paleártica de la especie.
Palabras clave: Coleoptera, Ptinidae, Xyletininae, Xyletinus (Calypterus) bucephalus s. lat., sistemática, distribución.
Abstract
New systematic and distribution data for Xyletinus (Calypterus) bucephalus s. lat. (Coleoptera: Ptinidae: Xyletininae)
Xyletinus (Calypterus) bucephalus s. lat. is a species of Xyletininae with a very wide Palaearctic distribution and closely related to semi-arid areas and with sheep, goats and horses faeces. Species that have such an extensive distribution area, present always a certain individual variability, which has no specific validity. Such variability have made possible describe one subspecies and two new species.
After the study of 208 specimens from practically the entire distribution area, it has been possible to verify that there are no specific valid differences for their separation and that the aedeagus, with a very well characterized piece in the internal sac, does not vary in all the studied specimens, so the following taxa are synonymized with Xyletinus (Calypterus) bucephalus: Xyletinus (Calypterus) bucephalus theanus Reitter, 1901 n. syn., Xyletinus (Calypterus) bucephalus omanus Toskina n. syn., 1998, Xyletinus (Calypterus) materliki Toskina, 2002 n. syn. and Xyletinus (Calypterus) lepidus Toskina, 2006 n. syn. The habitus of several of the specimens studied, their aedeagus and those drawn by different authors as well as the Palaearctic distribution of the species are represented.
Key word: Coleoptera, Ptinidae, Xyletininae, Xyletinus (Calypterus) bucephalus s. lat., systematic, distribution.
Resum
Noves dades sistemàtiques i de distribució de Xyletinus (Calypterus) bucephalus s. lat. (Coleoptera: Ptinidae: Xyletininae)
Xyletinus (Calypterus) bucephalus s. lat. és una espècie de Xyletininae amb una distribució paleàrtica molt àmplia i estretament lligada a zones semiàrides i a excrements de bestiar oví, caprí i equí. Espècie que, com totes les que tenen una àrea de distribució tan extensa, presenta una certa variabilitat individual, que no té cap validesa específica. Aquesta variabilitat ha possibilitat la descripció d’una subespècie i de dues noves espècies. Després de l’estudi de 208 espècimens de pràcticament tota l’àrea de distribució s’ha pogut constatar que no hi ha diferències especifiques vàlides per a la seva separació i que el edeagus, amb una peça molt ben caracteritzada en el sac intern, no varia en tots els exemplars examinats de manera que es passa a sinonímia de Xyletinus (Calypterus) bucephalus els següents taxons: Xyletinus (Calypterus) bucephalus theanus Reitter, 1901 n. syn., Xyletinus (Calypterus) bucephalus omanus Toskina, 1998 n. syn., Xyletinus (Calypterus) materliki Toskina, 2002 n. syn. i Xyletinus (Calypterus) lepidus Toskina, 2006 n. syn. Es representa l’habitus de diversos exemplars estudiats, els seus edeagus i els dibuixats per diferents autors així com el mapa de distribució de l’espècie.
Paraules clau: Coleoptera, Ptinidae, Xyletininae, Xyletinus (Calypterus) bucephalus s. lat., sistemàtica, distribució paleàrtica.
urn:lsid:zoobank.org:pub:E903B1CC-8458-44F0-92A8-5DA232360F46
Butlletí de la Institució Catalana d’Història Natural, 85 (3): 131-138. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition) DOI:
131 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
GEA, FLORA ET FAUNA
10.2436.20.1502.01.85
GEA,
Introducción
Xyletinus (Calypterus) bucephalus s. lat. es una especie del área paleártica conocida de la zona mediterránea europea, africana y Asia Menor, alcanzando la Península Arábiga y citada también del norte de la región afrotropical (Viñolas, 2020; Zahradník, 2007). Su distribución está ligada a las zonas semiáridas con gran presencia de ganado bovino, caprino o equino, ya que las larvas se desarrollan en los excrementos secos de los citados herbívoros (Viñolas, 2020).
Se estudia la validez de las tres subespecies de X. (C.) bucephalus actualmente existentes, así como la validez de otras dos especies recientemente descritas de Túnez y Yemen, que nos parecen indiferenciables especificamente de X. (C.) bucephalus.
Se representa el habitus de diferentes ejemplares de la especie, así como las diferentes representaciones del edeago de X. (C.) bucephalus s. lat. realizadas por diferentes autores, incluyéndose también un mapa con la distribución de la especie.
Material y métodos
Los especímenes estudiados estaban montados sobre etiquetas entomológicas y la mayoría de ejemplares fueron re-
colectados mediante diferentes metodologías de trampeo, en especial mediante luz ultravioleta. En los ejemplares en los que se ha realizado el estudio del edeago, tras un procedimiento de despegado y ablandamiento se procedió a la extracción del órgano, que se ha montado en preparación microscópica sobre una lámina de poliestireno transparente de la marca Evergreen®, con líquido DMHF. Todos los ejemplares se han montado otra vez en seco sobre etiquetas entomológicas. Las fotografías se han realizado con una cámara Canon® model EOS 760D, con objetivo de microscopía mediante el método de capas, con tratamiento de las imágenes obtenidas mediante el programa Zerene Stacker®. Los dibujos se han realizado con el programa de Adobe® Ilustrator CS5, con la obtención de archivos PostScript® 3™.
En los ejemplares referenciados con el acrónimo «MZB» el número corresponde al registro del espécimen en la colección del Museu de Ciències Naturals de Barcelona.
Resultados
Hasta hace pocos años solo se conocían dos subespecies, X. (C.) bucephalus bucephalus (Illiger, 1807) (Fig. 1) propia del Mediterráneo central y occidental, X. (C.) bucephalus theanus Reitter, 1901 (Fig. 2) de la parte oriental, éste último descrito con material procedente de la isla de Andros

132 Butlletí ICHN 85 (3), 2021
FLORA
A. VIÑOLAS & J. MUÑOZ-BATET
ET FAUNA
1 2
Figuras 1-2. Habitus de: 1) Xyletinus (Calypterus) bucephalus bucephalus (Illiger, 1807), de Tahla, Taza, Marruecos; 2) X. (C.) bucephalus theanus Reitter, 1901, del Mar Muerto, Jordania. Escala = 1,5 mm.
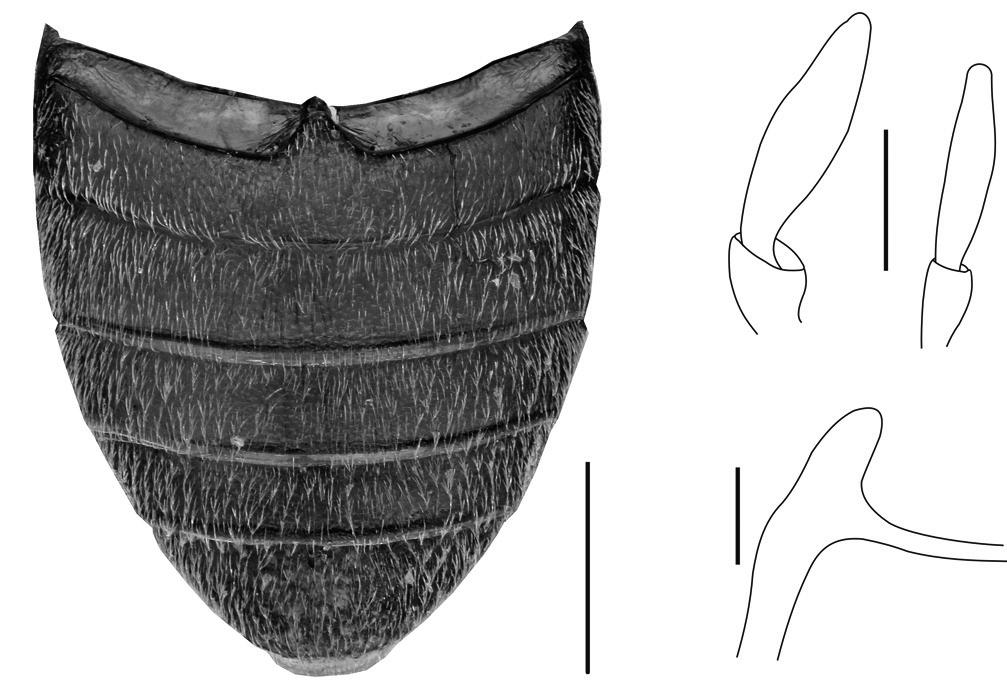
una raza del mismo». Hemos podido estudiar 43 ejemplares asignados a la subespecie procedentes del territorio Palestino, Jordania y Arabia Saudí y en ellos no hemos podido encontrar ninguna diferencia externa ni en la configuración del edeago que sirva para separarlo de X. (C.) bucepahlus bucephalus. Como se puede observar en las figuras 7 y 8, en el saco interno siempre aparece la misma pieza falciforme, grande y muy característica, ya claramente representada por Español (1979b) en el dibujo del saco evaginado de un ejemplar de la Arabia Saudí (Fig. 11).
Español (1979b) en el estudio de los Anobiinae de la Arabia Saudí sitúa los ejemplares como X. (C.) bucephalus bucephalus, mientras que Zahradník (1988) en el estudio de los Ptinidae de Turquía sitúa los ejemplares en la subespecie X. (C.) bucephalus theanus, indicando que esta es propia del Mediterráneo oriental. Zahradník (2007) en el catálogo paleártico da la siguiente distribución para la subespecie theanus: Bulgaria, Egipto, Grecia, Israel, Siria y Turquía, curiosa distribución, ya que el área de la subespecie divide en dos la distribución de X. (C.) bucephalus bucephalus.
(islas Cícladas, Grecia) como especie válida que fue pasada posteriormente a subespecie de X. (C.) bucephalus (Zaradik, 2007). En la descripción original, Reitter (1901) sólo la separa en la clave de X. (C.) bucephalus por el colorido del cuerpo e indica «Por lo demás similar al anterior y posiblemente
Así pues, se propone que Xyletinus (Calypterus) bucephalus theanus Reitter, 1901 debe pasar a n. syn. de Xyletinus (Calypterus) bucephalus (Illiger, 1807).
Bercedo & Arnáiz (2008) pasaron a sinonimia de X. (C.) bucephalus el Xyletinus (Calypterus) wollastoni Gottwald,
133 Butlletí ICHN 85 (3), 2021
FLORA ET FAUNA NUEVOS DATOS SISTEMÁTICOS Y DE DISTRIBUCIÓN DE XYLETINUS (CALYPTERUS) BUCEPHALUS S. LAT.
GEA,
6
Figuras 3-6. Xyletinus (Calypterus) bucephalus bucephalus: 3) abdomen; 4) último artejo de los palpos maxilares; 5) último artejo de los palpos labiales; 6) parte anterior del segmento genital. Escalas a = 0,3 mm; b = 0,1 mm.
54 3
8 7
Figuras 7-8. Edeago de: 7) Xyletinus (Calypterus) bucephalus bucephalus de Marruecos; 8) X. (C.) bucephalus theanus de Jordania. Escala = 0,5 mm.
GEA, FLORA ET FAUNA
Figuras 9-14, 9) Edeago de Xyletinus (Calypterus) bucephalus bucephalus: de Marruecos, según Español (Mediterránia, 3. 1979); 10) de Arabia Saudita, según Español (Fauna of Saudi Arabia, 1. 1979). 11) Saco interno del edeago de X. (C.) bucephalus bucephalus de Tánger, según Español (Fauna
Saudi Arabia, 1. 1979). Edeago de: 12) X. (C.) bucephalus Arabia Saudi, según Toskina (Fauna of Arabia, 17. 1998); 13) X. (C.) materliki del Yemen, según Toskina (Russian Entomological Journal, 11(2). 2002; 14) X. (C.) lepidus de Túnez, según Toskina (Zoosystematica Rossica, 14(2). 2006).
134
Butlletí ICHN 85 (3), 2021
11 10 9 14 13 12 A. VIÑOLAS & J. MUÑOZ-BATET
of
1977 de Canarias (La Palma y Tenerife) por los mismos motivos que X. (C.) bucephalus theanus, sobre todo por la pieza idénticamente conformada del saco interno del edeago.
Toskina (1998) describió la subespecie Xyletinus (Calypterus) bucephalus omanus con una hembra recolectada en Oman. La diferencia del típico X. (C.) bucephalus por el color del cuerpo y de las antenas, por el protórax más transverso y por la puntuación de los intervalos elitrales con tendencia a borrarse en el ápice y zonas laterales. Los datos que aporta para separar la subespecie están incluidos en la variabilidad de X. (C.) bucephalus. En el caso del protórax aporta para X. (C.) bucephalus bucephalus que es 1,5 veces más ancho que largo y para la nueva subespecie da el valor de 1,67, indicando que este dato seguramente se deberá al sexo. En los ejemplares que hemos estudiado la anchura del protórax con relación a su longitud oscila entre 1,59 y 1,78. Los caracteres que aporta para separa X. (C.) bucephalus omanus de X. (C.) bucephalus entran, pues, dentro de la variabilidad especifica de X. (C.) bucephalus
Por lo tanto, se propone que Xyletinus (Calypterus) bucephalus omanus Toskina, 1998 debe pasar a n. syn. de Xyletinus (Calypterus) bucephalus (Illiger, 1807).
El edeago de X. (C.) bucephalus ha sido representado por diferentes autores y con diversos nombres específicos y subespecíficos, aunque en todas las representaciones se puede observar la misma pieza en el saco interno del lóbulo medio; la posición de las pequeñas piezas no tiene ningún valor, ya que forman una corona apical con el saco evaginado (Fig. 11) y con éste retraído su posición suele ser muy variable. En las figuras 7 y 8 representamos el edeago de X. (C.) bucephalus bucephalus de Marruecos y de X. (C.) bucephalus theanus de Jordania respectivamente. En la figura 9 podemos observar la representación del edeago de X. (C.) bucephalus de Marruecos según Español (1979a), en las figuras 10, 11 de Arabia Saudí según Español (1979b), en la figura 12 de Arabia Saudí según Toskina (1998), en la figura 13 de X. (C.) materliki Toskina, 2002 de Yemen y en la figura 14 de X. (C.) lepidus Toskina, 2006 de Túnez.
Toskina (2002) describió Xyletinus (Calypterus) materliki con dos ejemplares procedentes del Yemen, holotipo macho y paratipo sin indicación de sexo. Indica los siguientes caracteres para separar la especie de bucephalus, élitros proporcionalmente más cortos, la peculiar disposición de la pubescencia elitral que parece indicar franjas, la conformación del protórax y su pubescencia de diferente patrón y por la estructura del edeago (disposición de los parámeros y piezas del saco) (Fig. 13).
La proporción elitral longitud/anchura entra dentro de la variabilidad específica de X. (C.) bucephalus, la pubescencia elitral que indica es la propia de ejemplares frescos, no frotados, igualmente sucede con la pubescencia protonal y en la conformación del edeago no encontramos ninguna diferencia, ya hemos indicado que es muy variable la posición de las piezas del saco interno cuando está retraído (Fig. 7 y 13).
Por lo tanto se propone que Xyletinus (Calypterus) materliki Toskina, 2002 debe pasar a n. syn. de Xyletinus (Calypterus) bucephalus (Illiger, 1807)
Toskina (2006) describió Xyletinus (Calypterus) lepidus de Túnez con un ejemplar macho de Sakes (Túnez). Indica que se diferencia de X. (C.) bucephalus, X. (C.) materliki y X. (C.) productus Toskina, 2006 por el estrechamiento elitral y su pubescencia diferentemente dispuesta, por las estrías elitrales borradas en la parte apical y por la diferente estructura de las piezas del edeago.
El carácter de la pubescencia elitral no tiene ningún valor como ya hemos indicado y varia mucho si se trata de ejemplares frescos o muy frotados; entre todos los ejemplares estudiados o revisados hemos observado que algunos poseen las estrías elitrales bien marcadas hasta el ápice, otros las tienen muy poco marcadas en la parte apical y en otros se observa que están totalmente borradas en el ápice, por lo que este carácter no tiene ninguna validez para separar la especie, y tampoco encontramos ninguna diferencia estructural en la conformación del edeago (Figs. 7 y 14).
Por lo tanto se propone que Xyletinus (Calypterus) lepidus Toskina, 2006 debe pasar a n. syn. de Xyletinus (Calypterus) bucephalus (Illiger, 1807).
En la nota sobre Xyletinus (Calypterus) y Xyletinus (Xyletinus) de Toskina (2006) se describen (incluida la especie anterior) veintiuna nuevas especies y dos nuevas subespecies de la región paleártica asiática. Excepto dos, el resto de las especies están descritas con un solo ejemplar, y la gran mayoría son hembras. Tampoco indica el sexo del holotipo en cuatro de las especies descritas. Creemos que se deberá realizar un estudio exhaustivo del contenido específico de dicho trabajo para constatar la validez o no de las especies en él tratadas.
Género Xyletinus Latreille, 1809
Xyletinus Latreille, 1809. Genera Crustaceorum et Insectorum: 376
Xylotinus Sturm, 1826. Catalog meiner Insecten-Sammlung: 59 (variación ortográfica)
Xiletinus Stephens, 1829. A systematic Catalogue of British Insects: 12 (variación ortográfica)
Xytelinus Griffith & Pidgeon, 1832. The class Insecta arranged by Baron Cuvier: 350 (variación ortográfica)
Sternoplus Mulsant & Rey, 1864. Histoire naturelle des Coléoptères de France. Térédiles: 264
Ryletinus Zaitsev, 1956. Akademii Nauk Gruzinskoi SSR, Trudy Instituta Zooloologii, 14: 76 (variación ortográfica)
Xylentinus Santoro, 1957. Revista de Investigaciones Forestales, 1 (4): plancha (variación ortográfica)
Especie tipo: Ptilinus ater Creutzer, 1796, designado por el ICZN.
Subgénero Calypterus Mulsant & Godart, 1859
Calypterus Mulsant & Godart, 1859. Opuscules Entomologiques, 9: 181
Notiomimus Wollaston, 1861. The Annals and Magazine of Natural History, Including Zoology, Botany, and Geology, ser. 3, 7: 15
Especie tipo: Calypterus sericans Mulsant & Godart, 1859, por monotipia (= Ptilinus bucephalus Illiger, 1807)
135 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
CALYPTERUS) BUCEPHALUS S. LAT.
NUEVOS DATOS SISTEMÁTICOS Y DE DISTRIBUCIÓN DE XYLETINUS (
GEA, FLORA ET FAUNA
Definido por los ojos de desarrollo normal sin apenas sobrepasar el contorno de la cabeza; antenas aserradas en ambos sexos; palpos labiales y maxilares con el último artejo fusiforme y aguzado en el ápice (Figs. 4-5); tarsos más cortos que las tibias (Español, 1992).
Xyletinus (Calypterus) bucephalus (Illiger, 1807)
Ptilinus bucephalus Illiger, 1807. Magazin für Insectenkunde, 6: 16
Xyletinus striatipennis Fairmaire, 1857. Annales de la Société Entomologique de France, (3) 5: 638
Calypterus sericans Mulsant & Godart, 1859. Opuscules Entomologiques, 9: 181
Xyletinus peregrinus Chevrolat, 1861. Revue et Magasin de Zoologie Pure et Appliquée, (2) 13: 154
Notiominus holosericius Wollaston, 1861. The Annals and Magazine of Natural History, Including Zoology, Botany, and Geology, ser. 3, 7: 17
Xyletinus (Calypterus) theanus Reitter, 1901. Verhandlungen des naturforschenden Vereines in Brünn, 40: 29 n. syn.
Xyletinus brunnescens Obenberger, 1917. Archiv für Naturgeschichte, A, 82 (4) (1916): 36
Xyletinus megatomoides Roubal, 1917. Societas Entomologica, 32 (3): 11
Xyletinus bucephalus var. bicoloricornis Pic, 1924. L’Échange, Revue Linnéenne, 39 (418): 29
Xyletinus (Calypterus) wollastoni Gottwald, 1977. Acta Entomologica Bohemoslovaca, 74: 61
Xyletinus (Calypterus) bucephalus omanus Toskina, 1998. Fauna of Arabia, 17: 55 n. syn.
Xyletinus (Calypterus) materliki Toskina, 2002. Russian Entomological Journal, 11(2): 207. n. syn.
Xyletinus (Calypterus) lepidus Toskina, 2006. Zoosystematica Rossica, 14(2): 225 n. syn.
Material estudiado de X. (C.) bucephalus 53 ejs., etiquetados: 1 ej. «13-IV-1990, El Hamma, Tunéz, J.L. Moreno & T. Mestre leg»; 1 ej. «IV-1971, Es Castells, Menorca, P. Oromí leg.»; 13 ejs. «10-VIII-1993, La Retuerta, Pina de Ebro, Zaragoza, J. Blasco leg.»; 6 ejs. «26-VII1997, Torrente Vallcorna, Fraga, Huesca, T. Yélamos leg.»; 1 ej. «8-VIII-1997, Serós, Lleida, T. Yélamos leg.»; 13 ejs. «12/19-VII-1998, La Hoya, Almería, J. Mateu leg.»; 1 ej. «15-VII-1999, Serós, Lleida, A. Viñolas leg.»; 3 ejs. «3-VIII2009, Salinas Cabo de Gata, Almería, J. Muñoz leg.»; 1 ej. «8-VII-2010, Garriguella, Girona, J. Soler leg.»; 1 ej. «30VIII-2011, Tabernas, Almería, R. Macià leg.»; 1 ej. «VII2012, Villanueva de Sigena, Huesca, R. Macià leg.»; 2 ejs. «13-VIII-2012, Villanueva de Sigena, Huesca, A. Viñolas leg.»; 1 ej. «5-IV-2014, Rambla de Tabernas, Almería, J. Muñoz leg.»; 1 ej. «28/29-III-2016, Paphos prov., Akamas Mts., 94 m, Cyprus, T. Staněk leg.»; 1 ej. «4-VI-2017, Tahla, Marruecos, M. Tomàs leg.»; 3 ej. «19-V-2018, Rambla de la Sierra, Tabernas, Almería, R. Macià leg.». Depositados en la colección de A. Viñolas.
Material revisado de X. (C.) bucephalus bucephalus 112 ejs., etiquetados: 1 ej. « Fayed, Egipto, H. Priesner leg.»
«MZB | 87-8475»; 16 ejs. «Camarge, Bouches-du-Rhône, Francia, L Puel leg.» «MZB | 87-8450, 87-8461,87-8462, 87-8477, 87-8490, 87-8494, 87-8495, 87-8497»; 2 ejs. «Camarge, Bouches-du-Rhône, Francia, J. Thérond leg.«MZB | 87-8405, 87-8505»; 5 ejs. «Étang de Valcarès, Camargue, Francia» «MZB | 87-8476, 87-8510»; 3 ejs. «Saintes Maries de la Mer, Camargue, Bouches-du-Rhône, Francia, A. Chobaut leg.» «MZB | 87-8470, 87-8488»; 2 ejs. «Collioure, Pyrénées orientals, Francia, V. Mayet leg.» «MZB | 87-8401, 87-8501»; 1 ej. «Marruecos, West, R.M.» «MZB | 87-8486»; 1 ej. «España» «MZB | 76-2929»; 1 ej. « El Alquián, Almería, España, A. Cobos leg.» «MZB | 87-8479»; 1 ej. «Sierra Alhamilla, Almería, España, J. Mateu leg.» «MZB | 87-8485»; 5 ejs. « San Roque, Cádiz, España, J. Ramirez leg.» «MZB | 87-8406, 87-8473, 87-8506»; 1 ej. « San Roque, Cádiz, España, J. Vives leg.» «MZB | 87-8469»; 1 ej. «Archidona, Málaga, España, R. Yus Ramos leg.» «MZB | 87-8407»; 1 ej. «Valls, Tarragona, España, F. Español leg.» «MZB | 87-8465»; 3 ejs. «21/27-V-1914, Étang de Valcarès, Camargue, Francia» «MZB | 87-8512»; 1 ej. «IV-1926, Mohammedia, Grand Casablanca, Marruecos, M. Antoine leg.» «MZB | 87-8496»; 1 ej. «15-V-1929, Taroudant, Marruecos» «MZB | 87-8464»; 2 ejs. «17-XI-1929, Ceuta, España, F. Español leg.» «MZB | 87-8484»; 10 ejs. «13-IV-1932, Camarge, Bouches-du-Rhône, Francia, J. Thérond leg.» «MZB | 87-8471, 87-8472, 87-8493»; 1 ej. «20-VII-1932, Hauta Kasdir, 1750 m, Marruecos, J. de Ferrer leg.» «MZB | 734667»; 1 ej. «IX-1932, Casablanca, Marruecos, M. Antoine leg.» «MZB | 87-8468»; 1 ej. «27-I-1933, Gabal Elba Rabdet, Egipto, H. Priesner leg.» «MZB | 87-8460»; 1 ej. «26-IV1934, Zalboun Tlemcen, Argelia» «MZB | 87-8487»; 4 ejs. «30-VIII-1934, Al Hoceima-Taounate, Taza, Marruecos, P. Rotrou leg.» «MZB | 87-8489»; 1 ej. «20-V-1936, Sirte, Tripolitania, Libia, R. & C. Koch leg.» «MZB | 87-8453»; 1 ej. «21-III-1939, Meadi, Egipto, H. Priesner leg.» «MZB | 878474»; 4 ejs. «-IV-1941, Mariguari, Mazuza, Melilla, España, A. Pardo Alcaide leg.» «MZB | 87-8403, 87-8503»; 1 ej. «07-VI-1946, Tortosa, Tarragona, España, F. Español leg.» «MZB | 87-8466»; 1 ej. «17-VIII-1946, Al Hoceima-Taounate, Taza, Marruecos» «MZB | 87-8498»; 1 ej. «14-IX-1946, Ras el Od, Taza, Marruecos» «MZB | 87-8467»; 1 ej. «11VII-1948, ilegible» «MZB | 87-8402»; 1 ej. «III-1949, Almería, Almería, España, J. Mateu & A. Cobos leg.» «MZB | 878494»; 1 ej.«22-III-1952, Rostrogordo, Melilla, España, A. Pardo Alcaide leg» «MZB | 87-8463»; 5 ejs. «VIII-1955, El Alquián, Almería, España, A. Cobos leg.» «MZB | 87-8478, 87-8451, 87-8452, 87-8500, 87-8509»; 2 ejs. «V-1965, San Roque, Cádiz, España, J. Ramirez leg.» «MZB | 87-8508, 878513»; 1 ej. «VII-1971, San Roque, Cádiz, España, J. Vives leg.» «MZB | 87-8499»; 1 ej. «IX-1973, Rostrogordo, Melilla, España, A. Pardo Alcaide leg.» «MZB | 87-8514»; 1 ej. «26-IX-1973, Rostrogordo, Melilla, España, R. Yus Ramos leg.» «MZB | 87-8481»; 2 ejs. «V-1974, San Roque, Cádiz, España, J. Ramirez leg.» «MZB | 87-8511»; 1 ej. «1976, Qarraah, Khamis, M. 2000 m, Arabia Saudi, W. Büttiker leg.» «MZB | 87-8524» 3 ejs. «VII-1976, Archidona, Málaga, España, R. Yus Ramos leg.» «MZB | 87-8483, 87-8507»; 1 ej.
136 Butlletí ICHN 85 (3),
2021
& J. MUÑOZ-BATET
A. VIÑOLAS
NUEVOS DATOS SISTEMÁTICOS Y DE DISTRIBUCIÓN DE
«09/11-III-1978, Sinkat, 84 m, Kassala, Sudan, H. J. Bremer leg.» «MZB | 87-8528»; 3 ejs. «26-III-1978, El Morche, Málaga, España, R. Yus Ramos leg.» «MZB | 87-8404, 87-482, 87-8504» 1 ej. «28-IV-1984, Illa de Cabrera, Illes Balears, España, E. Petitpierre leg.» «MZB | 76-6196»; 1 ej. «23VII-1988, Las Barrios, Montera del Torero, Cádiz, España, J. de Ferrer leg.» «MZB| 76-6820»; 3 ejs. «10-VIII-1993, La Retuerta, Pina de Ebro, Zaragoza, España, J. Blasco leg.» «MZB | 87-8455»; 9 ejs. «20-VIII-1993, La Retuerta, Pina de Ebro, Zaragoza, España, J. Blasco leg.» «MZB | 87-8454». Depositados en la colección del Museu de Ciències Naturals de Barcelona.
Material estudiado de X. (C.) bucephalus theanus
21 ejs., etiquetados: 6 ejs. «4-X-1965, Mar Muerto, Jordania»; 15 ejs. «5-XI-1966, Mar Muerto, Jordania». Depositados en las colecciones de A, Viñolas y J. Muñoz.
Material revisado de X. (C.) bucephalus theanus
22 ejs., etiquetados: 1 ej. «1-VII-1948, Ofra, Palestinian territories» «MZB | 87-8502»; 9 ejs. «29-IX-1965, Totes Meer, Jordania» «MZB | 87-8480, 87-8517, 87-8518, 87-519, 878520, 87-8522, 87-8527»; 7 ejs. «5-XI-1966, Totes Meer, Jordania» «MZB | 87-8515, 87-8527»; 2 ejs. «7-X-1966, Totes Meer, Jordania» «MZB | 87-8521»; 1 ej. «26-I-1976, Jeddah, Saudi Arabia, W. Büttiker leg.» «MZB | 87-8525»; 1 ej. «15-VI-1976. Jeddah, Saudi Arabia, W. Büttiker leg.» «MZB | 8583 »; 1 ej. «11-V-1976, Jeddah, Saudi Arabia, W. Büttiker leg.» «MZB | 87-8526». Depositados en la colección del Museu de Ciències Naturals de Barcelona.
Longitud de 2,9-4,2 mm. Cuerpo de contorno oval, mate, de color moreno amarillento o rojizo, con la parte superior cubierta de pubescencia corta y muy densa, en especímenes frescos, y que en los élitros forma franjas longitudinales. Cabeza con los ojos bien desarrollados y poco salientes del contorno de la cabeza; antenas de once artejos, aserrados del cuarto al décimo, sobrepasando en longitud el protórax en el macho, más cortas en la hembra; último artejo de los palpos labiales y maxilares fusiforme y aguzado en el ápice (Figs. 4-5); superficie con punteado muy pequeño pero muy denso. Protórax convexo, con una proporción media de 1,8 veces más ancho que largo, con la máxima anchura en la base; márgenes laterales en línea entrante hacia el ápice, este y la base redondeados; ángulos anteriores y posteriores redondeados, estos ligeramente explanados; el contorno pronotal muy finamente bordeado; superficie cubierta de punteado fino y muy denso, similar al de la cabeza y con tendencia a presentarse contiguo junto a los márgenes, ápice y base. Escutelo triangular. Élitros de contorno paralelo en los dos tercios basales y redondeado en el apical, convexos y con una proporción media de 1,57 veces más largos que anchos tomados conjuntamente; húmeros redondeados nada salientes hacia delante; estrías muy marcadas, provistas de gruesos puntos que desbordan ligeramente su contorno; la primera, novena y décima alcanzan el ápice libremente, la segunda unida en el ápice a la cuarta, quinta y octava, la tercera libre y la sexta y séptima unidas entre sí; estría escutelar presente, corta, pero como el resto bien marcada; intervalos ligeramente convexos, provistos de puntos pequeños no muy densamente dispuestos y con pequeños pliegues junto a la base. Abdomen (Fig. 3) con to-
137 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
Figura 15. Mapa con la distribución conocida de Xyletinus (Calypterus) bucephalus
(CALYPTERUS) BUCEPHALUS S. LAT.
XYLETINUS
GEA, FLORA ET FAUNA
dos los segmentos en curva regular, bien indicados, los dos primeros algo más largos que el tercero y cuarto, la superficie cubierta de corta pubescencia, menos densa que en la parte superior del cuerpo. Edeago (Figs. 7-8) con el contorno de los parámeros y de sus lóbulos típico del género; saco interno con una estructura muy curiosa (Fig. 11), provisto de una gran pieza falciforme muy característica. Segmento genital según figura 6.
Distribución
Especie con una amplia distribución paleártica, también se ha reportado de la región afrotropical, citada de Arabia Saudí, Argelia, Bulgaria, Cáucaso, Chipre, Córcega, Cerdeña, Creta, Egipto, España, Francia mediterránea, Grecia, Iraq, Israel, islas Canarias, islas Baleares, Italia en su mitad sur, Jordania, Líbano, Libia, Marruecos, Omán, Portugal, Sicilia, Túnez, territorios Palestinos, Turquía y Yemen (Fig. 15) (Chavanon, 2018; Español, 1979a, b, 1992, 1995; Mulsant & Rey, 1864; Viñolas, 2020; Zahradník, 1998, 2007).
Biología
Tal como ya se ha indicado, la especie realiza su ciclo a expensas de los excrementos secos de herbívoros, con preferencia los del ganado ovino y caprino y en menor medida de equino.
Agradecimientos
A Glória Masó y Berta Caballero, conservadoras del Museu de Ciències Naturals de Barcelona, el habernos permitido consultar y estudiar los ejemplares de la especie depositados en la colección de la entidad.
A los diferentes entomólogos que nos han cedido ejemplares de la especie para estudio, recolectados en sus campañas entomológicas.
Bibliografía
Bercedo, P. & Arnáiz, L. 2008. Nuevos sinónimos de Xyletinus (Calypterus) bucephalus bucephalus (Illiger, 1807) y Calymmaderus (Calymmaderus) solidus (Kiesenwetter, 1877) (Coleoptera: Bostrichoidea: Ptinidae: Xyletininae, Dorcatominae). Boletín de la Sociedad Entomológica Aragonesa, 43: 431-433.
Chavanon, G. 2018. Catalogue des coléoptères de la région orientale du Maroc. (Province de Guercif exceptée). Travaux de l’Institut Scientifique, Série Zoologie, 57: 1-192.
Español, F. 1979a. Los Xyletinus Latr. de Marruecos (Col. Anobiidae. Nota 89). Mediterránea, 3: 3-22.
Español, F. 1979b. Insects of Saudi Arabia. Coleoptera: Fam. Anobiidae. Fauna of Saudi Arabia, 1: 251-256.
Español, F. 1992. Coleoptera, Anobiidae. In: Fauna Iberica, vol. 2. Ramos, M. A. et al. (Eds.). Museo Nacional de Ciencias Naturales. CSIC. Madrid. 195 p.
Español, F. 1995. Anobiidae (Coleoptera) de la fauna íberomarroquí, con una nueva especie de Stagetus Wollaston, 1861 Misceŀlània Zoològica, 17 (1993-1994): 149-152.
Mulsant, E. & Rey, Cl. 1864. Histoire naturelle des coléoptères de France. Tédériles. F. Savy. Paris. 391 p. + 10 pl.
Toskina, I. N. 1998. Rare and new Anobiidae (Coleoptera) from the Arabian Peninsula. Fauna of Arabia, 17: 51-91.
Toskina, I. N. 2002. A new species of the genus Xyletinus (Coleoptera: Anobiidae) from Arabia and a note about Mesocoelopus Russian Entomological Journal, 11(2): 207-208.
Toskina, I. N. 2006. New Palaearctic species of the genus Xyletinus Latreille, 1809 (Coleoptera: Anobiidae). Zoosystematica Rossica, 14 (2): 223-260.
Viñolas, A. 2020. Catálogo comentado de los Ptinidae (Coleoptera) de la Península Ibérica, Islas Baleares e Islas Canarias. Monografies de la Institució catalana d’História Natural, 1. 3-179.
Zahradník, P. 1998. Anobiidae of Turkey. Klapalekiana, 34: 263286.
Zahradník, P. 2007. Ptinidae (excepto subfamilias Gibbiinae y Ptininae). P. 339-362. In: Löbl, I. & Smetana, A. (eds.). Catalogue of Palaearctic Coleoptera. Vol. 4. Apollo Books. Stenstrup. 935 p
138 Butlletí ICHN 85 (3), 2021
& J. MUÑOZ-BATET
A. VIÑOLAS
Neuroterus titou n. sp. from Costa Rica (Cynipidae: Cynipini)
Juli Pujade-Villar* & Paul Hanson**
* Universitat de Barcelona. Facultat de Biologia. Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals. Avda. Diagonal, 645. 08028 Barcelona. Catalonia. A/e: jpujade@ub.edu
** Universidad de Costa Rica. Escuela de Biología. Universidad de Costa Rica. San Pedro, Costa Rica. A/e: phanson91@gmail.com
Corresponding author: Juli Pujade-Villar, A/e: jpujade@ub.edu
Rebut: 14.09.2021; Acceptat: 20.09.2021; Publicat: 30.09.2021
Abstract
A new species of Neuroterus Hartig, 1840 (Hymenoptera: Cynipidae: Cynipini) is described from Costa Rica: Neuroterus titou n. sp. This new species is the second species of the genus Neuroterus recorded from Costa Rica. Neuroterus titou n. sp. induces leaf galls on Quercus copeyensis CH Mull. (Fagaceae, subgen. Quercus, sect. Quercus, White Oaks, Series Leucomexicanae). The diagnostic morphological features, the description of the adult and the gall, and other biological data of this new species are given and discussed.
Key words: Cynipidae, Cynipini, Neuroterus, Quercus, Costa Rica; new species.
Resum
Neuroterus titou n. sp. de Costa Rica (Cynipidae: Cynipini)
Es descriu de Costa Rica una nova espècie de Neuroterus Hartig, 1840 (Hymenoptera: Cynipidae: Cynipini): Neuroterus titou n. sp. Aquesta nova espècie és la segona espècie del gènere Neuroterus registrada a Costa Rica. Neuroterus titou n. sp. indueix gales foliars a Quercus copeyensis CH Mull. (Fagaceae, subgènere. Quercus, sect. Quercus, Alzines blanques, Sèrie Leucomexicanae). Es donen i es discuteixen els caràcters morfològics diagnòstics, la descripció de l’adult i de la gala, i altres dades biològiques d’aquesta nova espècie.
Paraules clau: Cynipidae, Cynipini, Neuroterus, Quercus, Costa Rica, nova espècie.
urn:lsid:zoobank.org:pub:3DB26C70-29BB-46EB-AEF9-4C0B12C1A846
Introduction
Neuroterus Hartig (Hymenoptera, Cynipidae) is one of the more species-rich genera of Cynipini, along with Andricus Hartig (Stone et al., 2002). Since Hartig (1840) first described the genus, based only on European species, Neuroterus has become one of the more problematic genera of Cynipini with respect to its generic limits, and the identification and classifications of the included species (Medianero & Nieves-Aldrey, 2017). It is widely distributed around the world where Quercus is present, although it is not yet known from eastern Panama or Colombia (in America) and the Oriental region (in Eurasia). The character that allows recognition of the genus (in its traditional sense) is the absence of a transcutellar suture separating the mesoscutum and the scutellum. For many years all Cynipini without this suture were included in Neuroterus, thereby becoming a chaotic and non monophyletic genus (Liljeblad et al.; 2008; Melika et al., 2010). In the Eurasian area Pujade-Villar et al. (2004) considered the subgenus Pseudoneuroterus Kinsey as a valid genus, and Melika et al. (2010) described Cerroneuroterus Melika & Pujade-Villar for some Neuroterus inducing galls on oaks in the Cerris section and considered the subgenus Latuspina Monzen as a valid genus. Mayr (1907) described from the eastern Palearctic the genus Trichagalma, which does not have a transscutel-
lar suture but is very pubescent (unlike Neuroterus which is glabrous). Thus, in the Palearctic region there are five genera without a transcutellar suture that induce galls on Quercus More recently, a new genus closely related to Neuroterus, Cycloneuroterus Melika & Tang, was described for a group of Oriental species (Taiwan and mainland China) associated with Cyclobalanopsis, Lithocarpus and Castanopsis (Tang et al., 2011; Pénzes et al., 2018).
In America all species without a transcutellar suture are included in Neuroterus, but these species need to be revised and should probably be divided into several genera as was done for the Eurasian species. In fact, Pujade-Villar et al. (2015) argued that Nearctic Neuroterus is a polyphyletic group as currently defined and suggested that new genera will need to be established. An attempt in this direction was made by Kinsey (1923), who recognized 6 subgenera, 3 of them for the American species.
Neuroterus includes around eighty species (Melika et al., 2010; Medianero & Nieves-Aldrey, 2017; Pénzes et al., 2018; Pujade-Villar et al., 2018), sixty-five of them from America: 51 have been listed from the United States and Canada (Burks, 1979; Melika & Abrahamson, 1997); 11 from Mexico (Pujade-Villar et al., 2018), 2 from Panama (Medianero & Nieves-Aldrey, 2017) and a single species from Costa Rica (Medianero & Nieves-Aldrey, 2017).
139 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA GEA,
FLORA ET FAUNA
Butlletí de la Institució Catalana d’Història Natural, 85 (3): 139-145. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition)
10.2436.20.1502.01.86
DOI:
The aim of this study is to describe a new species of Neuroterus from Costa Rica belonging to the subgenus Diplobius Kinsey and producing a leaf gall very similar to that of N. pulchrigalla, which was described and illustrated by Medianero & Nieves Aldrey (2017).
Material and methods
Adult gall wasps were reared from galls collected in Cerro de la Muerte by the second author on Quercus copeyensis belonging to section Quercus, in 1997 and 2000. The galls were stored in rearing containers. The adults emerged under laboratory conditions and were preserved in ethanol, with selected specimens subsequently dried and point mounted.
We follow Liljeblad & Ronquist (1998) and Melika (2006) with respect to morphological terminology, except abbreviations for forewing venation follow Ronquist & Nordlander (1989); cuticular surface terminology follows that of Harris (1979). Measurements and abbreviations used here include: F1–F12, 1st and subsequent flagellomeres; POL (post-ocellar distance) is the distance between the inner margins of the posterior ocelli; OOL (ocellar-ocular distance) is the distance from the outer edge of a posterior ocellus to the inner margin of the compound eye; LOL, the distance between lateral and frontal ocelli. The width of the forewing radial cell is measured from the margin of the wing to the Rs vein.
The type specimens of the newly described species are deposited in the following institutions: UB, Universitat de Barcelona, Catalonia (Juli Pujade-Villar col.); MZUCR, Museo de Zoología, Universidad de Costa Rica (Paul Hanson curator).
The SEM pictures were acquired using a field-emission gun environmental scanning electron microscope (FEI Quanta 200 ESEM), for high-resolution imaging without gold-coating the specimens. Gall images were taken with a Canon A1 camera with a 50 mm macro lens followed by Adobe Photoshop CS3 software processing. Adult images were taken with an Olympus SC30 camera, coupled to an Olympus U–CMAD3, adapted to an Olympus SZX10 stereomicroscope. Multiple photograph combinations were achieved with a Helicon Focus 6.2.2 for stacking and image processing. Forewing with a Canon camera (Power Shot SX 210 IS) directly from the microscope eyepiece.
Results
Neuroterus titou Pujade-Villar & Hanson n. sp. (Figs. 1-4)
urn:lsid:zoobank.org:act:9D8091E3-B083-4B46-95F4-90CB6B5DF612
Diagnosis
The galls are similar to those of Neuroterus pulchrigalla Medianero & Nieves-Aldrey, 2017 but the adults have 12

140 Butlletí ICHN 85 (3), 2021
FLORA ET FAUNA J. PUJADE-VILLAR & P. HANSON
GEA,
a c b
Figure 1. Neuroterus titou n. sp: a) head in frontal view; b) head in dorsal view; c) antenna.

flagellomeres and the mesoscutum is coriaceous and basally with some longitudinal rugae (11 flagellomeres and almost smooth mesoscutum in N. pulchrigalla). These morphological characters mentioned for the new species are also present in N. glandiphilus Nieves-Aldrey & Medianero, 2017 but the galls of this latter species occur in acorns. Morphologically, N. titou n. sp. differs from N. glandiphilus in the clypeus (rounded in N. titou n. sp., incised medially in N. glandiphilus), in the POL-OOL length (POL 2.0x as long as OOL and OOL 1.3x as long as diameter of lateral ocellus in N. titou n. sp., POL 1.6x as long as OOL and OOL 2.6x as long as diameter of lateral ocellus in N. glandiphilus) and in the forewings (radial cell 3.8x as long as broad in N. titou n. sp.; radial cell 4.3x as long as broad in N. glandiphilus). Also, the new species is an asexual form and N. glandiphilus a sexual form (see discussion).
Type material
HOLOTYPE deposited in UB (col. JP-V) with the following labels: ‘Costa Rica, San José, Cerro de la Muerte. Est. Biol. Cuericí. 2600 m., ii.2000. P. Hanson’ (white label); ‘Q. copeyensis, bristly cylinder’ (white label); ‘Holotype of Neuroterus titou Pujade-Villar & Hanson n. sp. ǒ, desig. JP-V 2021’ (red label). PARATYPES (6ǒ deposited in UB, 3 in MZUCR): 6ǒ with same data of holotype; 2ǒ idem ii.1997; 1ǒ idem ix.1997.
Description (Figs. 1-3, 4c-d)
Asexual Female
Body length, 1.7 – 2.0 mm (N=7)
Color
Body chestnut to dark brown. Scape, pedicel and F1 yellowish, remaining flagellomeres darker. Mandibles yellowish
141 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA NEUROTERUS TITOU N. SP. FROM COSTA RICA (CYNIPIDAE: CYNIPINI)
Figure 2. Neuroterus titou n. sp.: a-b) mesosoma in dorsal view; c) propodeum; d) mesosoma in lateral view.
d c b a
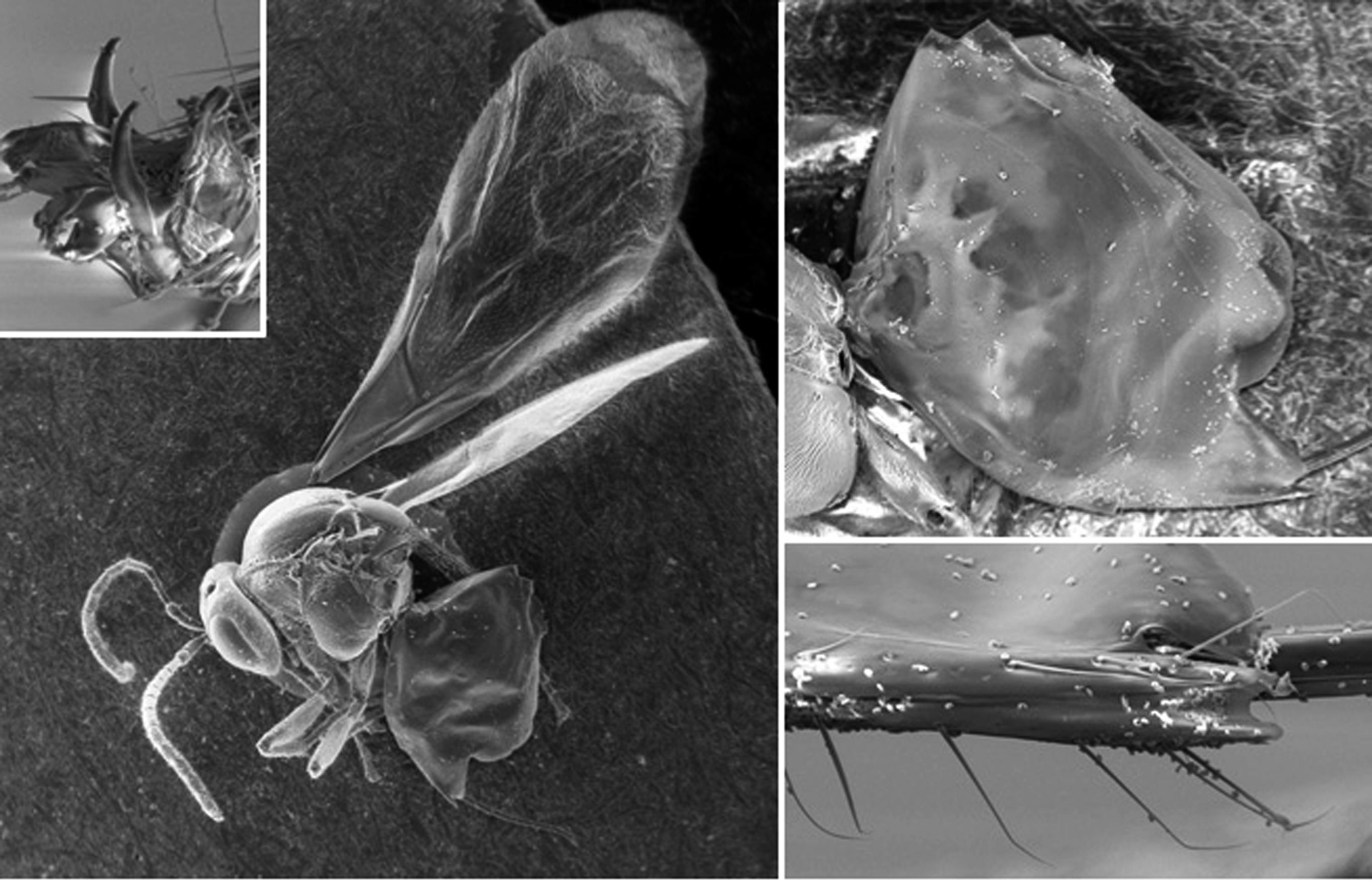
with red tooth with black margin. Legs predominantly brownyellowish, with coxae distally, trochanters, distal femora and metatibiae darker. Forewings hyaline with veins brown.
Head
Transversely oval in frontal view, 1.2x as high as broad; in dorsal view 2.5x as broad as long and as broad as mesosoma. Vertex, frons and face uniformly coriaceous; vertex and frons without setae, face with sparse white setae in the centre. Genae very slightly expanded behind eyes. Clypeus trapezoidal, as wide as high, shining, moderately pubescent, ventral margin rounded and projecting over mandibles; median area of clypeus with alutaceous sculpture; lateral and ventral projecting areas smooth. Anterior tentorial pits conspicuous; epistomal sulcus present and clypeo-pleurostomal lines inconspicous. Malar space 0.3x the height of compound eye, with a distinctive, complete and well-impressed malar sulcus running through a smooth or nearly smooth area. Transfacial line 1.2x as long as height of eye. Compound eyes slightly converging ventrally. Toruli located at mid-height of head, diameter of antennal torulus 1.3x as long as distance between them, distance between torulus and eye as long as diameter of torulus. Ocellar plate slightly raised. POL 2.0x as long as OOL, OOL 1.3x as long as diameter of lateral ocellus and longer than LOL; all ocelli ovate and of the same size.
Antenna
Longer than head+mesosoma, with 12 flagellomeres; pedicel 1.5x as long as broad; F1 2.0x as long as pedicel and
1.3x as long as F2; F2 1.1x as long as F3; F3=F4, F5=F6 but slightly shorter than F4, F7 to F12 slightly shorter than F6 and subequal. Relative lengths of antennal segments: 5/3/6/ 4.5/4/4/3.5/3.5/3/3/3/3/3/3. Placodeal sensilla white, distinct on F3–F12.
Mesosoma
Short, around 1.2x as long as broad in dorsal view, very slightly longer than high in lateral view and with dorsal margin strongly convex. Pronotum medially very short, pronotal plate indistinct, with coriaceous sculpture in lateral view with some carinae on posterior margin; glabrous, pubescent only on the anterior margin. Mesoscutum coriaceous, glabrous, basally with some longitudinal rugae visible. Notauli indistinct but shortly traceable basally; median mesoscutal impression absent; anteroadmedian signa reaching 3/4 mesoscutal length, parapsidal signa absent; parascutal carina broad, reaching anterior part of mesoscutum. Transcutellar suture absent. Scutellum nearly circular or very slightly longer than broad, approximately 0.6x as long as mesoscutum, smooth on dorsal surface and disk sparsely pubescent, circumscutellar carina present with some short rugae; coriaceous and rugose distally behind the circumscutellar carina. Scutellar foveae in form of deep, transverse, shining, inverted V-shaped groove. Scutellum not overlapping metascutelum posteriorly in lateral view. Mesopleuron (including speculum) weakly coriaceous, alutaceous in the lower part and glabrous on entire surface; mesopleural triangle alutaceous with some carinae, sparsely pubescent. Metapleural
142 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
J. PUJADE-VILLAR & P. HANSON d c b a
Figure 3. Neuroterus titou n, sp.: a) tarsal claws; b) habitus; c) metasoma in lateral view; d) ventral spine of hypopygium in ventral view.

sulcus poorly defined, reaching posterior margin of mesopectus at about half the distance from ventral margin, lower part glabrous, delimiting smooth area with some weak rugae, upper part of sulcus absent. Metascutellar carinae rugose with strongly coriaceous sculpture; metanotal trough smooth and glabrous. Propodeum almost smooth and very slightly pubescent on the sides; lateral propodeal carinae absent; medial area with an inconspicuous medial coriaceous carina and with a coriaceous rugose area next to the nucha; basally and laterally with some weak radiating carinae; nucha very short inconspicuously sculptured.
Legs
Tarsal claws simple, without a basal lobe or tooth.
Forewing
1.7x as long as body, hyaline, setose; apical margin with a fringe of long setae. Radial cell 3.8x as long as broad, open along anterior margin, areolet triangular, closed and distinct; R1 and Rs reaching margin of wing; Rs not expanded at apex but continuing slightly parallel to the margin; Rs+M complete or almost reaching the basal vein at its midpoint. First abscissa of radius (2r) curved. Basal cell glabrous and costal cell setose.
Metasoma
Shorter than head + mesosoma combined; as long as high in lateral view. All metasomal tergites smooth, glabrous, second metasomal tergum extending to 2/3 length of metasoma in dorsal view, with very few and sparse short white setae anterolaterally; all terga and hypopygium without micropunctures, prominent part of ventral spine of hypopygium slightly longer than broad in ventral view; lateral margins of
hypopygial spine with setae, the subapical ones slightly projected over apical end of the spine.
Galls (Fig. 4a-b).
The galls produced by this species have the form of upright cylinders, measuring 4.5 × 2.0 mm., unilocular, located on the undersides of the leaves, inserted on a primary or secondary vein. The gall surface is covered with long, erect and dense yellowish to brown hairs. The color is yellow when fresh, orange when mature and brown when old. The galls are generally found in clusters and are oriented side by side; they are rarely isolated. Internally, two parts are visible; an ovoid larval cell at the attachment of the gall on the leaf and an apical empty part.
Biology
N. titou n. sp. induces galls on Q. copeyensis CH Mull. (section Quercus), and only the asexual form is known. Trees were not examined throughout the year and so the phenology is incompletely known. However, the vast majority of adults emerged in February.
Distribution
Currently only known from Costa Rica (Cerro de la Muerte).
Etymology
Named in honour of our friend and French biologist recently deceased: Roger Folliot (25th November 1925 - 23th of July 2021). ‘Titou’ was the nickname he had during the resistance in WWII (http://asaapicardie3945.fr/index.php/ english/rescuers/219-roger-folliot).
143 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
NEUROTERUS TITOU N. SP. FROM COSTA RICA (CYNIPIDAE: CYNIPINI) d b c a
Figure 4. Galls of Neuroterus titou n. sp.: a) several galls on the underside of Q. copeyensis leaf; b) detail of an old gall; c) habitus; b) forewing.
Discussion
The new species described here, and the species described from Panama (Medianero & Nieves-Aldrey, 2017), show some important morphological diagnostic characters that do not fit with those defining Neuroterus given by Melika et al (2010). According to Medianero & Nieves-Aldrey (2017) the anomalous characters are mainly the simple tarsal claws, antenna with 11/12 flagellomeres and the propodeal sculpture.
The morphology of the asexual form described here resembles the sexual form of N. glandiphilus (see diagnosis), which occurs in acorn galls. Thus, it is possible that the new species could be the asexual form of N. glandiphilus
Nevertheless, Neuroterus species are extremely uniform in morphology, and finding reliable diagnostic characters that allow for their identification is extremely difficult (Medianero & Nieves-Aldrey, 2017). Also, Medianero & Nieves-Aldrey (2017) mentioned that although the asexual generation is unknown in N. glandiphilus, a damaged female, presumably asexual according its morphology, was collected from different, smaller galls in the acorn cup, which may be the alternating form of N. glandiphilus. Both N. titou n. sp. and N. glandiphilus belong to the subgenus Diplobius of Kinsey (1923), but differ from the American species included in this subgenus in the number of antennal segments (13 in the subgenus Diplobius, 13-14 in N. glandiphilus and 14 in N. titou n. sp.). Kinsey (1923) mentions that the sexual and asexual forms of species belonging to the subgenus Diplobius have nearly identical galls, but the galls of N. titou n. sp. occur on leaves and the galls of N. glandiphilus are in acorns.
The gall of the new species described here is extremely similar (almost identical) to that of N. pulchrigalla described and illustrated by Medianero & Nieves-Aldrey (2017), which is also an asexual form. However, females of N. titou n. sp. have a 14 segmented antenna whereas N. pulchrigalla have a 13-segmented antenna. The mesoscutum of the former has a coriaceous mesoscutum whereas the latter species has a nearly smooth mesoscutum.
Finally, it should be noted that Neuroterus titou n. sp. was collected from galls on Q. copeyensis CH Mull. but, according to Medianero & Nieves-Aldrey (2017), N. glandiphilus and N. pulchrigalla were collected on Q. bumelioides Liebm. These authors, based on Kappelle (1996), considered Q. copeyensis to be a synonym of Q. bumelioides. The name Q. bumelioides was also used instead of Q. copeyensis in the manual of plants of Costa Rica (Morales, 2010). However, Q. bumelioides is currently considered to be a synonym of Q. sapotifolia Liebm. (Hélardot, 2020), which belongs the section Lobatae whereas Q. copeyensis belongs to the section Quercus. Based on the illustrations in Medianero & Nieves-Aldrey (2017), we believe that they also collected from Q. copeyensis
Acknowledgements
We are very grateful to Víctor Cuesta-Porta (Facultat de Biologia. Departament de Biologia Evolutiva, Ecologia i
Ciències Ambientals, UB) for taking the photograph of the habitus shown in figure 4c.
Bibliography
Burks, B. D. 1979. Superfamily Cynipoidea. In: K. V. Krombein, P. D. Hurd Jr., D. R. Smith & B. D. Burks (eds.). Catalog of Hymenoptera in America of North of Mexico. Volume 1. Symphyta and Apocrita Smithsonian Institution Press. Washington: 1045-1107.
Harris, R. 1979. A glossary of surface sculpturing. Occasional Papers in Entomology, 28: 1-31.
Hartig, T. 1840. Ueber die Familie der Gallwespen. III. Zeitschrift für Entomologie (Germar), 2: 176-209.
Hélardot, J. L. 2020. Oaks of the world. Disponible en: http://oaks. of.the.world.free.fr [Last consulted: 21-08-2021].
Kappelle, M. 1996. Los Bosques de Roble (Quercus) de la Cordillera de Talamanca, Costa Rica: biodiversidad, ecología, conservación y desarrollo. Instituto Nacional de Biodiversidad y Universidad de Amsterdam. Heredia. 336 p.
Kinsey, A. C., 1923. The gall wasp genus Neuroterus (Hymenoptera). Indiana University Studies, 58: 1-150.
Liljeblad, J. & Ronquist, F. 1998. A phylogenetic analysis of higher-level gall wasp relationships (Hymenoptera: Cynipidae). Systematic Entomology, 23, 229-252. https://doi.org/10.1046/ j.1365-3113.1998.00053.x
Liljeblad, J., Ronquist, F., Nieves-Aldrey, J. L, Fontal-Cazalla, F., Ros-Farre, P., Gaitros, D. & Pujade-Villar, J. 2008. A fully webillustrated morphological phylogenetic study of relationships among oak gall wasps and their closest relatives (Hymenoptera: Cynipidae). Zootaxa, 1796: 1-73.
Mayr, G. 1907. Zwei Cynipiden, Marcellia, 6: 3-7.
Medianero, E. & Nieves-Aldrey, J. L. 2017. First record of the oak gall wasp genus Neuroterus Hartig, 1840 (Hymenoptera, Cynipidae, Cynipini) from Central America with description of three new species from Panama and Costa Rica. Graellsia, 73 (1): e057. http://dx.doi.org/10.3989/graellsia.2017.v73.178.
Melika, G. 2006. Gall wasps of Ukraine. Cynipidae. Vestnik Zoologii, suppl., 21 (1): 1-300.
Melika, G. & Abrahamson, W. G. 1997. Descriptions of four new species of cynipid gall wasps of the genus Neuroterus (Hymenoptera: Cynipidae) with redescriptions of some known species from the eastern United States. Proceedings of the Entomological Society of Washington, 99: 560-573.
Melika, G., Pujade-Villar, J., Abe, Y., Tang, C. T., Nicholls, J., Wachi, N., Ide, T., Yang, M. M., Pénzes, Z. S., Csóka, G. Y. & Stone, G. N. 2010. Palaearctic oak gallwasps galling oaks (Quercus) in the section Cerris: re-appraisal of generic limits, with descriptions of new genera and species (Hymenoptera: Cynipidae: Cynipini). Zootaxa, 2470: 1-79.
Morales, J. F. 2010. Fagaceae. In: B. E. Hammel, M. H. Grayum, C. Herrera & N. Zamora (eds.). Manual de Plantas de Costa Rica, Vol. V. Dicotiledóneas (Clusiaceae – Gunneraceae). Monographs in Systematic Botany from the Missouri Botanical Garden, 119: 776-781.
Pénzes, Z., Tang, C. T., Stone, G. N., Nicholls, J. A., Schwéger, S., Bozsó, M. & Melika, G. 2018. Current status of the oak gallwasp (Hymenoptera: Cynipidae: Cynipini) fauna of the Eastern Palaearctic and Oriental Regions. Zootaxa, 4433 (2): 245-289. https://dx.doi.org/10.11646/zootaxa.4433.2.2
Pujade-Villar, J., Cibrián-Tovar, D., Barrera-Ruiz, U. M. & Cuesta-Porta, V. 2018. Las especies de Neuroterus (Hartig, 1840) de México, con la descripción de una especie nueva (Hym., Cynipidae). Entomología mexicana, 5: 453-461.
144 Butlletí ICHN 85 (3), 2021
GEA, FLORA ET FAUNA
PUJADE-VILLAR & P. HANSON
J.
Pujade-Villar, J., Cibrián-Tovar, D., Barrera-Ruíz, U. M. & Melika, G. 2015. First record of Neuroterus galls on twigs in Mexico with description of two new species (Hym.: Cynipidae). Butlletí de la Institució Catalana d’Història Natural, 78 (2014): 3-8.
Pujade-Villar, J., Kovalev, O. & Liljeblad, J. 2004. Status of Pseudoneuroterus Kinsey, 1923 and remarks on closely related genera (Hymenoptera, Cynipdiae). Nouvelle Revue d’Entomologie (Nouvelle Série), 20: 353-360.
Ronquist, F. & Nordlander, G. 1989. Skeletal morphology of an archaic cynipoid, Ibalia rufipes (Hymenoptera: Ibaliidae). Entomo-
logica Scandinavica, Supplement, 33: 1-60.
Stone, G. N., Schönrogge, K., Atkinson, R. J., Bellido, D. & PujadeVillar, J. 2002. The population biology of oak gall wasps (Hymenoptera: Cynipidae). Annual Review of Entomology, 47: 633668.
Tang, C. T., Melika, G., Nicholls, J. A., Yang, M. M. & Stone, G. N. 2011. A new genus of oak gallwasps, Cycloneuroterus Melika & Tang, with the description of five new species from Taiwan (Hymenoptera: Cynipidae: Cynipini). Zootaxa, 3008 (1): 33-62. https://dx.doi.org/10.11646/zootaxa.3008.1.2
145 Butlletí ICHN 85 (3), 2021
GEA, FLORA ET FAUNA
NEUROTERUS TITOU N. SP. FROM COSTA RICA (CYNIPIDAE: CYNIPINI)
NOTA BREU
Rebut: 26.07.2021. Acceptat: 20.09.2021. Publicat: 30.09.2021 Butlletí de la Institució Catalana d’Història Natural, 85 (3): 147-150. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print
Sobre la variabilitat de Pedicularis pyrenaica i la presència de Pedicularis mixta (Orobanchaceae) als Pirineus orientals
On the variability of Pedicularis pyrenaica and the presence of Pedicularis mixta (Orobanchaceae) in the Eastern Pyrenees
Ignasi Soriano*
* Departament de Biologia Evolutiva, Ecologia i Ciències ambientals. Secció Botànica i Micologia. Institut de Recerca de la Biodiversitat. Diagonal, 643. E 08028 Barcelona. A/e: isoriano@ub.edu
El gènere Pedicularis es troba representat als Pirineus per una dotzena de tàxons, entre espècies i subespècies, d’herbes hemiparàsites de dimensions mitjanes o petites (Atlas de la flora dels Pirineus, 2021a). Sobre aquestes plantes hem publicat diversos treballs, com la síntesi de Flora iberica (Soriano, 2009) o les fitxes de les dues versions de l’Atlas de la Flora dels Pirineus (2021a, 2021b). En aquesta nota presentem diverses precisions i observacions sobre dues de les espècies del gènere complicades de destriar en alguns casos, un problema que les imatges publicades en línia els darrers anys ha fet més evident.
Pedicularis pyrenaica J. Gay i P. mixta Gren. són dos endemismes pirenaicocantàbrics de la Secció Rostratae, la més diversificada i complexa del gènere dins l’àmbit geogràfic
esmentat. Les flores de Bolòs & Vigo (1996) i Bolòs et al. (2005) les agrupen en una única espècie (P. pyrenaica en sentit ampli), amb rang de subespècies: P. pyrenaica subsp. pyrenaica i P. pyrenaica subsp. lasiocalyx (Gren.) O. Bolòs & Vigo, respectivament. Segons Flora iberica (Soriano, 2009) i Flora gallica (Tison et al., 2014) els trets distintius més evidents entre els dos tàxons es troben en els òrgans reproductors: color de la coroŀla, nombre de flors i forma de la inflorescència, i indument del calze i les bràctees (vegeu Taula 1 i Figs. 1 i 2). Per nosaltres, un dels caràcters de més valor diagnòstic és sens dubte el color de la coroŀla: concolora, rosa viu, a P. pyrenaica (Figs. 1a-b); discolora, amb el llavi superior porpra i l’inferior rosa pàŀlid, a P. mixta (Figs. 2a-b) Aquest tret, però, només és apreciable en exemplars vius o


147 Butlletí ICHN 85 (3), 2021
BREU
NOTA
edition)
DOI: 10.2436.20.1502.01.87
Figura 1– Pedicularis pyrenaica. a) Hàbit de la planta (Vallferrera);b) detall de les inflorescències (vall d’Eina).
b a


bTaula 1. Comparació de les principals característiques morfològiques de Pedicularis pyrenaica i P. mixta segons Flora iberica (Soriano, 2009) i Flora gallica (Tison et al., 2014), lleugerament modificades pel que fa a P. pyrenaica
Pedicularis pyrenaica
Pedicularis mixta
Inflorescència amb menys de 15(20) flors, que s’obren gairebé Inflorescència amb flors nombroses (15-35), que s’obren de alhora, en raïm capitat o curtament espiciforme. manera gradual, en un llarg raïm espiciforme.
Coroŀla concolora o gairebé, rosa viu, de vegades amb el llavi Coroŀla discolora, amb el llavi superior porpra i el llavi inferior superior de color un xic més viu que l’inferior. rosa pàŀlid.
Calze i bràctees glabres o + densament pilosos.
dessecats amb molta cura, perquè es deteriora ràpidament en premsar-los per fer-ne plecs d’herbari.
Pel que fa a ecologia i distribució P. mixta és una planta exclusiva, o gairebé, de les molleres cantàbriques i dels Pirineus centrals, que vers l’est arribaria fins Andorra pel vessant sud (Carrillo et al., 2008), i fins el Conflent pel vessant nord (Bolòs & Vigo, 1996). Al seu torn, P. pyrenaica es distribueix arreu dels estatges superiors de les muntanyes pirenaicocantàbriques; viu en pastures d’altitud, sobretot acidòfiles, però també en molleres i sòls humits (Soriano, 2013, 2018). En aquests darrers ambients sovint es troba representada per morfotips de característiques més o menys diferenciats dels de les pastures, amb diversos trets que els apropen a P. mixta, com les tiges erectes o curtament ascendents i les inflorescències un xic allargades, amb una desena llarga de flors; en canvi, les P. pyrenaica dels prats solen presentar tiges corbades-ascendents i inflorescències curtes i denses, sovint amb menys flors. A més, en algunes àrees, sobretot de la meitat
Calze i bràctees amb pilositat llanosa densa.
oriental dels Pirineus, els calzes i les bràctees de les poblacions de les molleres es troben coberts d’una pilositat poc o molt densa (Fig. 3), característica que, val a dir, també s’observa esporàdicament en alguns exemplars de pastures. Que nosaltres sapiguem, fins ara no s’ha donat entitat taxonòmica a aquestes plantes, més enllà de l’etiquetatge d’algunes mostres d’herbari com a presumptes híbrids de P. pyrenaica i P. mixta.
Les claus de les flores de Bolòs & Vigo (1996) i de Bolòs et al. (2005) distingeixen els dos taxons per l’indument del calze i les bràctees i la forma de la inflorescència (vegeu la Taula 2), però no contemplen el color de la coroŀla. Per tant, si en determinar exemplars de calze i bràctees pilosos, inflorescència més o menys desenvolupada i coroŀla concolora (o bé mancats de coroŀla, o amb les coroŀles passades) hom fa atenció sobretot a l’indument, s’arriba a la subespècie lasiocalyx (= P. mixta), i aleshores el problema està servit. De fet, diverses fotografies publicades en línia atribueixen a P. mixta
148 Butlletí ICHN 85 (3), 2021 NOTA BREU I. SORIANO
Figura 2. Pedicularis mixta. a) Hàbit de la planta (Vall d’Aran);b) detall de la inflorescència (massís de Mampodre, León).
a
Taula 2. Comparació de les característiques morfològiques de Pedicularis pyrenaica subsp. pyrenaica i P. pyrenaica subsp. lasiocalyx segons la Flora dels Països catalans (Bolòs & Vigo, 1996).
Pedicularis pyrenaica subsp. pyrenaica
Calzes i bràctees glabres o subglabres.
Pedicularis pyrenaica subsp. lasiocalyx
Calzes i bràctees pilosos, llanosos. Espiga curta i compacta. Espiga més oblonga i laxa.
plantes de la part oriental dels Pirineus d’aquestes característiques amb la coroŀla concolora: vegeu, per exemple, Pedicularis pyrenaica subsp. lasiocalyx | Flora Catalana; https:// www.tela-botanica.org/bdtfx-nn-48159-illustrations; https:// www.biodiversidadvirtual.org/herbarium/Pedicularis-mixta1de5-img419742.html. Nosaltres mateixos hem observat i recoŀlectat plantes similars a les muntanyes que envolten la Cerdanya, i més a l’oest; també als herbaris BCN, BC i P, se’n conserven diverses mostres, de les que en donem la relació a l’annex 1.
Davant del dubte de si els noms Pedicularis mixta i P. pyrenaica subsp. lasiocalyx fan referència a la mateixa planta, cal remarcar que P. pyrenaica subsp. lasiocalyx, i el basiònim P. pyrenaica var. lasiocalyx Gren., són sinònims taxonòmics de P. mixta. De fet, en la descripció de la var. lasiocalyx a la flora de Grenier & Godron (1853: 617), s’hi explicita P. mixta com a sinònim; d’altra banda, el nom de l’espècie fou publicat aquell mateix any dins els Archives de la flore de France et Allemagne editats per F.W. Schulz (Grenier, 1853), amb referències a les mateixes localitats que la var. lasiocalyx.
Hem pogut examinar imatges dels materials utilitzats per Grenier per descriure la planta (sintipus) tant en el rang d’espècie com en el de varietat, els quals procedeixen dels Pirineus centrals (Castanesa, vessant sud del port de Benasc, Cauterets) i orientals (rodalia de Montlluís). Les caracteristiques dels primers es corresponen amb les indicades de P. mixta a la taula 1; per contra, l’exemplar de Montlluís (P00403949 https://science.mnhn.fr/institution/mnhn/collection/p/item/p 00403949?listIndex=2&listCount=114), per la inflorescència compacta, amb flors que semblen haver aparegut simultàniament i de coloració no discernible donada l’antiguitat de la mostra, podria correspondre a una d’aquelles P. pyrenaica de calze densament pilós a què hem fet referència, i que com hem indicat són presents en localitats properes. Fem notar encara que Grenier no fa referència al color de la coroŀla en les descripcions, probablement perquè tot i que afirma haver observat la planta al camp (el 1836 a Castanesa i el port de Benasc: veure Grenier, 1853), la va descriure força anys més tard, a partir de materials propis però també de plecs d’herbari que li van ser enviats (de Montlluís i Cauterets), en els que el color de la coroŀla no es devia poder apreciar. Semblaria doncs que el problema que plantegem no és nou.
Sobre la base de tota aquesta informació, pensem que convé reconsiderar la interpretació que s’ha fet en força casos de P. mixta, i reservar aquest nom per les plantes que presenten el conjunt de característiques indicades a les flores recents, resumides a la taula 1. Pel que fa als morfotips de P. pyrenaica repetidament esmentats, sovint atribuïts a P. mixta, proposem donar-los entitat taxonòmica, ni que sigui modesta, i considerar-los una nova varietat subordinada a P. pyrenaica,
amb un nom que aŀludeix a la pilositat que recobreix el calze i les bràctees (Fig. 3):
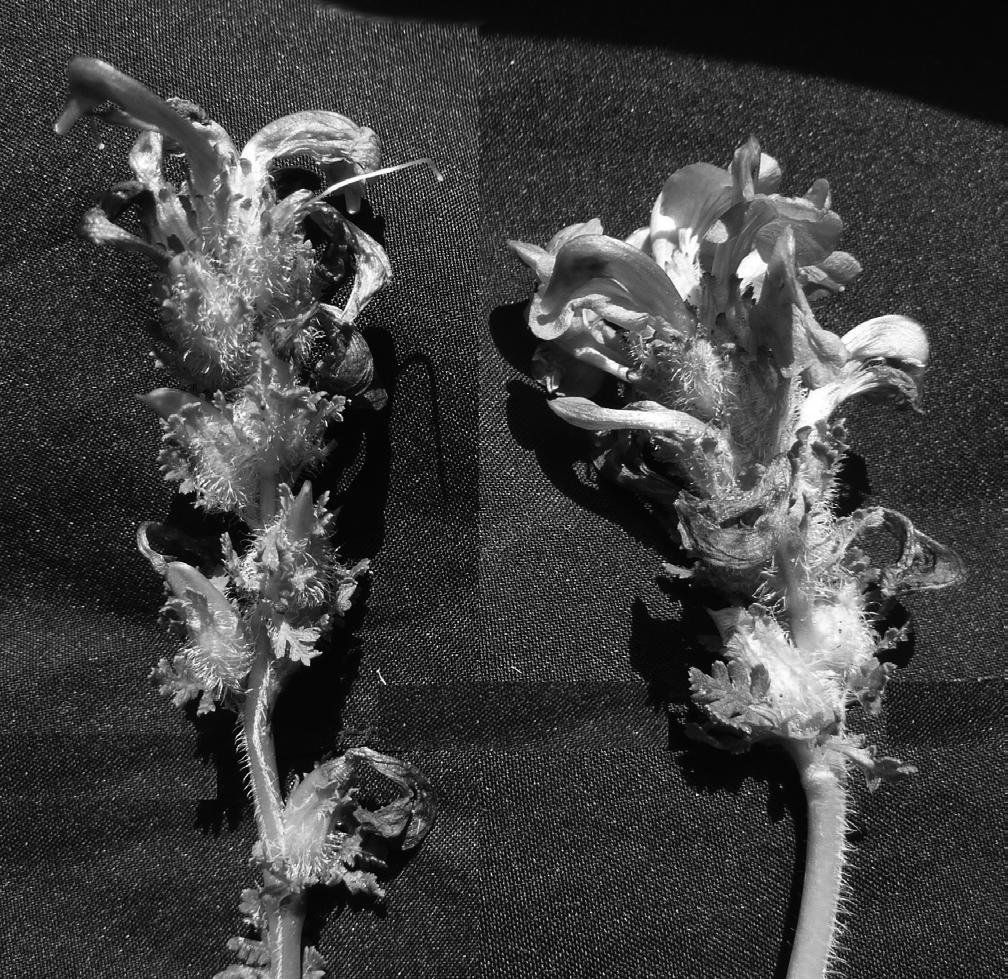
3. Pedicularis pyrenaica var. pilifera nova. Inflorescències del tipus de la varietat, en fresc (Conflent, massís de Madres).
Pedicularis pyrenaica var. pilifera nova
Stems erect or ascendent-erect; inflorescence slightly elongated, usually with more than 10 flowers; calyx and bracts hairy; corolla pink, concolorous. It lives mainly in wetlands and spring margins, in the Eastern part of the Pyrenees.
Holotype
El Conflent, massís de Madres, coma de Pontells, marges de rierols amb molleres i torberes d’esfagnes, 31T DH322209, 1.930 m, 12.VIII.2020, I. Soriano 3514 (BCN 161365).
Pel que fa a la presència (i eventualment l’abundància) de P. mixta a la part oriental dels Pirineus, del territori del Principat situat a l’est d’Andorra no ens en consten testimonis ni d’herbari ni fotogràfics que li siguin atribuibles de manera inequívoca. L’exemplar barrejat amb P. pyrenaica d’un plec de Núria de l’herbari Sennen (BC 981737) al que fa referència Vigo (1983) actualment és molt malmès i impossible de determinar. Quant al sector oriental del vessant nord, tot i que hi ha força citacions de territori administrativament francès (vegeu l’Atlas de la flora dels Pirineus, 2021a) entenem que almenys una part, o qui sap si totes, corresponen a la nova varietat de P. pyrenaica.
Recomanem doncs verificar, tant al camp com als herbaris, les citacions de P. mixta de la part oriental dels Pirineus,
149 Butlletí ICHN 85 (3), 2021 NOTA BREU SOBRE LA VARIABILITAT DE PEDICULARIS PYRENAICA I LA PRESENCIA DE PEDICULARIS MIXTA
Figura
NOTA BREU
per comprovar si corresponen realment a aquesta planta o bé a expressions de la variabilitat de P. pyrenaica. Això hauria de permetre, per una banda, precisar la distribució i també la magnitud de les poblacions de P. mixta, i alhora fer encara més evident la remarcable variabilitat morfològica i ecològica de P. pyrenaica
Agraïments
Als conservadors-es dels herbaris BCN i BC per facilitarnos les consultes i la gestió dels materials d’herbari.
Bibliografia
Atlas de la Flora dels Pirineus 2021a. Disponible a http://www. atlasflorapyrenaea.eu [Data de consulta: 15 de juliol de 2021]
Atlas de la Flora dels Pirineus (primera versió) 2021b. Disponible a http://www.florapyrenaea.com [Data de consulta: 15 de juliol de 2021]
Biodiversdad Virtual 2021. Disponible a http://www. biodiversidadvirtual.org [Data de consulta: 15 de juliol de 2021]
Bolòs, O. & Vigo, J. 1996. Flora dels Països Catalans 3. Barcino, Barcelona. 1230 p.
Bolòs, O., Vigo, J., Masalles, R. M. & Ninot, J. M. 2005. Flora manual dels Països catalans. Pòrtic, Barcelona. 1310 p.
Carrillo, E., Mercadé, A., Ninot, J. M., Carreras, J., Ferré, A. & Font, X. 2008. Check-list i Llista vermella de la flora d’Andorra. Centre d’estudis de la neu i de la muntanya d’Andorra (CENMA) de l’Institut d’Estudis Andorrans (IEA). Ministeri de Turisme i Medi Ambient - Departament de Medi Ambient. Andorra. 488 p
Flor Catalana 2021. Disponible a http://floracatalana.cat [Data de consulta:15 de juliol de 2021]
Grenier, J. C. M. 1853. Pedicularis mixta Gren. P. 279-281.In: Schulz, F .W. Archives de la flore de France et Allemagne. Bitche, Haguenau & Deux Ponts. P. 279-281.
Grenier, J. C .M. & Godron, D.A. 1853. Flore de France, ou description des plantes qui croissent naturellement en France et en Corse, vol. 2. Paris, J.B. Bailliere.
Soriano, I. 2009. Pedicularis L. P. 512-530. In: Benedí, C., Rico, E., Güemes, J. & Herrero, A. (eds.), Flora iberica, vol. 13 (Scrophulariaceae, Plantaginaceae). Real Jardín Botánico-CSIC. Madrid. 677 p.
Soriano, I. 2013. El gènere Pedicularis (Orobanchaceae) al Parc i les àrees veïnes.VIII Jornades sobre Recerca al Parc Nacional d’Aigüestortes y Estany de Sant Maurici, Boí, 17-19 d’octubre de 2012. P. 131-137.
Soriano, I. 2018. Híbridos pirenaicos de Pedicularis (Orobanchaceae). Collectanea Botanica (Barcelona) 37, e011. https://doi. org/10.3989/collectbot.2018.v37.011
Tela Botanica 2021. Disponible a http://www.tela-botanica.org [Data de consulta: 15 de juliol de 2021]
Tison, J.-M., Foucault, B. D. & Guiol, F. 2014. Flora Gallica: flore de France. Biotope Ed. Mèze. 1196 p.
Vigo, J. 1983. El poblament vegetal de la vall de Ribes. I. Generalitats. Catàleg florístic. Acta Botanica Barcinonensia 35: 1-793.
Annex 1. Materials estudiats de Pedicularis pyrenaica de calzes i bràctees pubescents, atribuibles a la var. pilifera
BARCELONA. Bagà, capçalera de la Vall de Gréixer, pastures mesòfiles subalpines, 31TDG08, + 1.900 m, 6.VII.1986, I. Soriano s.n. (BCN 9853). GIRONA. Meranges, estany sec (Malniu), molleres, 31TDH0002, 2.100 m, 30.VII.1998, I. Soriano 98150, R. Perarnau (BCN 10767). Meranges, devesa d’en Sala, prados mesohigrófilos, 31T CH983009, 1.735 m, 28.VI.2010, I. Soriano & al. PMu18 (BCN 78825). LLEIDA. Espot, La Mosquera, in pratis humidis, 1.900 m, 13.VII.1934, W. Rothmaler (BC 78095). Alins, pla de Boet, humedales acidófilos y orillas de riachuelos, 31T CH6819, 1.850 m, 5.VII.1999, I. Soriano 99144 (BCN 10818). Alins, pla d’Arcalís, humedales y prados higrófilos, 31TCH6819, 2.000 m, 5.VII.1999, I. Soriano 99138 (BCN 10819). Alins, pla de Baiau, molleres, 31TCH61, 2.180 m, 3-5.VII.1985, L. Casanovas I102-107, 114 (BCN 11684). ANDORRA. Arcalís, rierol, 2.200 m, 16.VI.1993, E. Del Castillo (BC 913619). Pleta de Siscaró, 2.200 m, 29.VII.1983, L. Casanovas I005 (BCN 10801). El Serrat d’Andorra, 31T CH81, 15.VI.1979, M. Perdigó (BCN 10843). Pla de Sorteny, 31T CH81, 1.750 m, 7.VIII.1948, P. Montserrat (BCN 13445). PIRINEUS ORIENTALS. Palau de Cerdanya, Clot de Malniu, mollera, 31T DG164910, 1.990 m, 11.VII.2009, V. González (BCN 111240). Porté-Puimorens, vora el rec d’en Garcia, marges del riu, 31T DH048148, 2.294 m, 6.VIII.2020, I. Soriano 3512 (BCN 161364). Porté-Puimorens, Vall de cortal Rossó, herbassars higròfils i marges de rierols, silici, 31T DH043123, 2.059 m, 10.VII.2021, I. Soriano 3527 (BCN s/n). Estany de la Bullosa, la Tet, 31T DH11, 2.040 m, 5.VIII.1985, L. Casanovas I198 (BCN 10792). Pélouses humides, au dessus du Lac des Bouillouses, 2.000 m env., 21.VII.1962, H. Bouby 5940 (P 230598 foto!) [sub× P. pyrenaica Gay. × P. mixta Grenier]. Estany de Vallserra, 31T DH21, 1.770 m, 14.VII.2007, R. Balada (BCN 143151). Bosc de Barres, molleres, 31T DH21, 1.700 m, 3.VIII.1985, L. Casanovas I187(BCN 10762). Près de Mt. Louis, m. Rivière [sub P. mixta Gren.] (P 00403949 foto?). Massís de Mades, coma de Pontells, marges de rierols amb molleres i torberes d’esfagnes, 31T DH322209, 1.930 m, 12.VIII.2020, I. Soriano 3514 (BCN 161365). Noedes, entre el refugi de la font de la perdiu i la Pelada, prat subalpí, 31T DH347212, 2.300 m, 4.VII.2009, R. Balada & T. Buira (BCN 94058). Estany negre, 31T DH32, 2.083 m, 12.VIII.2008, R. Balada (BCN 143337).
150 Butlletí ICHN 85 (3), 2021
I. SORIANO
Aportacions a la flora aŀlòctona
catalana
Pere Aymerich* & Llorenç Sáez**,***
* C. Barcelona, 29. ES 08600 Berga, Barcelona.
** Sistemàtica i Evolució de Vascular Plants (UAB) – Unitat Associada al CSIC. Departament Biologia Animal, Vegetal i Ecologia. Facultat de Biociències. Universitat Autònoma de Barcelona. ES 08193 Bellaterra, Barcelona.
*** Societat d’Història Natural de les Illes Balears. c/ Margarida Xirgu 16. ES 07003 Palma de Mallorca, Illes Balears.
Autor per a la correspondència: Pere Aymerich. A/e: pere_aymerich@yahoo.es
Rebut: 30.08.2021; Acceptat: 20.09.2021; Publicat: 30.09.2021
Resum
S’aporten dades sobre plantes aŀlòctones, trobades creixent de manera espontània, noves per a Catalunya o poc citades. Quinze tàxons es citen per primera vegada a Catalunya: Abies cephalonica, Abies nordmanniana, Allium tuberosum, Cotoneaster zabelii, Erodium ×variabile, Jasminum mesnyii, Larix decidua, Limonium caesium, Malus toringo, Pistacia vera, Puschkinia scilloides, Pyracantha crenulata, Robinia viscosa, Thuja plicata i Viburnum rhytidophyllum. Tres altres plantes ja havien estat indicades, però s’havien confós amb tàxons pròxims: Gaillardia ×grandiflora, Juglans ×intermedia i Salvia lavandulifolia subsp. gallica. Es confirma també la naturalització local de Berberis julianae, Cotoneaster divaricatus i Elaeagnus angustifolia, tres espècies xineses que fins ara es coneixen com a al·lòctones casuals.
Paraules clau: flora, plantes aŀlòctones, península Ibèrica, regió Mediterrània.
Abstract
Notes about alien flora in Catalonia
We provide new data about alien plants new or poorly known in Catalonia. Fifteen taxa are first reported from Catalonia: Abies cephalonica, Abies nordmanniana, Allium tuberosum, Cotoneaster zabelii, Erodium ×variabile, Jasminum mesnyii, Larix decidua, Limonium caesium, Malus toringo, Pistacia vera, Puschkinia scilloides, Pyracantha crenulata, Robinia viscosa, Thuja plicata and Viburnum rhytidophyllum. Three other plants were already known, but were confused with similar taxa: Gaillardia ×grandiflora, Juglans ×intermedia and Salvia lavandulifolia subsp. gallica. Local naturalisation of Berberis julianae, Cotoneaster divaricatus and Elaeagnus angustifolia –three species previously known as casual aliens– is confirmed.
Key words: flora, alien plants, Iberian Peninsula, Mediterranean region.
Introducció
Aquest article és una nova aportació de dades referides al coneixement de la flora aŀlòctona, grup on la biodiversitat va augmentant de forma sostinguda i creixent a Catalunya, i l’actualització del qual ha generat un nombre notable de publicacions en la darrera dècada. Aquest augment és degut a factors diversos (Aymerich & Sáez, 2019), però un dels més importants és el progressiu escapament de plantes cultivades com a ornamentals als jardins, com són la major part de tàxons que es citen en aquest treball. Aquest fenomen és creixent a causa del procés de suburbanització que experimenta bona part del territori, en especial a la franja marítima.
Les dades que es presenten es refereixen a localitats de plantes aŀlòctones, trobades creixent de manera espontània, i que serien noves o molt rares a Catalunya. Els tàxons són presentats en ordre alfabètic. Les citacions de cada tàxon inclouen aquesta informació: localització (per aquest ordre:
comarca, municipi, lloc i coordenades UTM 1×1 km amb sistema de referència ETRS89), altitud, hàbitat, recompte o estimació dels individus observats (sempre que ha estat possible), data o dates de l’observació i observador (especificant, si s’escau, si es conserven plecs testimoni als herbaris dels autors o en altres coŀleccions). Les citacions es complementen amb comentaris diversos sobre l’origen de la planta, el coneixement previ que se’n tenia a Catalunya o en altres àmbits geogràfics, el grau de naturalització observat i altres aspectes que poden resultar d’interès.
Abies cephalonica Loudon
Berguedà: Puig-reig, Colònia Prat, DG0749, 405-415 m, bardisses arbrades i jardí antic semiabandonat, 2 individus grossos (6-10 m) en bardisses i 10-15 juvenils (1-3 m) en l’àmbit del jardí, 27-I-2021 (P. Aymerich, herb. pers.).
Avet mediterrani endèmic de les muntanyes de Grècia i Macedònia, que en altres zones d’Europa es cultiva en jardi-
151 Butlletí ICHN 85 (3), 2021
FLORA ET FAUNA
GEA,
GEA, FLORA ET FAUNA
Butlletí de la Institució Catalana d’Història Natural, 85 (3): 151-162. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition) DOI: 10.2436.20.1502.01.88
GEA, FLORA ET FAUNA
neria i també, rarament, com a arbre forestal. La seva aparició com a planta aŀlòctona casual a l’entorn d’aquests cultius és freqüent, i també ha estat indicat com a naturalitzat en diversos sectors de la península Itàlica i de l’illa de Sardenya (Galasso et al., 2018; Baccheta et al., 2019). Les observacions de Puig-reig són les primeres de Catalunya com a planta escapada i provenen d’un gran jardí creat a primers del segle XX, actualment gairebé abandonat.
Abies nordmanniana (Steven) Spach subsp. nordmanniana
OsOna: Espinelves, riba dreta de la riera d’Espinelves, al nord de Can Botifarra, DG5234, 740 m, desenes d’exemplars juvenils dins l’àmbit d’una plantació d’avets, 2-II-2021 (P. Aymerich, herb. pers.).
Tàxon propi de les muntanyes que envolten per l’est la costa de la mar Negra, cultivat en jardineria, com a arbre de Nadal i rarament en plantacions forestals. A Catalunya es cultiva sobretot com a arbre de Nadal, i de fet és l’espècie més freqüent per aquesta finalitat. A Espinelves es van observar plançons relativament nombrosos dins l’àmbit d’una plantació poc cuidada. En altres llocs de l’àrea GuilleriesMontseny –i també puntualment a l’alt Berguedà– hem vist plantacions d’aquest avet, però sense constatar-hi la presència d’individus juvenils espontanis.
Abies pinsapo Boiss.
Berguedà: Castellar de n’Hug, Clot del Moro, DG1579, 945970 m, boscos clars en l’entorn d’una antiga zona enjardinada abandonada fa dècades, uns 25 individus juvenils (alçades màximes 2-4 m), febrer de 2021 (P. Aymerich); la Pobla de Lillet, bosc de Ventallola, DG1576, 920-940 m, plantació de coníferes, 150-200 joves, en general de menys de 0,5 m, gener de 2021 (P. Aymerich).
Noves dades sobre la reproducció espontània d’aquest avet mediterrani a l’alt Berguedà, on ja havia estat documentat en tres indrets més (Aymerich, 2013, 2015). Tot i la relativa abundància que pot assolir localment, per ara no s’ha verificat una veritable naturalització. En tots dos llocs s’han observat menys de 10 exemplars d’Abies pinsapo plantats, que són l’origen dels nuclis aŀlòctons. Al bosc de Ventallola, la freqüència de peus petits sembla relacionada amb un abandonament recent de la pastura de vaques al sotabosc.
Allium tuberosum Rottler
OsOna: Oristà, la Torre d’Oristà, cap al carrer de Baix, DG2145, 550 m, herbassars en una parceŀla sense cultivar adjacent a un jardí i hort, 5-IX-2020 (P. Aymerich, herb. pers.).
Aquest all originari de la Xina es coneix bàsicament com a planta cultivada o naturalitzada, i aparentment tindria una sola població salvatge (Jiemei & Kamelin, 2020). Generalment es cultiva com a verdura, però a Catalunya s’utilitza sobretot com a ornamental. A la Torre d’Oristà vam observar-ne quatre o cinc grups clonals en herbassars ruderals adjacents a un jardí rural, des del qual s’havia escapat aquesta planta; a més de forma espontània, dins l’àmbit del jardí i als marges d’un hort. Aquesta espècie fa llavors fèrtils que
germinen amb facilitat, cosa que n’afavoreix la dispersió. No citada fins ara de Catalunya com a planta aŀlòctona, però és comuna com a naturalitzada a l’est d’Àsia i a Europa s’ha citat com a mínim a Itàlia (Galasso et al., 2018). En altres països europeus hi ha citacions d’un tàxon pròxim i que sovint ha estat considerat conspecífic, A. ramosum L., algunes de les quals podrien correspondre també a A. tuberosum. Les plantes que hem observat corresponen a A. tuberosum per les fulles planes i la línia medial dels tèpals verdosa, mentre que A. ramosum tindria fulles fistuloses i línia rogenca.
Asparagus asparagoides (L.) Druce
Vallès Occidental: Sant Cugat del Vallès, torrent de Can Badal, DF1990, 95 m, talussos ombrívols del torrent, 2 individus, 9-V-2016 (L. Sáez).
Espècie originària d’Àfrica del Sud, que a Catalunya havia estat indicada ja de Blanes, la Selva (Aymerich, 2015) i de diversos punts del Maresme (Guardiola & Petit, 2020). Aparentment, aquesta seria la primera citació formal al massís de Collserola i l’entorn de Barcelona, tot i que la seva presència es coneixia per algunes fotos penjades en línia i sense indicació de localitat concreta.
Berberis julianae C.K. Schneid.
MOianès: Sant Quirze Safaja, fondal adjacent a la font del Solà, DG2820, 615-640 m, bosc i clarianes, mínim 40 individus, 28-X-2020 (P. Aymerich). Figura 1a.
Espècie originària del centre i el sud de la Xina, que a Europa és coneguda com a aŀlòctona casual o rarament naturalitzada (Stace, 2010; Pysek et al., 2012; Galasso et al., 2018; Verloove, 2020). A Catalunya es va detectar recentment en dues localitats del Ripollès, en una de les quals es va apuntar que hi havia indicis de naturalització incipient (Aymerich, 2020a). La nova localitat del Moianès és la primera en què s’ha verificat l’existència d’una població naturalitzada, formada per unes poques desenes d’individus escampats per una superfície d’1 ha, amb una estructura de mides heterogènia i amb evidències de reproducció recent per llavor.
Cedrus atlantica (Endl.) Carrière
Berguedà: Castellar de n’Hug, Erols de Rus, DG1380, 12601340 m., entorn de plantacions antigues de coníferes, 29-I2021 (P. Aymerich); OsOna: Espinelves, vessants obacs sobre la riera d’Espinelves, des del turó de les Lloberes fins a la carretera de Viladrau, DG5035-5036-5134-5135, 680-750 m, entorn de plantacions de coníferes actives, 2-II-2021 (P. Aymerich).
Espècie originària del nord d’Àfrica que es reprodueix amb relativa facilitat dins i al voltant de plantacions forestals. Ha estat molt poc citada a Catalunya com a planta escapada, a l’Alt Empordà, Berguedà, Osona i Ripollès (Casasayas, 1989, sub Cedrus libani subsp. atlantica; Aymerich, 2013) i, sense precisar localitzacions, en l’àmbit del massís del Montseny (Sáez et al., 2017). Dues de les citacions prèvies corresponen a localitats pròximes a les noves que s’aporten: Viladrau (Casasayas, 1989) i la Pobla de Lillet (Aymerich, 2013).
152 Butlletí ICHN 85 (3), 2021
P AyMERICH & L SáEz



En totes dues localitats hi ha de l’ordre de centenars d’individus joves, la majoria, i aparentment cap planta espontània no ha assolit l’estadi reproductor.
Cedrus deodara (G. Don) Loudon
Berguedà: Castellar de n’Hug, el Clot del Moro, al voltant de les ruïnes del Xalet i de l’ermita de Sant Jaume, DG1579, 950-970 m, arbredes mixtes dominades per pi roig, en una zona semiedificada abandonada fa dècades, 20-30 individus, 5-I-2021 (P. Aymerich); Gironella, Viladomiu Nou, vessant entre les cases i la riera de Clarà, DG0750, 430-445 m, arbredes i bardisses, menys de 10 individus, 27-I-2021 (P. Aymerich); Puig-reig, Colònia Prat, DG0749, 405-415 m, bardisses
arbrades i jardí antic abandonat, 20-30-individus, 27-I-2021 (P. Aymerich); ripOllès: Camprodon, perifèria oest del nucli urbà, al costat de l’avinguda de Maristany, DG4785, 960 m, fissures d’un mur en una zona semienjardinada, individu solitari, 25-VI-2021 (P. Aymerich).
Arbre procedent de l’Himàlaia occidental, que es cultiva sovint en jardineria i que, a Catalunya, només havia estat indicat com a escapat per Casasayas (1989), en dues localitats d’Osona i la Selva. Amb aquesta nota aportem tres localitats de la conca mitjana i alta del Llobregat, en les quals es dispersa a partir d’arbres plantats en jardins a primers del segle XX, i una del Ripollès. Tots els individus no cultivats que hem observat són juvenils no reproductors, el més alt d’uns
153 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA APORTACIONS A LA FLORA AŀLÒCTONA CATALANA
Figura 1. a) Berberis julianae; b) Cotoneaster divaricatus; c) Cotoneaster zabelii. Fotografies: Pere Aymerich.
a c b
GEA, FLORA ET FAUNA
6 m, però generalment de menys d’1 m. Els arbres de les tres localitats berguedanes havien estat citats erròniament com a C. atlantica a Aymerich (2013), determinació que hem corregit amb una revisió recent.
Cotoneaster apiculatus Rehder & E.H. Wilson [incl. C. hjelmqvistii Flinck & B. Hylmö]
MOianès: Sant Quirze Safaja, vessants adjacents als torrents del Gatell i de la Teuleria, a la perifèria oest de la zona urbanitzada, DG2819-2919-2820, 610-630 m, boscos i clarianes, mínim 200 individus, 23-X-2020; Sant Quirze Safaja, urbanització Solà del Boix, DG2920, 620-650 m, retalls de vegetació natural entre parceŀles edificades i enjardinades, freqüent, en conjunt desenes d’individus, 28-X-2020; Sant Quirze Safaja, fondal adjacent a la font del Solà, DG2820, 620-640 m, boscos i clarianes, desenes d’individus, 28-X-2020; Sant Quirze Safaja, baga del Cerdà, DG2919, 660-680 m, marges de bosc, 9 individus escampats en una superfície gran, 28-X-2020; Sant Quirze Safaja, pla del Badó, entre les cases de Poses i del Badó, parceŀla arbrada i sense edificar en una àrea semiurbanitzada, DG2919, 710 m, més de 100 individus, 28-X-2020 (P. Aymerich, herb. pers.); ripOllès: Sant Joan de les Abadesses, vessant nord-oest del turó de les Tres Creus, a la perifèria sud del nucli urbà, DG4075, 785 m, arbreda mixta de caducifolis, uns 10 individus, 2-XI-2020 (P. Aymerich).
Aquesta espècie, que sovint es confon amb C. horizontalis, havia estat indicada fins ara de quatre localitats catalanes, al Bages, Berguedà, Cerdanya i Ripollès (Aymerich, 2017, 2019, 2020), en les quals s’havien vist com a molt dues desenes d’individus. A Sant Quirze Safaja està plenament naturalitzat, ha assolit una abundància i expansió que són insòlites al nostre territori i, a escala local, es pot qualificar gairebé com a espècie invasora. Només al sector més pròxim al nucli urbà de Sant Quirze, s’ha estimat que l’espècie és present en una superfície d’unes 20 ha. La població total del municipi és desconeguda, però es considera probable que volti el miler d’individus.
Cotoneaster divaricatus Rehder & E.H. Wilson
MOianès: Sant Quirze Safaja, fondal adjacent a la font del Solà, DG2820, 620-680 m, boscos i clarianes, mínim 175 individus, 28-X-2020 (Figura 1b); Sant Quirze Safaja, baga del torrent de la Teuleria, sota Cal Colomer, DG2820, 610 m, bosc clar, un individu, 23-X-2020 (P. Aymerich, herb. pers.).
Tàxon molt rar com a escapat a Catalunya, citat prèviament de tres localitats del Bages i del Ripollès, amb 1-5 individus en cadascuna (Aymerich, 2016, 2020a). Les dades de Sant Quirze són les primeres que es refereixen a una població plenament naturalitzada i aparentment en expansió.
Cotoneaster franchetii Bois
MOianès: Sant Quirze Safaja, vessants adjacents als torrents del Gatell i de la Teuleria, a la perifèria oest de la zona urbanitzada, DG2819-2820, 610-640 m, boscos i clarianes, 8 individus, 23-X-2020 (P. Aymerich, herb. pers.); Sant Quirze Safaja, baga del Cerdà, DG2919, 670 m, un individu, 28-
X-2020 (P. Aymerich); ripOllès: Camprodon, perifèria oest del nucli urbà, al vessant sobre l’avinguda de Maristany, DG4785, 960 m, marge d’una de bosc prop de cases, individu reproductor solitari, 2-XI-2020 (P. Aymerich).
Espècie freqüent com a escapada a l’Europa temperada, però rara a Catalunya, citada fins ara d’altres localitats del Moianès i del Ripollès i d’una de la Cerdanya, on es coneix l’única població naturalitzada (Aymerich, 2020a). A Sant Quirze Safaja hi ha indicis d’un procés de naturalització incipient, ja que s’han observat individus molt dispersos en una àrea forestal extensa, en la qual és de detecció difícil i podria estar subestimat.
Cotoneaster horizontalis Decne.
MOianès: Sant Quirze Safaja, vessants adjacents als torrents del Gatell i de la Teuleria, a la perifèria oest de la zona urbanitzada, DG2819-2919-2820, 605-625 m, població gran estimada en 300-500 individus, boscos i clarianes, 23-X-2020 (P. Aymerich); Sant Quirze Safaja, baga del Cerdà, DG2919, 670-690 m, 7 individus, 28-X-2020 (P. Aymerich).
Tot i ser rar, aquest és un dels Cotoneaster asiàtics més observat a Catalunya, i actualment ja s’ha citat de la major part de les comarques interiors del centre-est del país, en un triangle que va del Baix Llobregat a la Cerdanya i a la Garrotxa (cf. Aymerich, 2020a). Al Moianès només havia estat citat de Moià, i un sol individu (Aymerich, 202a). Les dades que s’aporten de Sant Quirze Safaja són interessants perquè és la localitat catalana on, amb gran diferència, s’ha observat una població naturalitzada més nombrosa i extensa.
Cotoneaster integrifolius (Roxburgh) G. Klotz
MOianès: Sant Quirze Safaja, baga sobre el torrent del Gatell, a la perifèria oest de la zona urbana (carrer d’en Guitert), DG2919, 625 m, talús al marge d’un bosc de roures, 5-7 individus, 23-X-2020 (P. Aymerich, herb. pers.).
Espècie que fins ara només havia estat citada una vegada a Catalunya, d’una localitat de la Cerdanya en què es va observar un sol individu (Aymerich, 2020a).
Cotoneaster salicifolius Franch.
MOianès: Sant Quirze Safaja, perifèria oest de la zona urbana, sota cal Colomer, DG2820, 615 m, bosc molt antropitzat, un individu, 23-X-2020 (P. Aymerich).
A Catalunya, aquesta espècie va ser citada per primera vegada com a aŀlòctona al Moianès, concretament a Castellterçol (Mercadé, 2016). Més endavant es va trobar en tres localitats de la Cerdanya i del Ripollès (Aymerich, 2020a). En tots aquests llocs s’han observat molt pocs individus, bé que en algunes localitats dels Pirineus hi ha indicis d’una naturalització incipient.
Cotoneaster zabelii C.K. Schneid.
MOianès: Sant Quirze Safaja, fondal adjacent a la font del Solà, DG2820, 620-640 m, bosc clar, uns 20 individus, 28-X2020 (P. Aymerich, herb. pers.). Figura 1c.
154 Butlletí ICHN 85 (3), 2021
P AyMERICH & L SáEz
Espècie no citada fins ara, segons la informació disponible, a Catalunya ni al sud d’Europa. Sí que és coneguda com a rarament naturalitzada a Gran Bretanya (Stace, 2010) i com a aŀlòctona casual a Alemanya i Bèlgica (Dickoré & Kasperek, 2010; Verloove, 2020). A Sant Quirze presenta indicis de naturalització incipient, però per ara sembla preferible considerar-la com a casual.
La distinció entre C. zabelii i C. fangianus T.T. Yü és controvertida. Tots dos tàxons es diferencien bé d’altres espècies amb fulles similars, de mida petita-mitjana i tomentoses, perquè tenen els fruits penjants. En canvi, la separació entre els dos ha estat interpretada de formes diverses i no sembla que encara s’hagi arribat a un consens satisfactori. Dickoré & Kasperek (2010) simplement consideren C. fangianus com a un sinònim de C. dielsianus E. Pritz., espècie que no té els fruits pènduls. Lingdi & Brach (2003) sí que reconeixen C. zabelii i C. fangianus com a espècies diferents, criteri adoptat també per Stace (2010). Les plantes de Sant Quitze Safaja concorden amb els caràcters que atribueixen a C. zabelii els treballs anteriors (fruits pènduls de 7-10 mm, en general obovoides, amb 2 pirens, 3-10 flors, fulles eŀlíptiques o ovades, típicament de 15-30 mm, obtuses o apiculades). Per contra, Fryer & Hylmö (2009) fan una interpretació diferent i atribueixen a C. zabelii típicament fruits de 10-12 mm amb 3 pirens i fulles de 30- 40 mm, mentre que C. fangianus tindria fruits més petits amb 2 pirens i fulles de 20-35 mm. Amb aquest segon criteri, les plantes de Sant Quirze es podrien atribuir a C. fangianus. Considerant aquesta incertesa, hem preferit assignar la població del Moianès a C. zabelii, perquè és el criteri adoptat per la flora de referència al territori d’origen d’aquesta espècie (Lingdi & Brach, 2003), tot i que no es pot excloure que noves aportacions taxonòmiques indiquin que el tractament adequat és el proposat per Fryer & Hylmö (2009).
Elaeagnus pungens Thunb.
Vallès Occidental: Sant Cugat del Vallès, serra de Collserola, torrent de la Salamandra, DF2686, 20 m, marge forestal humit, 26-X-2020 (L. Sáez, herb. pers.).
Espècie de jardineria provinent de l’est d’Àsia, que a Catalunya s’havia citat com a escapada de dues localitats del Ripollès i una d’Osona (Aymerich, 2020a), amb un nombre baix d’individus. A la localitat de Collserola E. pungens formava dos rodals densos més o menys lineals, amb unes dimensions aproximades de 12×3 i 5×2 m, situats entre un camí i el curs del torrent. En la data d’observació presentava flors abundants. Localment es considera naturalitzada i aparentment en expansió.
Erodium ×variabile A.C. Leslie
Vallès Occidental: Sant Cugat del Vallès, voltants de l’aparcament de l’Hospital General, DF2092, 125 m, marge herbós humit, IV-2018 (L. Sáez).
Planta generada en jardineria i d’origen discutit, tot i que se sol acceptar que és un híbrid artificial entre Erodium corsicum i E. reichardii, endemismes respectivament de les illes
de Còrsega i Mallorca. Actualment n’existeixen nombroses varietats de cultiu (International Geraniaceae Group, 2021). A Sant Cugat del Vallès es va trobar un únic exemplar en flor, que podria correspondre al cultivar «Roseum», amb pètals rosats i venació porpra o violàcia. Posteriors prospeccions de la zona no van permetre retrobar aquesta planta, la presència de la qual devia ser esporàdica. Erodium ×variabile actualment es fa servir força en jardineria urbana (Frey & Moretti, 2019; Kaltsidia et al., 2020), de manera que és probable un increment dels escapaments en hàbitats urbans.
Gaillardia ×grandiflora Van Houtte [G. aristata Pursh × G. pulchella Foug.]
cerdanya: Prats i Sansor, urbanització del Pla, DG0591, 1125 m, marge de camí, individu solitari, 21-IX-2020 (P. Aymerich); ripOllès: Ripoll, part alta de la zona semiurbanitzada de Ciutat Jardí, DG3371, 730 m, vores de camí i fissures de paviment, 4 individus, 8-VII-2020 (P. Aymerich). Figura 2a,b.
Híbrid de jardineria entre dues espècies de l’oest d’Amèrica del nord, que es cultiva sovint com a ornamental i que ocasionalment s’escapa. Tot i que sovint es comercialitzen amb el nom d’alguna de les espècies parentals (especialment G. aristata), les plantes hortícoles són habitualment híbrids (Strother, 2006) i aquestes són les que es presenten com a aŀlòctones a Europa (Stace, 2010; Verloove, 2020). L’aspecte general de les plantes observades a les localitats indicades és més similar al de G. aristata, però se’n separen per caràcters com els pecíols curts de les fulles caulinars. Citacions prèvies de G. aristata a Catalunya (Casasayas, 1989) s’han de referir amb tota probabilitat a G. ×grandiflora
Hypericum calycinum L.
MOianès: Sant Quirze Safaja, perifèria nord-est de la urbanització del Pla de Badó, cap al Grony, DG2919, 700 m, franja de matollar proper a cases que s’estassa de forma regular, taca clonal d’uns 20 m2, 28-X-2020 (P. Aymerich).
Aquesta espècie originària de la Mediterrània oriental es cultiva molt sovint en jardineria, però s’escapa rarament i a Catalunya només en coneixem quatre citacions prèvies, dues d’Osona (Casasayas, 1989) i dues del Berguedà (Aymerich, 2014). Sembla que no es multiplica per llavor, de manera que la dispersió sol ser deguda a l’abocament de restes de jardineria i pot formar poblacions puntuals que es mantenen per creixement clonal.
Jasminum mesnyi Hance
Vallès Occidental: Sant Cugat del Vallès, torrent de Can Cabassa, DF2091, 100 m, talussos rocosos del marge del torrent, 1 individu, 9-V-2016 (L. Sáez).
Espècie originària de la Xina que es cultiva en jardineria i de la qual a Catalunya no es tenia cap dada publicada referida a plantes clarament escapades. Al Banc de Dades de la Biodiversitat de Catalunya s’indica del barri del Carmel a Barcelona (DF2985, 186 m), sobre la base de recoŀlecci-
155 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
APORTACIONS A LA FLORA AŀLÒCTONA CATALANA




ons realitzades per C. Gómez (BC 879685 i BCN 124802), però desconeixem si es tracta de plantes realment escapades o de restes de cultiu. Com a persistent de cultiu i amb creixement vegetatiu important també es coneix d’un gran jardí antic semiabandonat de la Colònia Prat (Puig-reig, Berguedà) DG0749, 405-410 m (P. Aymerich, obs. pers.).
Juglans ×intermedia Carrière
Berguedà: Olvan, entorn de la riera de la Riba a l’alçada de

Cal Llop, DG0855, 475-490 m, dins l’àmbit d’una plantació de nogueres semiabandonada i a les bardisses adjacents, desenes d’individus juvenils, IX-2020 (P. Aymerich). Figura 2c.
Fa uns anys (Aymerich, 2013) es va assenyalar la presència en aquesta localitat de peus joves espontanis de Juglans nigra L a l’entorn d’una plantació fustanera d’aquest arbre americà. Una revisió recent d’aquestes plantes ha mostrat que no es tracta de J. nigra, sinó de nogueres híbrides entre una espècie nord-americana i l’espècie euroasiàtica J.
156 Butlletí ICHN 85 (3), 2021
FLORA ET FAUNA
GEA,
a b c e d
Figura 2. a, b) Gaillardia ×grandiflora; c) Juglans ×intermedia; d) Malus toringo; e) Nicandra physalodes. Fotografies: Pere Aymerich.
P AyMERICH & L SáEz
regia L. Les característiques d’aquestes nogueres cultivades i subespontànies es corresponen amb les del cultivar comercial MJ209, originat en vivers a França i sovint cultivat al sud d’Europa, on s’adapta més bé que J. nigra (Aletà et al., 2003; Fernández-Moya et al., 2020). En general (Gonin et al., 2013) es considera que les espècies parentals de l’híbrid MJ209 són J. regia i J. major (Torr.) A. Heller, una noguera americana amb àrea principal a les muntanyes de l’oest mexicà, però sembla que existeix alguna incertesa en la participació real de J. major. A causa d’aquesta incertesa, aquí assignem aquestes plantes que neixen espontàniament a J. ×intermedia en sentit ampli, un nom també incert (cf. http:// www.theplantlist.org) que se sol aplicar en silvicultura als híbrids entre J. nigra i J. regia, i per extensió també a altres nogueres híbrides euroamericanes.
Larix decidua Mill.
Berguedà: Castellar de n’Hug, Erols de Rus, DG1380, 1340 m, entorn de plantacions antigues de coníferes, 29-I-2021 (P. Aymerich).
Aquesta conífera, amb una àrea natural difosa per les muntanyes del centre d’Europa, havia estat indicada de Catalunya com a cultivada, però no en coneixem cap citació com a espècie si més no escapada. L’observació del Berguedà és anecdòtica i es refereix a un sol peu de mida petita (2 m) però ja reproductor, que creixia a uns quants metres dels dos únics individus i de mida gran (10-15 m) vistos en aquesta zona, un vessant de muntanya en què l’administració forestal va fer plantacions experimentals de diverses coníferes en les dècades de 1960 i 1970. Larix ×marschlinsii Coaz, un híbrid entre L. decidua i L. kaempferi (Lamb.) Carr., es planta força més sovint i a Catalunya mostra una naturalització incipient en una localitat del Ripollès (Aymerich, 2019).
Limonium caesium (Girard) Kuntze
tarragOnès: Roda de Berà, entre la platja dels Capellans i la Platja Llarga, CF7158, 5-15 m, roques i talussos marítims, 1-II-2020 (L. Sáez, herb. pers.).
Primera citació a Catalunya d’aquesta espècie endèmica del sud-est ibèric. El material recoŀlectat a Roda de Berà presenta espiguetes uniflores, de 5-6,2 mm de longitud, caràcters que permeten separar-lo de L. insigne (Coss.) Kuntze, que té espiguetes 2-3-flores i de 8-10 mm de longitud (Erben, 1993). Amb tota probabilitat, L. caesium es va plantar a la zona en el marc de treballs destinats a recuperar i estabilitzar la franja litoral. La població observada inclou uns 70 individus que es troben en roques i vessants marítims. Hem observat individus joves que colonitzen pendents i que no es corresponen amb la zona de plantació original. La població sembla haver resistit amb èxit els forts temporals del gener del 2020, que van afectar bona part de la costa meridional. A la zona, L. caesium creix juntament amb altres espècies mediterrànies, però també aŀlòctones a Catalunya, com és el cas de Pallenis maritima
Linaria ×versicolor (Jacq.) Chaz.
BarcelOnès: Barcelona, tram inferior del riu Besòs, DF3486,
5 m, herbassars ruderals, 3-V-2015, L. Gustamante (P. Aymerich, herb. pers.).
Tàxon llistat com a casual per Aymerich & Sáez (2019) per a la depressió de l’Ebre (en base a un material d’herbari recoŀlectat a la zona d’Igualada, leg. L. Garrido) i indicat amb dubtes per al sector litoral de Catalunya. L’estudi del material procedent de Barcelona permet confirmar que es tracta de L. ×versicolor. Aquesta espècie correspondria a formes ornamentals cultivades, que de vegades són comercialitzades sota el nom de «L. maroccana hort.» i que podrien tenir un origen híbrid entre L. maroccana Hook. f. i alguna espècie de la sect. Versicolores com ara L. bipartita (Vent.) Willd., L. gharbensis Battandier & Pitard, i L. incarnata (Vent.) Spreng. (vegeu Sáez, 2019).
Lonicera ligustrina Wall.
sOlsOnès: Guixers, Vilamantells, torrent de les Salines, CG8265, 1030 m, torrentera seca en una roureda, taca clonal d’uns 6 m2, 4-IV-2021 (P. Aymerich).
Arbust d’origen xinès, que a Europa es cultiva sovint en jardins i de vegades s’escapa. A Catalunya havia estat indicat de Berga i de Ripoll (Aymerich, 2020a). El nucli del Solsonès correspon a la var. yunnanensis Franch. [L. nitida E.H. Wilson]. Es trobava en un hàbitat natural i lluny de cases, i suposem que es va originar per l’abocament de restes de jardineria.
Mahonia ×decumbens Stace
MOianès: Sant Quirze Safaja, fondal adjacent a la font del Solà, DG2820, 620-640 m, bosc i clarianes, població nombrosa, probablement de més de 100 individus, 28-X-2020 (P. Aymerich).
Constatem la naturalització al Moianès d’aquest híbrid de jardineria [aparentment M. aquifolium (Pursh.) Nutt. × M. repens (Lindl.) G. Don]. Hi corresponen també gairebé totes les Mahonia escapades a Catalunya (Aymerich, 2019, 2020), territori en el qual en els darrers anys s’han anat detectant de forma creixent poblacions naturalitzades als boscos adjacents a zones semiurbanitzades de les comarques una mica humides. A més de la població naturalitzada de la font del Solà, a Sant Quirze n’hem observat individus isolats o petits grups (menys de 5) en diversos punts de la baga del Cerdà (DG2919) i de les vores del torrent de la Teuleria (DG2820). Al Moianès, M. aquifolium havia estat indicada per Mercadé (2016) com a cultivada i rarament subespontània a Sant Quirze Safaja, però es pot assumir que aquestes observacions es refereixen també a M. ×decumbens.
Malus toringo (Siebold) Siebold [M. sieboldii Rehder]
MOianès: Sant Quirze Safaja, baga del Cerdà, sota el pla de Badó, DG2919, 690 m, clariana de bosc al costat d’un camí i a 60-100 m de cases amb jardins, 2 individus petits, no reproductors, 28-X-2020 (P. Aymerich, herb. pers.). Figura 2d.
Primera citació a Catalunya, com a planta aŀlòctona escapada, d’aquesta espècie que, segons la informació disponi-
157 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
APORTACIONS
LA FLORA AŀLÒCTONA CATALANA
A
GEA, FLORA ET FAUNA
ble, tampoc no havia estat citada de regions pròximes del sud d’Europa. Es tracta d’un arbust o petit arbre de fulles petites i generalment trilobades, originari d’Àsia oriental (Xina, Corea i Japó), que es cultiva com a ornamental i també s’utilitza com a portaempelts de pomera (Malus domestica). S’ha naturalitzat a Amèrica del nord (Dickson, 2013) i a Europa és conegut com a espècie aŀlòctona (amb estatus divers, de vegades poc clar) en força països del centre i del nord, sobretot al voltant del mar Bàltic (Keil & Loos, 2005, Nielsen et al., 2019). Les plantes europees són referibles sovint a la var. sargentii (Rehder) Asami, que algunes vegades és tractada com a espècie (M. sargentii Rehder). La mida petita de les plantes trobades al Moianès i la manca de flors o fruits no permet avaluar amb fiabilitat si poden correspondre a aquesta varietat, però alguns dels caràcters observats (creixement clarament arbustiu, presència de nombroses espines) semblen més habituals en la var. sargentii que en els exemplars de M. toringo típics.
Muscari armeniacum Leichtlin
ripOllès: Vilallonga de Ter, el Catllar, DG4189, 1215 m, herbassar nitròfils prop de la casa, 30-III-2021 (P. Aymerich).
Aquesta espècie prové de l’àrea compresa entre la Mediterrània oriental i el Caucas, es cultiva en jardineria i es naturalitza sovint. Tot i això, a Catalunya sembla que hi ha una sola dada publicada, de Castellar de n’Hug, Berguedà (Aymerich, 2014), potser perquè ha estat confosa amb Muscari autòctons, principalment M. neglectum. Al Catllar hi vam observar un grup clonal d’uns 15 individus, 8 dels quals reproductors. En aquest mateix lloc s’han trobat altres plantes bulboses rares de jardineria (cf. Aymerich, 2019, 2020a), que persisteixen o es reprodueixen de forma limitada, però fins ara no s’ha verificat que cap estigui naturalitzada.
Myoporum insulare R. Br.
Baix eMpOrdà: Calonge, entre el cap Roig i el cap de les Penyes Blanques, EG0730, 15 m, roques marítimes silícies, 2-X-2016, M. Guardiola & L. Sáez (L. Sáez, herb. pers.).
Planta originària del sud d’Austràlia, d’estatus taxonòmic incert. A Catalunya va ser indicada per Paiva (2001) de l’àmbit provincial de Tarragona, sense més precisions, i se’n desconeixia cap localitat concreta. Al litoral del Baix Empordà es van observar dos exemplars adults ben desenvolupats, creixent en un vessant rocós situat entre una zona urbanitzada i la línia litoral.
Nicandra physalodes (L.) Gaertn.
Baix eMpOrdà: Colomers, sector dels Recs i el pla de Roda, DG9758-9759, 25 m, camps de blat de moro, 4 individus dispersos en una àrea extensa, 15-X-2020 (P. Aymerich). Figura 2e.
Solanàcia originària d’Amèrica del sud que actualment es presenta com a planta aŀlòctona en moltes regions tropicals, subtropicals i temperades del planeta. A Catalunya només ens consten dues citacions modernes publicades, al Maresme (Casasayas, 1989) i al Montsià (Senar & Cardero, 2019) i, a més, hi ha dos plecs d’herbari de la fi del segle XIX (Ca-
sasayas, 1989), els quals resulta incert que corresponguin a plantes no cultivades. La presència a la localitat del Baix Empordà sembla atribuïble a la barreja de llavors de Nicandra i altres males herbes en alguna partida de grans de blat de moro, ja que els mateixos camps en què es va observar tenien una gran població d’una altra solanàcia americana, Physalis angulata L. (Aymerich, 2020b).
Pistacia vera L.
Segrià: Maials, el Catllar, 9083-9183, 315-330 m, talussos en marges de camps, en un sector en què hi ha camps d’aquesta espècie, 2 individus juvenils, 7-VII-2021 (P. Aymerich). Figura 3a.
Les darreres dècades s’han fet en aquest sector del Segrià algunes plantacions comercials de festuc, originari d’Àsia central. Tot i la relativa extensió del conreu, sembla que l’escapament de plantes fora dels camps és molt anecdòtica, perquè tan sols s’han observat dos individus i encara en ambients agraris, sense penetrar a les taques de vegetació natural (pinedes i brolles) adjacents, que probablement resulten massa seques per aquesta espècie.
Puschkinia scilloides Adams
ripOllès: Vilallonga de Ter, el Catllar, DG4189, 1215 m, herbassar nitròfils prop de la casa, 4 individus reproductors, 30-III-2021 (P. Aymerich, herb. pers.). Figura 3b.
Petita asparagàcia originària de les muntanyes compreses entre la serralada del Caucas i del Líban, que es cultiva com a ornamental. Ha estat indicada com a espècie aŀlòctona en alguns països del nord i centre d’Europa, per exemple com a casual a Txèquia (Pysek et al., 2012) i sense categoria definida a Suècia (Tyler et al., 2015) i a Finlàndia (https://laji.fi/en/ taxon/MX.41583).
Pyracantha crenulata (D. Don) M. Roemer
OsOna: Espinelves, riba dreta de la riera d’Espinelves, vora el pont de Masjoan, DG5135, 695 m, plantació de coníferes antiga, 2-II-2021 (P. Aymerich, herb. pers.).
Confirmació de la presència a Catalunya, com a aŀlòcton, d’aquest arbust ornamental originari de l’Himàlaia. Tot i haver estat citada ja per Casasayas (1989) d’una localitat d’Osona i una de la Selva, i més endavant d’altres llocs, P. crenulata no va ser inclosa a la checklist de la flora aŀlòctona de Catalunya (Aymerich & Sáez, 2019) perquè es va constatar que habitualment aquest nom s’havia aplicat de forma errònia a P. fortuneana (Maxim.) H.L. Li, una espècie xinesa similar i que és freqüent com a naturalitzada en zones suburbanes. P. crenulata es distingeix de P. fortuneana sobretot pel contorn oblong o oblanceolat de les fulles, que són obovades en la segona espècie (Cuizhi & Spongberg, 2003). A Espinelves hi vam observar un exemplar solitari, ben desenvolupat.
Quercus rubra L.
MOianès: Sant Quirze Safaja, pla del Badó, entre les cases de Poses i del Badó, parceŀla arbrada i sense edificar en una àrea
158 Butlletí ICHN 85 (3), 2021
P AyMERICH & L SáEz

a b c d



semiurbanitzada, DG2919, 710 m, 2 individus joves d’1-3 m d’alçada, 28-X-2020 (P. Aymerich).
Roure originari de l’est d’Amèrica del nord, citat prèviament de cinc localitats catalanes (cf. Verloove & Aymerich, 2020), sempre amb caràcter d’espècie aŀlòctona casual. Els dos individus observats a Sant Quirze provenien de la dispersió a pocs metres de llavors de quatre individus plantats.
Robinia viscosa Vent.
Berguedà: Berga, polígon industrial de la Valldan, DG0261, 770-790 m, vores de vials i erms entre parceŀles edificades,
uns 20 individus joves a distàncies inferiors a 50 m’arbres plantats, 15-VIII-2020 (P. Aymerich).
Novetat per a la flora aŀlòctona catalana. Aquest arbre és originari de l’est d’Amèrica del nord, amb una àrea natural limitada a un sector de la serralada dels Apalatxes, i es separa de R. pseudoacacia L. per les flors rosades i les branques i llegums amb glàndules (Peabody, 1984). Es fa servir en jardineria i està introduïda en altres llocs d’Amèrica (Nova Anglaterra) i també a Europa (Peabody, 1984). A l’Europa meridional aquesta espècie gairebé no ha estat citada, amb l’excepció d’Itàlia, on es considera naturalitzada i invasora
159 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
APORTACIONS A LA FLORA AŀLÒCTONA CATALANA
Figura 3. a) Pistacia vera; b) Puschkinia scilloides; c) Salvia lavandulifolia subsp. gallica; d) Viburnum rhytidophyllum. Fotografies: a, b, d: Pere Aymerich; c: Llorenç Sáez.
GEA, FLORA ET FAUNA
local a la Llombardia (Banfi & Galasso, 2010). Els escassos individus observats a Berga corresponen a una presència casual en hàbitats netament antròpics, però no es pot excloure una expansió futura, com la que ha experimentat a la mateixa localitat R. pseudoacacia, molt més abundant i ja present en hàbitats seminaturals.
Salvia lavandulifolia subsp. gallica W. Lippert [S. officinalis subsp. gallica (W. Lippert) Reales & al.]
Vallès Oriental: Sant Feliu de Codines, vessant sud del pla de la Costa, DG3016, 545 m, brolles en antigues marjades, en terreny calcari i vora la xarxa viària, 19-V-2014 (L. Sáez, herb. pers.). Figura 3c.
Tàxon que ja s’havia indicat prèviament de Catalunya, però del qual se’n tenia informació confusa tant en relació amb el seu caràcter aŀlòcton com amb el tractament taxonòmic. Les plantes del grup de Salvia. officinalis – S. lavandulifolia que tenen calze netament bilabiat i amb una nervadura molt visible entre els nervis principals del calze corresponen a S. officinalis L. en sentit estricte (Sáez, 2010; Reales et al., 2004), mentre que les que presenten el calze feblement bilabiat (i ocupen una posició intermèdia entre l’espècie anterior i la típica S. lavandulifolia Vahl) correspondrien al tàxon que ens ocupa, que també s’ha tractat com a subspècie de S. officinalis (Reales et al., 2004). A la revisió del gènere per a Flora iberica (Sáez, 2010) es va incloure S. lavandulifolia subsp. gallica dins d’un concepte ampli de S. officinalis, no només per l’estreta relació taxonòmica entre aquests tàxons, sinó també per la poca disponibilitat de dades relatives a quines poblacions es trobaven naturalitzades al territori ibèric i balear. Salvia officinalis és originària de les penínsules balcànica i itàlica, i està introduïda a la península Ibèrica, trobantse ocasionalment com a persistent de cultiu o casual, i de vegades localment naturalitzada. Aquesta mateixa situació es donaria per a S. lavandulifolia subsp. gallica, les poblacions naturals de la qual es troben al sud de França. Les localitats ibèriques aportades per Reales et al. (2004), basades en testimonis d’herbari, no corresponen realment a poblacions naturals (aspecte no explicitat pels autors anteriors), sinó que serien exemplars escapats de zones situades prop de nuclis urbans, en llocs més o menys antropitzats o pertorbats. En el cas de Catalunya havia estat indicada de Canet de Mar, Premià de Mar, Arbúcies i Manlleu, on no hi ha poblacions naturals de plantes del grup de S. officinalis - lavandulifolia, com així ja ho indiquen Bolòs & Vigo (1996) i més concretament Montserrat (1968) per a les poblacions del Maresme. A la nova localitat de Sant Feliu de Codines es van observar tres exemplars reproductors i des del 2014 no s’ha observat reclutament; de fet dos exemplars han desaparegut en temps recents, motiu pel qual sembla molt improbable un establiment local.
Symphoricarpos albus (L.) S.F. Blake
Berguedà: Avià, Graugés, al costat de cal Torrentbò, DG0457, 600 m, marges de camp i bardisses, 4-X-2019 (P. Aymerich); MOianès: Sant Quirze Safaja, entre el nucli urbà i el torrent de la font del Boix, al sector parcialment edificat del carrer
d’en Guitert, DG2919, 615 m, bosc de roures sota una casa amb jardí, 4 individus espontanis, 23-X-2020 (P. Aymerich).
Arbust d’origen nord-americà que es cultiva com a ornamental i sovint s’escapa a Europa, però que ha estat molt poc citat a Catalunya. Les noves dades corresponen a una dispersió a distància curta al Moianès –a menys de 50 m d’exemplars cultivats– i a una naturalització vegetativa in situ al Berguedà, similar a la del Solsonès.
Thuja plicata Donn
OsOna: Espinelves, riba dreta de la riera d’Espinelves, vora el pont de Masjoan, DG5135, 695 m, plantació de coníferes antiga i talús sobre la riera, 2 individus espontanis petits, 2-II-2021 (P. Aymerich).
Conífera originària de la costa oest d’Amèrica del Nord, que es cultiva a Europa com a arbre forestal i ornamental. És freqüent com a escapada de cultiu a l’oest i nord del continent, però no havia estat citada de Catalunya. A Espinelves es van observar dos individus joves (1 i 0,3 m d’alçada) separats una distància d’uns 20 m.
Trachycarpus fortunei (Hook.) H. Wendl.
Vallès Occidental: Sant Cugat de Vallès, serra de Collserola, capçalera del torrent de la Salamandra, DF2686, 330-370 m, fondal del torrent, 22-X-2020 (L. Sáez).
Tot i que la presència d’aquesta palmera d’origen xinès és més o menys coneguda des de fa anys a Collserola, no sabem que n’hi hagi cap citació publicada formalment. A la localitat indicada l’espècie sembla plenament naturalitzada i la seva presència ha de remuntar-se a dècades enrere, ja que no rarament es troben exemplars de fins a prop de 10 metres d’alçada. En els darrers temps, T. fortunei s’ha anat citant cada vegada més a Catalunya, i en diverses localitats mostra indicis de naturalització; per a una síntesi de dades consulteu Verloove & Aymerich (2020).
Trifolium suaveolens Willd. [T. resupinatum L. var. majus Boiss.]
Berguedà: l’Espunyola, voltants de la Casanova, CG9756, 830-840 m, herbassars al costat de camins rurals i també com a mala herba en un camp, uns quants centenars d’individus, 6-VI-2021 (P. Aymerich, herb. pers.).
Tàxon originari de l’Orient Pròxim, que a Catalunya només havia estat citat d’una localitat de la Selva (Mercadal, 2019), aparentment en unes condicions similars a les de la nova dada que aportem, A l’Espunyola era cultivat en un camp i es va observar com a escapat en herbassars de quatre llocs diferents, amb una distància d’uns 600 m entre els punts extrems.
Viburnum rhytidophyllum Hemsl.
ripOllès: Camprodon, perifèria oest del nucli urbà, al vessant sobre l’avinguda de Maristany, DG4785, 965 m, arbreda seminatural (barreja de freixeneda, roureda i plantació de coníferes) prop de cases, individu solitari, 2-XI-2020 (P. Aymerich). Figura 3d.
160 Butlletí ICHN 85 (3), 2021
P AyMERICH & L SáEz
Novetat per a la flora aŀlòctona catalana. Aquesta espècie arbustiva, originària del sud de la Xina, es cultiva com a ornamental i s’està naturalitzant en diverses regions de l’Europa de clima temperat (Walther, 2000; Fuchs et al., 2006; Stace, 2010; Galasso et al., 2018; Verloove,2020). Recentment (Aymerich, 2020a) s’ha indicat d’una localitat pròxima, Sant Joan de les Abadesses, Viburnum ×rhytidophylloides J.V. Suringar, híbrid de jardineria entre V. rhytidophyllum i V. lantana L. La distinció entre V. rhytidophyllum i V. ×rhytidophylloides pot ser complexa, sobretot perquè el segon és un tàxon variable que es pot acostar més o menys a les dues espècies parentals. La planta de Camprodon ha estat atribuïda a V. rhytidophyllum pels caràcters que solen considerar-se distintius, com l’absència de pèls estelats a l’anvers foliar, les fulles amb relació longitud-amplada superior a 3 i el marge foliar enter (Stace, 2010; Langhe, 2012).
Agraïments
Volem agrair a L. Gustamante la recoŀlecció de material de Linaria versicolor i a Moisès Guardiola l’ajut en la prospecció de camp que va permetre detectar Myoporum insulare
Referències
Aletà, J., Ninot, A. & Voltas, J. 2003. Caracterización del comportamiento agroforestal de doce genotipos de nogal (Juglans sp.) en dos localidades de Cataluña. Investigación agraria: Sistemas y recursos forestales, 12: 39-50.
Aymerich, P. 2013. Plantas alóctonas de origen ornamental en la cuenca alta del río Llobregat (Cataluña, noreste de la Península Ibérica). Bouteloua, 16: 52-79.
Aymerich, P. 2014. Notes florístiques de les conques altes dels rius Segre i Llobregat (II). Orsis, 28: 7-47.
Aymerich, P. 2015. Notes florístiques de les conques altes dels rius Segre i Llobregat (III). Orsis, 29: 1-28.
Aymerich, P. 2016. Contribució al coneixement de la flora aŀlòctona del nord i centre de Catalunya. Orsis 30: 11–40.
Aymerich, P. 2017. Notes sobre flora aŀlòctona a Catalunya. Butlletí de la Institució Catalana d’Història Natural, 81: 97-116.
Aymerich, P. 2019. Notes sobre flora aŀlòctona a Catalunya. II. Butlletí de la Institució Catalana d’Història Natural, 83: 3-21.
Aymerich, P. 2020a. Notes sobre flora aŀlòctona a Catalunya. III. Butlletí de la Institució Catalana d’Història Natural, 84: 101124.
Aymerich, P. 2020b. Physalis angulata (Solanaceae), naturalizada en la Península Ibérica. Acta Botanica Malacitana, 45: 221-224.
Aymerich, P. & Sáez, L. 2019. Checklist of the vascular alien flora of Catalonia (northeastern Iberian Peninsula, Spain). Mediterranean Botany, 40: 215-242.
Bacchetta, G., Calvia, G. & Ruggero, A. 2019. Notulae to Italian vascular flora, 7: Abies cephalonica Loudon. Italian Botanist, 7: 159.
Banfi, E. & Galasso, G. 2010. La flora esotica lombarda. Regione Lombarda & Museo Storia Naturale di Milano. 273 p.
Bolòs, O. & Vigo, J. 1996. Flora dels Països Catalans Vol. 3. Ed. Barcino. Barcelona. 1230 p.
Casasayas, T. 1989. La flora aŀlòctona de Catalunya. Tesi doctoral. Facultat de Biologia, Universitat de Barcelona. 880 p. https:// www.tesisenred.net/handle/10803/969 [Accés: 1 agost 2021]
Cuizhi, G. & Spongberg, S.A. 2003. Pyracantha M. Roemer. P. 108111. In: zheng-yi, W.; Raven, P. H.; Hong, D. Flora of China. Vol. 9. Missouri Botanical Garden Press. 496 p.
Dickoré, W. & Kasperek, G. 2010. Species of Cotoneaster (Rosaceae, Maloideae) indigenous to, naturalising or commonly cultivated in Central Europe. Willdenowia 40: 13-45.
Dickson, E. E. 2013. Malus L. In: Flora of North America Editorial Comitee (Eds.) Flora of North America north of Mexico. Vol. 9. P. 472-480. Oxford University Press. 600 p.
Férnandez-Moya, J., Lieca, R. J., Urbán, I., Catro, R. & Ramírez, C. 2020. Clonal effect of rooting and acclimation rates for in-vitro micropropagation in hybrid walnut (Juglans ×intermedia Mj 209): preliminary observations. Annals of silvicultural research, 44: 41-46.
Frey, D. & Moretri, M. 2019. A comprehensive dataset on cultivated and spontaneously growing vascular plants in urban gardens. Data in Brief, 25: 103982.
Fuchs, R., Kutzelnigg, H. & Feige, G. B. 2006. Seminatural ancient forests in urban agglomeration area Ruhrgebeit. Acta Biologica Benrodis, 13: 91-104.
Fryer, J. & Hylmö, B. 2009. Cotoneasters A comprehensive guide to shrubs for flowers, fruit, and foliage. Timber Press. Portland. 344 p.
Galasso, G., Conti, F., Peruzzi, L., Ardenghi, N. M. G, Banfi, E., Celesti-Grapow, L., Albano, A., Alessandrini, A., Bacchetta, G., Ballelli, S., Bandini Mazzanti, M., Barberis, G., Bernardo, L., Blasi, C., Bouvet, D., Bovio, M., Cecchi, L., Guacchio, E., Domina, G., Fascetti, S., Gallo, L. Gubellini, L., Guiggi, A., Iamonico, D., Iberite, M., Jiménez-Mejías, P., Lattanzi, E., Marchetti, D., Martinetto, E., Masin, R. R., Medagli, P., Passalacqua, N. G., Peccenini, S., Pennesi, R., Pierini, B., Podda, L. , Poldini, L., Prosser, F., Raimondo, F. M., Roma-Marzio, F., Rosati, L., Santangelo, A. Scoppola, A., Scortegagna, S., Selvaggi, A., Selvi, F., Soldano, A., Stinca, A., Wagensommer, R. P., Wilhalm, T. & Bartolucci, F. 2018. An updated checklist of the vascular flora alien to Italy. Plant Biosystems, 152: 179-303.
Gonin, P. (coord.) 2013. Autoecología de las frondosas nobles Institut pour le Développement Forestier. París. 64 p.
Guardiola, M. & A. Petit 2020. Aportacions a la flora aŀlòctona de la serralada Litoral central catalana i territoris propers. Butlletí de la Institució Catalana d’Història Natural, 84: 35-49.
International Geraniaceae Group. 2021. The Register of Erodium Cultivars Names. https://www.geraniaceae-group.org [Accés: 30 juliol 2021]
Kaltsidia, M. P., Fernández, R., Franco, A. & Pérez, S.M.P. 2020. Improving the performance of felt- based living wall systems in terms of irrigation management. Urban Forestry & Urban Greening, 54: 126782
Keil, P. & Loos, G. H. 2005. Non-established adventive plants in the western and central Ruhrgebeit (Nortrhine-Westphalia, Germany) – a preliminary overview. Electronic Publications of the Biological Station of Western Ruhrgebiet, 5: 1-16.
Langhe, J. 2012. Viburnum L. Adoxaceae. Vegetative key to species in cultivation. Ghent University Botanical Garden. 15 p.
Lingdi L. & Brach, A. 2003. Cotoneaster L.. In: Flora of China Editorial Comitee (Eds.) Flora of China.Vol. 9. P. 95-108. Science Press & Missouri Botanical Garden Press, Beijing & St. Louis.
Mercadal, G. 2019. Els prats de dall de la terra baixa catalana. Caracterització geobotànica, valoració agroambiental i estudi de les relacions fitosociològiques entre els prats dalladors de l’Europa occidental. Universitat de Girona. 2306 p. https://www. tdx.cat/handle/10803/669044#page=1
Mercadé, A. 2016. Estudis de flora i vegetació del Moianès i àrees properes. Tesi doctoral. Universitat de Barcelona. 619 p. https:// www.tesisenred.net/handle/10803/403059 [Accés 1 agost 2021]
161 Butlletí ICHN 85 (3), 2021 GEA, FLORA ET FAUNA
LA FLORA AŀLÒCTONA CATALANA
APORTACIONS A
GEA, FLORA ET FAUNA
Montserrat, P. 1968. Flora de la cordillera litoral catalan (porción comprendida entre los ríos Besós y Tordera). Caja de Ahorros de Mataró. 453 p.
Nielsen, T. F., Sand-Jensen, K., Dornalas, M. & Bruun, H. H. 2019. More is less: Net gain in species richness, but biotic homogeneization over 140 years. Ecology Letters, 22: 16501657.
Peabody, F. J. 1984. Revision of the genus Robinia (Leguminosae, Papilionidae). Retrospective Thesis and Dissertations. 7787. Iowa State University. 155 p.
Pyšek, P., Danihelka, J., Sádlo, J., Chrtek, J., Chytrý, M., Jarošík, V., Kaplan, z., Krahulec, F., Moravcová, L., Pergl, J., Štajerová, K. & Tichý, L. 2012. Catalogue of alien plants of the Czech Republic (2nd edition): checklist update, taxonomic diversity and invasion patterns. Preslia, 84: 155-255.
Reales, A., Rivera, D., Palazón, J. A. & Obón, C. 2004. Numerical taxonomy study of Salvia sect. Salvia (Labiatae). Botanical Journal of the Linnean Society, 145: 353-371.
Sáez, L. 2019. Linaria Mill. In Flora of North America North of Mexico. vol 17 Magnoliophyta: Tetrachondraceae to Orbobanchaceae: 27-33. New York and Oxford. Oxford University Press.
Sáez, L. 2010. Salvia L. P. 298-236. In: R. Morales, A. Quintanar, F. Cabezas, A. J. Pujadas & Cirujano, S. (eds.), Flora iberica. Vol.
XII. Verbenaceae-Labiatae-Callitrichaceae. Real Jardín Botánico de Madrid-CSIC. Madrid. 650 p.
Sáez, L., Pié, G. & Carnicero, P. 2017. Catàleg de la flora vascular del massís del Montseny. Tres segles d’investigació botànica (1716-2016). Coŀlecció Estudis. Sèrie Territori i Parcs Naturals 1. Diputació de Barcelona. 238 p.
Senar, R. & Cardero, S. 2019. Dades de plantes aŀlòctones per a l’est de la península Ibèrica. Collectanea Botanica (Barcelona), 38: e009.
Stace, C. 2010. New flora of the British Isles, 3th ed. Cambridge University Press. XXXII + 1232 p.
Strother, J. L.. 2006. Gaillardia Foug. In: Flora of North America north of Mexico. Vol. 21. P. 415-421. Oxford University Press. 616 p.
Tyler, T., Karlsson, T., Milberg, P., Sahlin, U. & Sundberg, S. 2015. Invasive plant species in the Swedish flora: developing criteria and definitions, and assessing the invasiveness of individual taxa. Nordic Journal of Botany, 33: 300-317.
Verloove, F. 2020. Manual of the alien plants of Belgium. Disponible a: http://alienplantsbelgium.be/ [Accés: gener a juny 2021].
Verloove, F. & Aymerich, P. 2020. Chorological novelties for the alien flora of the province of northeastern Catalonia (Iberian Peninsula). Butlletí de la Institució Catalana d’Història Natural, 84: 137-153.
Walther, G.R. 2000. Laurophyllisation in Switzerland. Swiss Federal Institute of Technology zurich. Tesi doctoral. 150 p.
162 Butlletí ICHN 85 (3), 2021
P AyMERICH & L SáEz
Aportacions al coneixement de la flora de les Guilleries orientals i àrees properes (nord-est de Catalunya) – III
Josep Gesti Perich* & Lluís Vilar Sais**
* Línia de recerca de Flora i Vegetació. Universitat de Girona. Campus Montilivi. 17003 Girona. A/e: josepgesti@gmail.com
** LAGP-Flora i Vegetació. Institut de Medi Ambient. Universitat de Girona. Campus Montilivi. 17003 Girona. A/e: lluis.vilar@udg.edu
Autor per a la correspondència: Josep Gesti. A/e: josepgesti@gmail.com
Rebut: 16.09.2021; Acceptat: 23.09.2021; Publicat: 30.12.2021
Resum
Aportem dades sobre diversos tàxons aŀlòctons observats a l’àrea oriental de les Guilleries, on no hi havien estat indicats prèviament i que són rars o poc coneguts al nod-est de Catalunya. Indiquem Acalypha rhomboidea i Pleioblastus simonii per primera vegada a Catalunya i donem noves localitats d’Aspidistra elatior, Cardamine occulta, Celosia cristata, Guizotia abyssinica, Hemerocallis fulva, Koelreuteria paniculata, Lupinus albus, Persicaria orientalis, Sedum palmeri, Silene pseudoatocion i Soleirolia soleirolii.
Paraules clau: flora aŀlòctona, Guilleries, Catalunya, península Ibèrica.
Abstract
Contributions to the knowledge of the flora of the Eastern Guilleries and nearby areas (northeastern Catalonia) – III
We provide data on alien flora observed in the eastern area of the Guilleries massif, not previously reported in this area and that are rare or little known in Catalonia. We indicate Acalypha rhomboidea and Pleioblastus simonii for the first time in Catalonia and we give new localities of Aspidistra elatior, Cardamine occulta, Celosia cristata, Guizotia abyssinica, Hemerocallis fulva, Koelreuteria paniculata, Lupinus albus, Persicaria orientalis, Sedum palmeri, Silene pseudoatocion and Soleirolia soleirolii.
Key words: alien flora, Guilleries, Catalonia, Iberian Peninsula. Butlletí
Introducció
En aquest article aportem informació sobre la presència de diverses plantes al sector oriental del massís de les Guilleries (nord-est de Catalunya). Seguint la línia de treballs anteriors (Gesti & Vilar, 2019, 2020), es tracta d’espècies que no hi havien estat indicades i que, a més, són infreqüents o poc conegudes a Catalunya. En aquesta ocasió totes les espècies tractades són aŀlòctones pel territori.
Material i mètodes
Les dades que aportem provenen dels municipis de Riudarenes i Santa Coloma de Farners, a l’extrem oriental del massís de les Guilleries, i han estat obtingudes al llarg de l’any 2021. El llistat dels tàxons es presenta en ordre alfabètic i, per cadascun, donem localitats concretes amb indicació de la comarca, municipi, lloc, quadrat UTM d’1 km de costat (sempre a la zona 31T, sistema de referència ETRS89), altitud, hàbitat, observador, data d’observació i –quan n’hi ha- el número de plec de l’Herbari de la Universitat de Girona (HGI). A cada tàxon fem una breu descripció de l’àrea de
procedència, les citacions anteriors al nord-est de Catalunya, l’hàbitat, l’estat de les poblacions localitzades i el grau de naturalització a nivell local.
Resultats i discussió
Acalypha rhomboidea Raf. [A. virginica L. var. rhomboidea (Raf.) Cooperr.]
Selva: Santa Coloma de Farners, a tocar del parc de Sant Salvador, DG7133, 120 m, herbassar ruderal (HGI 24251, J. Gesti, 13-IX-2021).
Euforbiàcia originària de l’Amèrica del Nord que s’ha introduït en algunes regions d’Europa (nord d’Itàlia, Àustria, Bèlgica i Portugal) (Domingues & da Costa, 2006; GBIF, 2021), on ocupa ambients antropitzats (a Portugal, per exemple, s’ha constatat que es va estendre per diferents jardins i espais viaris de Coimbra a partir de plantes cultivades al jardí botànic de la ciutat; Domingues & Da Costa, 2006). No hi ha un consens total en relació al seu rang taxonòmic, ja que per alguns autors es tracta d’una espècie independent (Levin, 1999, 2021; POWO, 2021), mentre que d’altres la tracten com una varietat d’A. virginica L. (var. rhomboidea (Raf.)
163 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA GEA, FLORA ET FAUNA
de la Institució Catalana d’Història Natural, 85 (4): 163-169. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition) DOI: 10.2436/20.1502.01.89

Cooperr.) (Cooperrider, 1984; GBIF, 2021). Els principals caràcters diferencials respecte d’A. virginica són les bràctees de les flors pistilades, que tenen un nombre reduït de lòbuls i presenten poca pilositat eglandular a la cara abaxial (els pèls glandulars són molt més aparents). A Europa A. virginica (en sentit ampli) ha estat indicada en diverses regions (Tutin et al., 1968; GBIF, 2021, Verloove, 2021), sobretot al nord d’Itàlia i territoris veïns, de manera que no es pot descartar que alguna de les cites correspongui al tàxon al que aquí ens referim.
N’hem observat una població formada per alguns centenars d’individus, en flor al mes de setembre, ocupant una superfície d’uns 40 m2 en una zona ruderalitzada situada entre un parc i una plantació de plàtans (Fig. 1). Tractant-se d’una planta anual, la presència d’un nombre tan gran d’efectius fa pensar que ja hi havia de ser en anys precedents i que està naturalitzada localment.
Aspidistra elatior Blume
Selva: Santa Coloma de Farners, al Veïnat d’Avall, entre els horts i la riera, DG7233, 120 m, talús d’un rec ombrívol, J. Gesti & L. Vilar, 11-II-2021; Santa Coloma de Farners, a la riera dels Frares, DG7134, 130 m, talús sobre la riera, J. Gesti & L. Vilar, 11-II-2021.
Asparagàcia originària de l’Àsia de l’Est que és emprada en jardineria com a planta ornamental. A Catalunya ha estat indicada fora de cultiu al Baix Llobregat (Gómez-Bellver et al., 2019).
A les localitats reportades hi hem observat només alguns individus aïllats de grans dimensions (Fig. 2), que creixien

164 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA J. GESTI PERICH & L. VILAR SAIS
Figura 2. Aspidistra elatior, Santa Coloma de Farners.
a b c d
Figura 1. Acalypha rhomboidea, Santa Coloma de Farners: a) hàbitat; b) port general; c) detall de les fulles amb les inflorescències disposades a les axiŀles; d) detall de la inflorescència.
en talussos humits i ombrívols. En un dels indrets (riera dels Frares) hi hem constatat freqüents abocaments de restes vegetals, que podrien constituir el mecanisme d’arribada de la planta a aquests espais. Tot i que s’utilitza sovint com a planta d’interior, tolera bé el fred i un cert grau de sequera, fet que li permetria subsistir en aquests ambients durant llargs períodes de temps. Al territori estudiat és una aŀlòctona casual.
Cardamine occulta Hornem.
Selva: Riudarenes, a la riera de Santa Coloma, prop de can Sala, DG7628, 80 m, sorral fluvial, J. Gesti, 14-III-2021; Santa Coloma de Farners, a la riera de Santa Coloma, aigües avall del pont del balneari, DG7232, 100 m, codolar fluvial (HGI 24252, J. Gesti, 1-III-2021); Santa Coloma de Farners, a la riera de Santa Coloma, davant la sorrera, DG7233, 115 m, sorral fluvial, J. Gesti, 4-III-2021.
Brassicàcia d’origen asiàtic que sembla trobar-se en expansió a Europa, tot i que la seva distribució és encara mal coneguda per la possible confusió amb C. hirsuta L. o C. flexuosa With. i per la controvèrsia existent durant anys sobre la seva assignació taxonòmica (es pot consultar una síntesi sobre aquests aspectes a Aymerich, 2016a). A Catalunya es coneix de l’Alt Urgell, el Pallars Jussà, el Solsonès, la Garrotxa i Barcelona (Aymerich, 2016a, 2017).
L’hem observada en diferents punts al llarg de la riera de Santa Coloma, sobre substrats sorrencs i pedregosos de la llera (Fig. 3), on s’ha naturalitzat i manté poblacions formades per centenars d’individus.

Celosia cristata L. [C. argentea L. var. cristata (L.) Kuntze]
Selva: Riudarenes, a la riera de Santa Coloma, DG7629, 85 m, sorral fluvial, J. Gesti & L. Vilar, 18-VIII-2021.
Amarantàcia nativa de les zones tropicals del vell món introduïda a les àrees tropicals i subtropicals –i algunes temperades– del planeta (POWO, 2021). El rang taxonòmic d’aquesta Celosia d’inflorescències crestades i flors de color porpre (coneguda popularment com «cresta de gall») és controvertit, ja que alguns autors la consideren una varietat (var. cristata (L.) Kuntze), una forma (f. cristata (L.) Schinz) o
un sinònim de C. argentea L., mentre que d’altres la separen a nivell específic (es pot consultar una síntesi sobre la seva circumscripció taxonòmica a la revisió feta per Iamonico, 2013). A Catalunya ha estat indicada a l’Empordà (Vayreda, 1902: «Lladó, Sagaró, etc., semiespontánea») i al Centre de Documentació de Biodiversitat Vegetal de la Universitat de Barcelona hi ha un plec d’herbari (BCN 91866, sub Celosia cristata L. var. fasciata, 9-XI-1934, recoŀlector desconegut) amb l’anotació «Barcelona, carrer de Mariana Pineda» que podria correspondre, per tant, a un exemplar cultivat. Per altra banda, recentment també s’ha trobat a Catalunya C. argentea (en sentit estricte) al Maresme, al riu Tordera (Verloove & Aymerich, 2020).
N’hem observat un únic individu a la llera seca de la riera de Santa Coloma, sobre substrat sorrenc, florit al mes d’agost (Fig. 4). Al territori estudiat és una aŀlòctona casual.

Guizotia abyssinica (L. f.) Cass.
Selva: Santa Coloma de Farners, a la riera de Santa Coloma, DG7233, 120 m, herbassar fluvial humit (HGI 24241, J. Gesti, 30-VIII-2021).

165 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA APORTACIONS AL CONEIXEMENT DE LA FLORA DE LES GUILLERIES ORIENTALS – III
Figura 3. Cardamine occulta, Santa Coloma de Farners.
Figura 4. Celosia cristata, Riudarenes.
Figura 5. Guizotia abyssinica, Santa Coloma de Farners.
GEA, FLORA ET FAUNA
Aquesta asteràcia originària de l’Àfrica oriental va ser citada per primera vegada fora de cultiu a la península Ibèrica a principis de la dècada dels anys 70 del segle passat (Cid, 1971) i poc després també es trovaba a Catalunya (Folch & Abella, 1974). Des d’aleshores ha estat indicada en nombroses localitats del litoral i prelitoral de la meitat meridional de Catalunya (Font, 2021) però, en canvi, ho ha estat molt escassament a la meitat nord: la Selva (Casasayas, 1989), Baix Empordà (HGI 17399, Santa Cristina d’Aro, A. Mallol & J. Maynés, 7-I-2003; Mallol & Maynés, 2008), Alt Urgell i Berguedà (Aymerich, 2016a).
N’hem observat dos exemplars, en flor a final d’agost, en un herbassar sobre sòl sorrenc humit a la llera de la riera (Fig. 5). És casual al territori.
Hemerocallis fulva (L.) L.
Selva: Santa Coloma de Farners, a la riera de Santa Coloma, sota el Bagís, DG6634, 260 m, verneda, J. Gesti, 6-VII2021.
Indiquem la presència d’aquesta asfodelàcia nativa de l’Àsia oriental al sector oriental del massís de les Guilleries, d’on no se’n tenia constància, per bé que ja era coneguda d’àrees properes com ara el Montseny (Bolòs et al., 1986; Casasayas, 1989; Sáez et al. 2015, 2017), les Guilleries occidentals (Pérez-Haase et al., 2013), el curs mitjà del riu Ter (Girbal, 1984), l’estany de Sils (Vilar, 1987), etc.
N’hem observat dos rodals propers entre ells, ocupant densament una superfície total d’uns 5 m2, en una verneda ben constituïda del tram alt de la riera de Santa Coloma. No sembla una espècie plenament naturalitzada al territori, sinó casual.
Koelreuteria paniculata Laxm.
Selva: Santa Coloma de Farners, entre la sorrera i el balneari, DG7232, 120 m, talús d’una petita riera, J. Gesti & L. Vilar, 2-IX-2021.
Aquesta sapindància originària de l’Àsia oriental és utilitzada com a arbre ornamental en parcs i jardins. Al nord-est de Catalunya (com a la resta del territori català) hi ha relativament poques cites fora de cultiu (vegeu localitats a Font, 2021), cap d’elles gaire propera a la nostra zona d’estudi: Berguedà, Garrotxa, Vallès Oriental (Casasayas, 1989), Maresme (Guardiola & Petit, 2020), Bages (Aymerich, 2020), etc.
N’hem observat un únic individu, de port arbustiu (1,5 m aproximadament), que creixia en un talús sobre d’un petit curs d’aigua intermitent. Al territori estudiat és una aŀlòctona casual.
Lupinus albus L.
Selva: Riudarenes, prop del molí d’en Pastrells, DG7431, 100 m, marge de camp, J. Gesti, 27-III-2021.
Es tracta d’una fabàcia, probablement originària de la Mediterrània oriental, que s’ha estès a causa del seu cultiu per al consum de les llavors i com a farratgera. A Catalunya se’n coneixen diverses indicacions, sobretot al sector nord-orien-
tal, per bé que la majoria d’elles corresponen a observacions de final del segle XIX i de la primera meitat del segle XX (Font, 2021). Les més properes a la nostra zona d’estudi són les cites fetes al Montseny per Joan Cadevall (1915-1919) i per Pius Font i Quer (Casasayas, 1989).
A la localitat indicada n’hi hem observat algunes desenes d’individus, en flor al mes de març, que ocupaven el marge d’un camp plantat de faves (Fig.6). Desconeixem amb certesa el seu origen, per bé que és possible que s’hi hagin instaŀlat a partir de llavors procedents de cultius anteriors en aquell mateix indret o bé als camps del seu entorn.

Persicaria orientalis (L.) Spach [Polygonum orientale L.]
Selva: Santa Coloma de Farners, a la riera de Santa Coloma, aigües amunt del pont del balneari, DG7233, 115 m, herbasar fluvial humit (HGI 24242, J. Gesti, 30-VIII-2021).
Aquesta amarantàcia originària del sud-est asiàtic, que es cultiva com a planta ornamental, ja era coneguda en àrees properes a la nostra zona d’estudi: al Montseny (Romo, 1987; Pié et al., 2010), a les Guilleries occidentals (Batriu et al., 2012; Pérez-Haase et al., 2013), al curs mitjà del riu Ter

166 Butlletí ICHN 85 (4), 2021
Figura 6. Lupinus albus, Riudarenes.
J. GESTI PERICH & L. VILAR SAIS
Figura 7. Persicaria orientalis, Santa Coloma de Farners.

(Girbal, 1984), a la plana de la Selva (HGI 9416, Maçanet de la Selva, L. Vilar, 15-VIII-1981), etc. Constatem també, doncs, la seva presència a les Guilleries orientals.
N’hem observat un únic exemplar, en flor a final d’agost, en un herbassar sobre sòl sorrenc humit a la llera de la riera (Fig. 7). És casual al territori estudiat.
Pleioblastus simonii (Carrière) Nakai [Arundinaria simonii (Carrière) Rivière & C. Rivière]
Selva: Santa Coloma de Farners, a la riera dels Frares, a l’alçada de l’antic escorxador, DG7134, 130 m, talús de la riera, J. Gesti, 18-II-2021; Santa Coloma de Farners, a la riera dels Frares, sota Can Malladó, DG7135, 140 m, talussos humits (HGI 24254, J. Gesti, 2-III-2021).
Bambú originari del Japó que s’utilitza en jardineria com a planta ornamental. Per bé que la determinació d’espècies d’aquest grup de poàcies és molt complexa –més encara tenint en compte l’existència de varietats de cultiu–, atribuïm el tàxon observat a P. simonii en base al conjunt de caràcters morfològics diferencials descrits a la bibliografia (Ohwi, 1965; Suzuki, 1978; Kobayashi & Furumoto, 2004). Les dimensions de les tiges (les més grans superant els 3 m d’alçada i els 2,5 cm de diàmetre), les nombroses ramificacions
als nusos, el marge superior de les beines foliars oblic i el revers foliar amb tendència a ser bicolor, el diferenciarien d’altres espècies semblants d’aquest gènere, com ara P. chino (Franch. & Sav.) Makino. A Europa ha estat indicat a les Illes Britàniques (Stace, 2019) i a GBIF (2021) s’hi recullen plecs procedents de Dinamarca i Galicia que, per les dades que hi consten, potser provinguin de plantes cultivades.
N’hem observat diversos nuclis al llarg d’un tram d’uns 900 m a la riera dels Frares, el més gran dels quals s’estén de manera discontínua sobre una superfície d’uns 200 m2, mentre que la resta són rodals d’uns pocs metres quadrats cadascun. Durant els mesos de febrer i març de 2021 el nucli principal ha presentat una profusa emergència de noves tiges que han florit abundantment (Fig. 8). Naturalitzat a nivell local.
Sedum palmeri S. Watson
Selva: Santa Coloma de Farners, al nucli urbà, DG7234, 140 m, teulada, J. Gesti & L. Vilar, 13-II-2021.
Crassulàcia originària de Mèxic que és freqüentment plantada com a espècie ornamental, sobretot en testos i rocalles. A Catalunya ha estat indicada fora de cultiu en unes poques localitats, al Berguedà, al Vallès Oriental (Aymerich & Sáez, 2015) i al Tarragonès (Verloove et al. 2019). Coincidim amb
167 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA
GUILLERIES ORIENTALS – III a b c d
Figura 8. Pleioblastus simonii, Santa Coloma de Farners: a) hàbitat; b) port general i aspecte de les tiges; c) detall de les fulles i les beines foliars; d) detall de la inflorescència.
APORTACIONS AL CONEIXEMENT DE LA FLORA DE LES
GEA, FLORA ET FAUNA
els autors d’aquestes citacions que és molt probable que es faci en d’altres indrets atesa la seva àmplia utilització en jardineria i la facilitat amb la que es propaga.
L’hem observada en teulades del poble, ocupant les canaleres formades per les teules i les canals de recollida d’aigua, on s’acumulen petits dipòsits de sediments (Fig. 9). Naturalitzada a nivell local.

Silene pseudoatocion Desf.
Selva: Santa Coloma de Farners, a la riera de Santa Coloma, sota el pont del balneari, DG7232, 110 m, herbassar humit en un sorral fluvial, J. Gesti, 16-IV-2021; Santa Coloma de Farners, al Veïnat d’Avall, DG7233, 125 m, herbassar ruderal al marge d’un petit rec agrícola, J. Gesti, 22-I-2021.
Cariofiŀlàcia pròpia de la Mediterrània meridional que és emprada en jardineria com a planta ornamental. Ens en consten poques indicacions a la meitat septentrional de Catalunya, on ha estat citada al Berguedà (Aymerich, 2016b) i també se’n conserven diversos testimonis d’herbari del Vallès Occidental i del Baix Llobregat (GBIF, 2021).

En una de les localitats que hem observat (Veïnat d’Avall) hi havia una desena d’individus a pocs metres d’un jardí on la planta havia estat cultivada. A l’altra localitat, en canvi, n’hem vist dos exemplars als sorrals de la riera, on podria haver arribat a través de llavors transportades per l’aigua (Fig. 10). Al territori estudiat és una aŀlòctona casual.
Soleirolia soleirolii (Req.) Dandy
Selva: Santa Coloma de Farners, a la font de la Pau (Castanyet), DG6837, 250 m, talús humit (HGI 24250, J. Gesti, 12-I-2021).
Es tracta d’una urticàcia originària de les illes de la Mediterrània occidental i d’Itàlia, que s’utilitza en jardineria com a planta de recobriment. A Catalunya ha estat indicada fora de cultiu al Bages, al Barcelonès, al Gironès, al Maresme i al Vallès (Casasayas, 1989; Juanola & Vilar, 1997; Bolòs, 1998; ORCA, 1999; Macías & Cebrián, 2018).
N’hem observat una única població, recobrint una superfície de poc menys d’1 m2, en talús humit sobre una font (Fig. 11). És casual al territori.

Bibliografia
Aymerich, P. & Sáez, L. 2015. Comentaris i precisions previs a la Checklist de la flora de Catalunya (nord-est de la península Ibèrica). Orsis, 29: 23-90.
Aymerich, P. 2016a. Notes florístiques de les conques altes dels rius Segre i Llobregat. IV. Orsis, 30: 133-165.
Aymerich, P. 2016b. Contribució al coneixement de la flora aŀlòctona del nord i el centre de Catalunya. Orsis, 30: 11-40.
Aymerich, P. 2017. Notes sobre flora aŀlòctona a Catalunya. Butlletí de la Institució Catalana d’Història Natural, 81: 97-116.
Aymerich, P. 2020. Notes sobre flora aŀlòctona a Catalunya, III. Butlletí de la Institució Catalana d’Història Natural, 84: 101124.
Batriu, E., Blanco-Moreno, J. M., Mercadé, A. & Pérez-Haase, A. 2012. Aportació al coneixement florístic de les Guilleries i del Collsacabra (Catalunya Oriental), III. Butlletí de la Institució Catalana d’Història Natural, 76: 147-157.
168 Butlletí ICHN 85 (4), 2021
Figura 9. Sedum palmeri, Santa Coloma de Farners.
Figura 10. Silene pseudoatocion, Santa Coloma de Farners.
Figura 11. Soleirolia soleirolii, Santa Coloma de Farners.
L. VILAR
J. GESTI PERICH &
SAIS
Bolòs, O. 1998. Atlas corològic de la flora vascular dels Països Catalans: Primera compilació general. Institut d’Estudis Catalans, Secció de Ciències Biològiques. Barcelona.
Bolòs, O., Nuet, J. & Panareda, J. M. 1986. Flora vascular del Montseny. P. 41-92. In: Terradas, J. & Miralles, J. (eds.). El patrimoni biològic del Montseny. Catàlegs de flora i fauna, 1. Diputació de Barcelona, Servei de parcs Naturals. 171 p.
Cadevall, J. 1915-1919. Flora de Catalunya. Vol. 2. Institut de Ciències. Barcelona. 470 p.
Casasayas, T. 1989. La flora aŀlòctona de Catalunya. Catàleg raonat de les plantes vasculars exòtiques que creixen sense cultiu al NE de la Península Ibèrica. Tesi Doctoral. Universitat de Barcelona. 892 p.
Cid, R. 1971. Guizotia abyssinica (L.) Cassini. Adventicia nueva para la flora española. Boletín de la Sociedad Castellonense de Cultura, 47: 36-37.
Cooperrider, T. S. 1984. Some species mergers and new combinations in the Ohio flora. The Michigan Botanist, 23(4): 173-176.
Domingues, J. & Da Costa, A. 2006. Acalypha rhomboidea Raf. (Euphorbiaceae), naturalized in portugal (new for Europe). Studia botanica, 25: 137-138
Folch, R. & Abellà, C. 1974. Galinsoga parviflora Cav. y Guizotia abyssinica (L.) Cass., dos adventicias nuevas para la flora catalana. Collectanea Botanica (Barcelona), 9: 183-189.
Font, X. 2021. Mòdul de Flora i Vegetació. Banc de Dades de Biodiversitat de Catalunya. Generalitat de Catalunya i Universitat de Barcelona. Disponible a: http://biodiver.bio.ub.es/biocat/ homepage.html [Data de consulta: 13 setembre 2021].
GBIF. 2021. Global Biodiversity Information Facility. Disponible a: https://www.gbif.org/ [Data de consulta: 13 setembre 2021].
Gesti, J. & Vilar, L. 2019. Aportacions al coneixement de la flora de les Guilleries orientals i àrees properes (nord-est de Catalunya). Butlletí de la Institució Catalana d'Història Natural, 83: 195198.
Gesti, J. & Vilar, L. 2020. Aportacions al coneixement de la flora de les Guilleries orientals i àrees properes (nord-est de Catalunya) –II. Butlletí de la Institució Catalana d'Història Natural, 84: 243248.
Girbal, J. 1984. Flora i vegetació del Gironès. Tesi Doctoral, Universitat Autònoma de Barcelona.
Gómez-Bellver, C., Nualart, N., Ibáñez, N., Burguera, C., Álvarez, H. & López-Pujol, J. 2019. Noves dades per a la flora aŀlòctona de Catalunya i del País Valencià. Butlletí de la Institució Catalana d'Història Natural, 83: 23-40.
Guardiola M. & Petit, A. 2020. Aportacions a la flora aŀlòctona de la serralada Litoral central catalana i territoris propers. Butlletí de la Institució Catalana d’Història Natural, 84: 35-49.
Iamonico, D. 2013. About the circumscription of Celosia argentea (Amaranthaceae) and the related Linnaean taxa. Phytotaxa, 90: 61-64.
Juanola, M. & Vilar, L. 1997. La flora vascular del massís de les Gavarres. Scientia Gerundensis, 23: 73-108.
Kobayashi, M & Furumoto, R. 2004. A phylogeny of Japanese dwarf bamboos, the Sasa-group based on RAPD- and morphological data analyses. Journal of Phytogeography and Taxonomy, 52: 1-24.
Levin, G. A. 1999. Notes on Acalypha (Euphorbiaceae) in North America. Rhodora 101(907): 217-233.
Levin, G. A. 2021. Acalypha. Flora of North America. Vol. 12. Euphorbiaceae. Disponible a: http://beta.floranorthamerica.org/ Acalypha [Data de consulta: 13 setembre 2021].
Macías, C. & Cebrian, J. 2018. Eruca vesicaria (L.) Cav. en els herbassars ruderals de Barcelona. Miconia, 2: 5-12.
Mallol, A. & Maynés, J. 2008. Nous xenòfits al Baix Empordà (Catalunya). Acta Botanica Barcinonensia, 51: 59-77
Ohwi, J. 1965. Flora of Japan. Smithsonian Institution. Washington DC. 1.067 p.
ORCA. 1999. Atlas corològic de la flora vascular dels Països Catalans. Vol. 6. Institut d’Estudis Catalans, Secció de Ciències Biològiques.
Pérez-Haase, A., Mercadé, A., Batriu, E. & Blanco-Moreno, J. M. 2013. Aportació al coneixement florístic de l’Espai Natural de les Guilleries-Savassona. Grup de Recerca de Geobotànica i Cartografia de la Vegetació. Universitat de Barcelona. Barcelona. 238 p.
Pié, G., Vilar, L. & Sánchez, S. 2010. Estudis corològics de plantes vasculars al Montseny. P. 115-118. In: Hernández, J.; Grau, J. & Melero, J. (coords.). VII Monografies del Montseny. Comunicacions presentades els dies 20 i 21 de novembre de 2008 a la VII Trobada d’Estudiosos del Montseny. Coŀlecció Documents de Treball. Sèrie Territori, 18. Diputació de Barcelona. 462 p.
POWO. 2021. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Disponible a: http://www. plantsoftheworldonline.org [Data de consulta: 13 setembre 2021].
Romo, A. M. 1987. Impatiens balfourii Hooker fil. i Polygonum orientale L. a la Selva. Butlletí de la Institució Catalana d’Història Natural, 54 (Sec. Bot., 6): 88-89.
Sáez, L., Galán De Mera, A., Pyke, S., Pié, G. & Carnicero, P. 2015. New data on vascular plants from Montseny massif (northeastern Iberian Peninsula). Orsis, 29: 205-230.
Sáez, L., Pié, G. & Carnicero, P. 2017. Catàleg de la flora vascular del massís del Montseny. Tres segles d’investigació botànica (1716-2016). Sèrie Territori i Parcs Naturals, 1. Coŀlecció Estudis. Diputació de Barcelona. 238 p.
Stace, C.A. 2019. New flora of the British Isles. (4 ed.). C&M Floristics. 1.300 p.
Suzuki, S. 1978. Index to Japanese Bambusaceae. Gakken Co., Ltd. Tokyo. 384 p.
Tutin, T. G., Heywood, V. H., Burges, N. A., Moore, D. M., Valentine, D. H., Walters, S. M. & Webb, D. A. 1968. Flora Europaea. Vol. 2. Rosaceae to Umbelliferae. Cambridge University Press. Cambridge. 469 p.
Vayreda, E. 1902. Plantas de Cataluña. Anales de la Sociedad Española de Historia Natural, sèrie 2, 10 (XXX): 491-582.
Verloove, F., Aymerich, P., Gómez-Bellver, C. & López-Pujol, J. 2019. Chorological notes on the non-native flora of the province of Tarragona (Catalonia, Spain). Butlletí de la Institució Catalana d’Història Natural, 83: 133-146.
Verloove, F. & Aymerich, P. 2020. Chorological novelties for the alien flora of northeastern Catalonia (Iberian Peninsula). Butlletí de la Institució Catalana d’Història Natural, 84: 137-153.
Verloove F. 2021. Euphorbiaceae. Manual of the Alien Plants of Belgium. Botanic Garden Meise, Belgium. Disponible a: https:// alienplantsbelgium.be [Data de consulta: 13 setembre 2021].
Vilar, L. 1987. Flora i vegetació de la Selva. Tesi doctoral. Universitat Autònoma de Barcelona. Bellaterra. 608 p.
169 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA
APORTACIONS AL CONEIXEMENT DE LA FLORA DE LES GUILLERIES ORIENTALS – III
Principals resultats de la campanya micològica russo-catalana als Pirineus catalans occidentals: prats alpins i subalpins i boscos subalpins
Jordi Vila*, Olga Morozova**, Anna Fedosova**, Tània Jiménez-Palomar*** & Xavier Llimona****
* Passatge del Torn, 4. 17800 Olot. Catalonia.
** Komarov Botanical Institute RAS. Laboratory of the Systematics and Geography of Fungi. 2 Prof. Popov Str., St Petersburg. Russia.
*** Societat Catalana de Micologia. Jordà, 10, baixos 2. 08034 Barcelona. Catalonia.
**** Institut de Recerca en Biodiversitat (IRBIO, CeDocBiv). Dept de Biologia Evolutiva, Ecologia i Ciències Ambientals (BEECA). Secció de Botànica i Micologia. Universitat de Barcelona. Diagonal, 643. 08028 Barcelona. Catalonia.
Autor per a la correspondència: Jordi Vila. A/e: entoloma.catalonia@gmail.com
Rebut: 10.09.2021; Acceptat: 04.11.2021; Publicat: 30.12.2021
Resum
Es va organitzar una visita de dos micòlogues de l’Institut Komarov (Sant Petersburg), que va coincidir amb unes condicions especialment favorables per a l’estudi dels fongs de comunitats alpines i subalpines, especialment prats i rasos. Entre la vall de Ribes (Pardines i Núria, Girona) i l’alta vall de la Noguera Pallaresa (Lleida), es prospectaren nombroses localitats, entre 1300 i 2200 m s.n.m. Una selecció de les espècies més interessants és la següent: Bryoglossum gracile, Hodophilus foetens, Entoloma azureopallidum, E. aranense, E. catalaunicum, E. glaucobasis, E. glaucodubium, E. linkii, E. minutigranulosum, E. montanum, E. muscoalpinum, E. porphyrogriseum, Hygrocybe turunda, Rhizomarasmius epidryas, Rhodophana nitellina
Mots clau: Fongs, Ascomicots, Basidiomicots, Pirineu, prats alpins/subalpins, geoglossàcies, Entoloma.
Abstract
Main results of the Russian-Catalan mycological survey, in the western Catalan Pyrenees: alpine and subalpine belt
A visit of two mycologists from the Komarov Institute (St. Petersburg) was organized, which coincided with particularly favorable conditions for the study of fungi from alpine and subalpine communities, especially meadows. Between the valley of Ribes (Pardines and Núria, Girona) and the high valley of the Noguera Pallaresa (Lleida), numerous localities were prospected, between 1300 and 2200 m a.s.l. A selection of the most interesting species is as follows: Bryoglossum gracile, Hodophilus foetens, Entoloma azureopallidum, E. aranense, E. catalaunicum, E. glaucobasis, E. glaucodubium, E. linkii, E. minutigranulosum, E. montanum, E. muscoalpinum, E. porphyrogriseum, Hygrocybe turunda, Rhizomarasmius epidryas, Rhodophana nitellina
Key words: Fungi, Ascomycota, Basidiomycota, Pyrenees, alpine/subalpine meadows, Geoglossaceae, Entoloma.
Introducció
La preocupació de Pius Font i Quer per l’estudi de la biodiversitat micològica de Catalunya és ben coneguda (Llimona, 2009: 136-140). Aviat coŀlaborà amb el metge Joaquim Codina (Codina & Font i Quer, 1931) i, més endavant invità diversos especialistes europeus, especialment Maire i Heim (Font i Quer, 1937). Quan finalment aconseguí contractar un micòleg de prestigi, Rolf Singer, com a membre fix del seu equip, li marcà com a principal objectiu l’estudi dels fongs d’una tria ben representativa de localitats dels Pirineus de Lleida. Singer es posà a la feina, el 12 de juliol de 1934 (Llimona, 1997), i explorà principalment localitats de la Vall d’Aran, passant després al Pallars Sobirà i a la vall d’Àneu. Retornat a Barcelona a l’agost, pujà de nou a l’Aran, del 23 de setembre a principis d’octubre.
Una petita part dels notables resultats científics (basats en més de 400 recoŀleccions) obtinguts en tan poc temps per Singer, va poder ser publicada abans de la guerra civil (Singer, 1935, 1936a, 1936b, 1936c), però la desgraciada detenció per motius polítics, el 12 d’octubre del mateix any de la seva arribada, truncà una campanya micològica enormement productiva i, amb ella, el projecte d’iniciar una escola micològica catalana puntera. El principal gruix dels resultats, redactat en francès, no apareixerà publicat fins després de la guerra (Singer, 1947).
L’alliberament de Singer es fa poc abans del 13 de novembre de 1935, al preu de la seva expulsió immediata del país, i aviat el trobem a París i, finalment, a Leningrad (nom que duia aleshores Sant Petersburg) on, el 22 de maig de 1935, ja treballa com a investigador, al Jardí Botànic de l’Acadèmia de Ciències, actualment Institut Botànic Komarov de Sant
171 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA GEA, FLORA ET FAUNA
de la Institució Catalana d’Història Natural, 85 (4): 171-186. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition)
10.2436/20.1502.01.90
Butlletí
DOI:
GEA,
Petersburg. Hi romandrà fins al 1941. Font i Quer l’ajuda econòmicament, i s’ocupa d’enviar-li després els seus llibres, equipatges i exsiccates de fongs. Les mostres així salvades seran incorporades a l’herbari LE.
La represa fou lenta, però els micòlegs catalans, agrupats en la Societat Catalana de Micologia, en el darrer decenni del segle passat esdevenen molt actius en els estudis dels fongs pirenencs (Esteve-Raventós, 1978; Rocabruna et al., 1994, 1996; Llistosella et al., 1994, 1996, 1998; Vidal & Vila, 1994; Vidal, 1994, 1997; Ballarà, 1995, 1996, 1997, 1998, 1999; Bon & Ballarà, 1995, 1996; Ballarà & Escànez, 1999; Vila et al., 1997, 1998, 2001, 2006, 2013; Galán et al., 1997; Esteve-Raventós & Vila, 1997, 1998; Llistosella, 1998; Gorris et al., 1999; Esteve-Raventós et al., 1999, 2001a, 2001b; Muntañola-Cvetkovic et al., 1997; Quadrada et al., 1997; Tabarés et al., 2004; Llorens-van-Waveren & Llistosella, 2004, 2005, 2006; Vila & Caballero, 2007, 2009). No esmentem treballs més tardans, però aquests són suficients per posar de manifest la represa de l’interès de Catalunya per la micologia dels Pirineus i els interessants resultats aconseguits.
No tenim més dades sobre la relació entre l’Institut Komarov i Catalunya, fins que el coneixement dels treballs catalans per part d’una de nosaltres (O.M.), que ja havia coŀlaborat amb el primer autor (J.V.) (Morozova et al., 2014), la decidí a proposar una exploració micològica d’estiu als Pirineus catalans, visitant algunes de les localitats on havia estat Singer, entre altres. Vam acceptar amb interès, i planificàrem una exploració de baix cost, assumible reunint els recursos que s’esmenten als Agraïments.
Les dates escollides van coincidir amb un moment especialment favorable, després de pluges de primavera i d’estiu, que induïren una extraordinària formació de carpòfors, especialment en els prats alpins i subalpins, els més rics en espècies del gènere Entoloma, en el qual tenien especial interès dos dels expedicionaris (O.M. i J.V.). Una tercera autora (A.F.), interessada en geoglossàcies, intervingué també en la prospecció d’altres grups.
Materials i mètodes
Les mostres es van recollir i / o registrar seguint el mètode de ruta a les ubicacions que es mostren a continuació. Al diari es van anotar les espècies comunes, que no tenien cap mena de dubte. Preferentment, es van recollir i preparar per herbari els exemplars que exigien una identificació addicional. Les característiques macromorfològiques es van estudiar a partir de coŀleccions fresques així com mitjançant l’anàlisi de les fotos realitzades al camp (Figs. 2-6). Es van estudiar caràcters microscòpics amb un microscopi Zeiss Axioscope A1 amb càmera AxioCam 1Cc 3 i les eines de programa AxioVisionRel.4.6 (Carl Zeiss, Alemanya). Es van observar basidiòspores, basidis i cistidis himenials en preparacions per compressió (squash) de petites parts de les làmines en KOH al 5 %. La pileipeŀlis es va examinar en una secció radial del píleu, l’estipitipeŀlis en una llesca longitudinal en KOH al 5 %. Per a la identificació es van utilitzar treballs clàssics (Noordeloos, 1992, 2004; Knudsen & Vesterholt, 2012), així com
treballs moderns sobre els Pirineus (Caballero & Vila, 2013; Vila & Caballero, 2007, 2009; Vila et al., 2013; Corriol, 2016). Hem obtingut dades moleculars (ITS, LSU) de totes les coŀleccions d’Entoloma del present article. Les seqüències seran publicades en un proper treball (Dima et al., 2022, en premsa). Pels fongs, hem seguit Mycobank (consultat fins al 27.7.2021). El material testimoni es conserva a l’herbari LE, del Komarov Botanical Institute of Russian Academy of Sciences, Sant Petersburg, Rússia, i en l’herbari del primer autor (JVG). Hem inclòs algunes coŀleccions addicionals obtingudes dies abans (o després) de la campanya principal, normalment en prospeccions on verificàrem l’evolució de les localitats per poder fer una millor tria de les mateixes i assegurar un bon nombre de recoŀleccions. Aquestes mostres apareixen indicades, en el text, amb un asterisc (*).
Localitats i caracterització micològica dels hábitats
Bona part de les espècies observades i identificades durant la prospecció han estat anotades, juntament amb un índex que reflecteix l’abundància de carpòfors observada, i un segon índex que expressa la seva sociabilitat (Taula 1). Durant cada recorregut d’exploració, s’ha valorat la presència de cada tàxon detectat mitjançant els dos índexs de BraunBlanquet (recobriment i sociabilitat) adaptats a l’estudi d’una comunitat de fongs (Darimont, 1975; Vila & Llimona, 1999). En aquest cas, l’índex de recobriment es substituïa per una estimació de l’abundància, entesa com el nombre aproximat de carpòfors en una superfície d’uns 400-4000 m2. Les categories de l’índex d’abundància són les següents:
+ =1 carpòfor o un petit grup dens de carpòfors; 1 = de 2 a 3 carpòfors; 2 = de 4 a 10 carpòfors; 3 = d’11 a 25; 4 = de 25 a 50; 5 = més de 50.
El segon índex, el de sociabilitat, ens informa sobre el grau d’agrupament dels carpòfors, segons l’escala següent: 1 = carpòfors isolats (no es posa quan n’hi ha un de sol); 2 = grups de 2 a 4 carpòfors; 3 = grups de 5 a 10 carpòfors; 4 = grups d’11 a 50 carpòfors; 5 = grups de més de 50 carpòfors.
Malgrat les seves limitacions, el llistat així indexat dona una idea de la comunitat detectada a cada àrea explorada, més precisa que la d’un llistat simple.
Ordenació cronològica i segons el número de localitat que figura als mapes (Fig. 1). Les espècies identificades o estudiades al laboratori s’inclouen en l’apartat de resultats:
22.08.2014. Port del Cantó. Parc Natural de l’Alt Pirineu. Comarca: Pallars Sobirà. Municipi: Soriguera.
1. Abans de Port del Cantó (venint de La Seu d’Urgell, en direcció a Sort). Bosc de Pinus sylvestris (exploració de curta durada).
2. Port del Cantó. 1800 m. Prats amb pendent suau N-NW, i roques calcàries o gresos vermells , prop d’un bosc de Pinus sylvestris
23.08.2014. D’Alós d’Isil al pont de Marimanya. Parc Natural de l’Alt Pirineu. Comarca: Pallars Sobirà. Municipi: Alt Àneu.
3. Pista a Montgarri, prop de la bifurcació al Port de Salau. N 42.732222°, E 001.1100°, alt. 1375 m. Prat amb
172 Butlletí ICHN 85 (4), 2021
J. VILA ET AL.
FLORA ET FAUNA
Eryngium bourgatii, Trifolium i algunes Betula, sobre roca calcària.
4. Pista a Montgarri, prop del pont de la Perosa. N 42.753611°, E 001.080833°, alt. 1513 m.
5. Pista a Montgarri, després de la Perosa. N 42.757778°, E 001.07500°, alt. 1550 m.
6. Pista a Montgarri. Bosc ombrívol d’Abies alba, amb un curs d’aigua que ve del vessant N. N 42.759722°, E 001.047222°, alt. 1582 m.
23.08.2014. De pont de Marimanha al Pla de Beret. Parc Natural de l’Alt Pirineu. Comarca: Era Val d’Aran. Municipi: Naut Aran.
7. Primers prats tot just entrant a la Vall d’Aran, amb Gentiana lutea. N 42.759167°, E 001.019444°, alt. 1606 m.
8. Davant de Montgarri. N 42.759167°, E 000.996389°, alt. 1650 m. Roques silícies.
9. Pla de Beret. Sobre roca àcida i bàsica. N 42.733889°, E 000.976667°, alt. 1796 m. Prats amb Pinus uncinata
23.08.2014. Marge oest del Port de la Bonaigua. Parc Nacional d’Aigüestortes i Estany de Sant Maurici. Comarca: Pallars Sobirà. Municipi: Espot.
10. Port de la Bonaigua. N 42.663611°, E 000.978333°, alt. 2052–2066 m.
24.08.2014. Parc Nacional d’Aigüestortes i Estany de Sant Maurici. Comarca: Pallars Sobirà. Municipi: Espot.
11. Circ dels Estanyets. Sobre Espot. N 42.5375°, E 001.0700°, alt. 2148 m. Prats rasos sobre roca calcària.
11a. Ibid. Alt. 2218 m. Comunitat de Dryas octopetala i Vaccinium uliginosum
12. Sobre Espot, pista a l’estació d’esquí. N 42.565833°, E 001.092778°, alt. 1494 m. Betula pendula i prats. Morrena amb materials calcaris i silícics.
24.08.2014. Marge oest del Port de la Bonaigua. Parc Nacional d’Aigüestortes i Estany de Sant Maurici. Comarca: Pallars Sobirà. Municipi Espot.
13 (10). Port de la Bonaigua (segona prospecció), 18 h després de la primera. N 42.663611°, E 0.978333°, alt. 2052 m.
14. Bosc del Gerdar N 42.642222°, E 001.0225°, alt. 1600 m. Bosc d’Abies alba
14a. Bosc del Gerdar. Banda talada sota la línia elèctrica, poblada amb herbes graminoides altes. N 42.642417°, E 001.023417°, alt. 1596 m.
25.08.2014. Parc Nacional d’Aigüestortes i Estany de Sant Maurici. Comarca: Pallars Sobirà. Municipi: Espot.
15. Plaça dels Arbres. N 42.582222°, E 001.035556°, alt. 1665 m. Bosc vell d’Abies alba, amb Betula pendula, amb prat a les clarianes.

173 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA PRINCIPALS RESULTATS DE LA CAMPANYA MICOLÒGICA RUSSO-CATALANA ALS PIRINEUS CATALANS OCCIDENTALS
Figura 1. Situació general de les localitats prospectades. Relació numèrica de les localitats al text.
GEA, FLORA ET FAUNA
Taula 1. Índex d’abundància per localitat.
174 Butlletí ICHN 85 (4), 2021
Localitats 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 Adelphella babingtonii + Agrocybe pediades +.2 + + 1.1 Amanita crocea + + Amanita excelsa + Amanita muscaria + Amanita pantherina + Arrhenia cf. acerosa +.2 Auriscalpium vulgare + Atheniella flavoalba +.2 Bovista aestivalis + + Bovista plumbea + Bryoglossum gracile +.3 Butyriboletus appendiculatus + Caloboletus calopus + Calocera cornea +.3 Calocybe carnea + Calyptella capula + Cantharellus cibarius + Chalciporus amarellus 1.2 Chlorociboria aeruginascens +.2 Chroogomphus rutilus 2.2 1.1 Clavulina cinerea 1.2 Clavulina rugosa +.2 + Clitocybe bresadolana +.2 Clitocybe fragrans 2.2 Clitocybe gibba 1.2 2.2 +.3 Clitocybe nebularis +.3 Clitocybe odora + + 2.2 Clitopilus prunulus +.2 2.1 Clitopilus scyphoides + 2.1 2.2 1.2 +.2 +.2 Collybia cirrhata +.2 Conocybe sp. 1.3 Coprinellus micaceus +.3 Cortinarius delibutus f. dryad. 2.2 Crinipellis scabella +.2 + +.2 Cudonia circinans 1.4 Cuphophyllus fornicatus +.2 Cuphophyllus pratensis +.2 +.2 1.2 Cuphophyllus virgineus + 1.1 + 1.2 +.2 Cystoderma amianthinum +.3 Cystolepiota bucknallii +.2 Cystolepiota hetieri + Cystolepiota sistrata 2.3 Dermoloma cuneifolium +.3 +.3 Entoloma anatinum +.2 1.1 2.2 Entoloma aranense +.2 Entoloma asprellum 1.2 +.2 +.2 +.2 Entoloma azureopallidum 2.2 Entoloma brunneoserrulatum +.2 +.2 Entoloma catalaunicum 2.3 Entoloma chalybaeum +.2 +.2 2.2 +.2 +.2 Entoloma clandestinum +.2 +.2 + 2.1 +.3 2.2 + + + + 1.2 Entoloma conferendum +.2 Entoloma exile 2.2 Entoloma formosum +.2 Entoloma fridolfingense 1.2 Entoloma glaucobasis + +.2 +.2 Entoloma glaucodubium 1.2 Entoloma griseocyaneum +.2 +.2 2.2 1.3 2.2 +.2 Entoloma incanum 2.3 + + Entoloma infula +.3 2.3 + Entoloma juncinum 1.1 Entoloma majusculum s.l. 2.2 2.2 2.2 Entoloma mediterraneense +.2 Entoloma mougeotii + + Entoloma muscoalpinum 1.3 Entoloma poliopus +.2 Entoloma porphyrogriseum +.2 +.2 1.2 + Entoloma prunuloides + Entoloma querquedula +.2 Entoloma sericellum s.l. +.2 +.3 +.2 Entoloma sericeum +.2 +.2 + + 2.1 1.2 2.2 +.2 J. VILA ET AL.
175 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA Localitats 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 Entoloma serrulatum 2.3 +.2 +.3 2.2 +.2 Entoloma turci 2.2 Entoloma undatum s.l. +.2 1.1 1.2 1.1 2.2 Entoloma viiduense + Entoloma violaceoserrulatum +.2 Entoloma weholtii +.2 + 2.2 1.2 2.2 Exidia saccharina +.2 Exobasidium rhododendri + 2.1 Galerina marginata +.3 Geastrum campestre +.3 Geastrum fimbriatum + Geastrum triplex + 2.3 Geoglossum cf. vleugelianum +.3 Geoglossum fallax +.3 +.2 Geoglossum sp. + 2.1 Gliophorus irrigatus 1.3 Gliophorus psittacinus +.2 +.4 Glutinoglossum sp. + Gymnopilus picreus 2.3 Gymnopus androsaceus +.2 Gymnopus confluens +.3 Gymnopus dryophilus +.2 Hebeloma sinapizans +.3 Hebeloma sp 1.2 Helvella gr. acetabulum + Helvella atra + 1.3 Helvella cupuliformis + Helvella elastica + + Helvella fibrosa + +.2 Helvella lacunosa +.3 Helvella macropus +.2 +.2 +.3 Hericium coralloides +.2 Hodophilus foetens +.3 Humaria hemisphaerica 1.3 Hygrocybe coccinea +.2 Hygrocybe conica 1.3 + 1.1 + + Hygrocybe miniata +.2 Hygrocybe reai +.3 + Hygrocybe turunda +.2 +.2 Hygrophorus chrysodon + Hygrophorus pudorinus +.2 Hymenoscyphus conscriptum +.4 Hymenoscyphus fructigenus +.3 Hypocrea leucopus + Hypoxylon howeanum +.2 Inocybe geophylla + 2.3 3.2 Inocybe lacera s.l. +.2 Inocybe sp. 1.2 Laccaria amethystea +.2 Laccaria laccata +.3 1.2 Lactarius aurantiacus 1.3 + Lactarius glyciosmus + 1.2 + Lactarius necator + Lactarius pyrogalus + + + + +.2 Lactarius salmonicolor + + + Lactarius spinosulus + Lactarius torminosus + Leccinellum pseudoscabrum 1.1 Leotia lubrica 1.3 +.2 2.2 Lepiota aspera 1.2 Lepiota castanea + Lepiota clypeolaria + + + Lepiota cristata +.2 + +.2 Lepiota magnispora +.2 Lepista panaeolus +.2 Leucocortinarius bulbiger + Leucocybe connata +.3 Lichenomphalia umbellifera + +.2 Lycoperdon echinatum +.2 + Lycoperdon excipuliforme +.2 + + + Lycoperdon utriforme 2.1 Marasmiellus vaillantii +.3 Marasmius buxi 2.3 Marasmius oreades 1.3 +.3 +.2 +.3
CAMPANYA MICOLÒGICA RUSSO-CATALANA ALS PIRINEUS CATALANS OCCIDENTALS
PRINCIPALS RESULTATS DE LA
GEA, FLORA ET FAUNA
176 Butlletí ICHN 85 (4), 2021
Localitats 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 Marasmius rotula 2.2 2.4 Marasmius torquescens + 3.2 Marasmius wynneae +.4 Megacollybia platyphylla 3.1 Melanoleuca brevipes 1.1 Melanophyllum haematospe. +.2 + 1.2 Meottomyces dissimulans 1.2 Microglossum olivaceum +.2 Mutinus caninus + Mycena acicula + Mycena megaspora +.2 Mycena pura +.2 2.3 + +.3 2.3 + Mycena renati +.3 Mycena stylobates + Mycetinis scorodonius 1.2 Otidea alutacea +.2 Otidea sp. + Panaeolus fimicola +.2 + +.2 Phallus impudicus + 3.1 Pholiota squarrosa +.4 Plicaturopsis crispa +.4 +.4 3.3 Pluteus romellii + Pluteus thomsonii + + Polyporus squamosus + Porphyrellus porphyrosporus + Protostropharia semiglobata + 1.1 1.1 Psilocybe coronilla 1.1 Psilocybe inquilina +.2 Ramaria fennica s.l. +.2 Ramaria largentii +.2 + Ramaria stricta +.2 Rhizomarasmius epidryas 3.2 Rhizopogon roseolus +.2 Rhodophana nitellina 2.2 Ripartites tricholoma +.2 Rugosomyces carneus +.1 + Russula consobrina 1.1 Russula sanguinaria + Scutellinia barlae +.2 Scutellinia scutellata +.2 Sebacina incrustans +.2 Spathularia flavida 1.2 +.3 1.3 Stereum hirsutum 1.5 Suillus granulatus 1.2 1.2 Suillus luteus 2.2 + Taphrina pruni +.3 Tapinella atrotomentosa +.2 Tarzetta cupularis +.2 + +.3 Tephrocybe rancida + Terana caerulea +.5 Thelephora caryophyllea +.3 Thelephora penicillata +.2 Tremiscus helvelloides + Trentepohlia aurea +.2 Trichoglossum hirsutum 1.3 +.2 Tricholoma bufonium + + +.4 Tricholoma fulvum + Tricholoma imbricatum + Tricholoma saponaceum + +.2 Tricholoma scalpturatum + +.2 Tricholoma sulphureum + +.2 Tricholomopsis decora 1.2 Tricholomopsis ornata + Trichophaea woolhopeia +.2 + Typhula sp. +.3 Vascellum pratense 1.1 Xerocomus subtomentosus s.l. 1.1 Xylaria hypoxylon +.2 Xylaria polymorpha +.2 J. VILA ET AL.

27.08.2014. Núria. Parc Natural de les Capçaleres del Ter i del Freser. Comarca: Ripollès. Municipi: Queralbs.
16. Núria, vora Santuari, prat, sobre substrat àcid. N 42.396361°, E 2.155417°, alt. 1950 m.
16a. Núria, principi de la Coma de l’Embut, prat bàsic, entre l’herba o Dryas octopetala. N 42.399814°, E 002.141859°, alt. 2150 m.
29.08.2014. Parc Natural de les Capçaleres del Ter i del Freser. Comarca: Ripollès. Municipi: Pardines.
17. Pardines i Serra Cavallera. Sota Corylus avellana, sobre sòl bàsic.
18, 19, 20. Pardines. Torrent de Burgil. N 42.3000°, E 002.236667°, 1325 m.
18. Llit de roca calcària.
19. Prat baix amb herbes curtes i denses.
20. Buxus sempervirens, matollar dens.
21, 22, 23. Pardines. Prop de Coll del Bac. N 42.2975°, E 002.225833°, alt. 1432 m.
21. Primera parada (final de la pista forestal).
22. Segona parada. Prop del riu. N 42.2975°, E 2.227222°, alt. 1461 m. Prat.
23. Prop del riu, amb Buxus, Corylus i Fraxinus, amb punts amb prat baix.
Resultats
Ascomycota
Geoglossomycetes
Geoglossales
Geoglossaceae
Geoglossum fallax E.J. Durand - Port de la Bonaigua (13), prat alpí, entre gramínies i Calluna vulgaris, sobre sòl àcid, 24 VIII 2014; Núria, principi de la Coma de l’Embut (16a), prat, sobre sòl bàsic, 27 VIII 2014 (LE 304016) (Fig. 6c).
Geoglossum cf. vleugelianum Nannf. - València d’Àneu (14a), 1596 m, sota línia d’alt voltatge, herbes graminoides altes, sobre sòl, 24 VIII 2014 (LE 304012).
Trichoglossum hirsutum (Pers.) Boud. - Plaça dels Arbres (15), entre gramínies, vora Plantago media i Trifolium sp., 25 VIII 2014 (LE 303902); Núria, vora Santuari (16), prat, entre gramínies, sobre sòl àcid, 27 VIII 2014 (LE 303903).
Leotiomycetes
Helotiales
Chlorociboriaceae
Chlorociboria aeruginascens (Nyl.) Kanouse ex C.S. Ramamurthi, Korf et L.R. Batra - Pardines (22), amb Buxus, Corylus i Fraxinus, 29 VIII 2014.
Helotiaceae
Bryoglossum gracile (P. Karst.) Redhead - Pla de Beret (9), sobre molses de la riba d’un rierol, 23 VIII 2014 (LE 304006) (Fig. 2).
Leotiaceae
Microglossum olivaceum (Pers.) Gillet sensu lato - Pardines (19), prat baix, amb herbes curtes i denses, sobre sòl, 29 VIII 2014.
Rhytismatales
Cudoniaceae
Cudonia circinans (Pers.) Fr. - Plaça dels Arbres (15), bosc d’Abies alba, terrícola, 25 VIII 2014 (LE 304015).
Spathularia flavida Pers. - Port del Cantó (2), sota Pinus sylvestris, sobre sòl bàsic, 22 VIII 2014 (LE 303997, LE 303998); Bosc del Gerdar de Sorpe (14), sobre sòl, bosc d’Abies alba, 24 VIII 2014 (LE 304012); Plaça dels Arbres (15), sobre sòl en bosc d’Abies alba , 25 VIII 2014 (LE 304014).
177 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA
Figura 2. Bryoglossum gracile (LE 304006) Foto: Fedosova.
PRINCIPALS RESULTATS DE LA CAMPANYA MICOLÒGICA RUSSO-CATALANA ALS PIRINEUS CATALANS OCCIDENTALS
GEA, FLORA ET FAUNA
Pezizomycetes
Pezizales
Helvellaceae
Helvella gr. acetabulum (L.) Quél. - Pardines (22), amb Buxus, Corylus i Fraxinus, sobre sòl, 29 VIII 2014.
Helvella atra J. König - Pista a Montgarri (4), prat subalpí, sobre sòl bàsic, 23 VIII 2014 (LE 304001); Pardines (22), 1461 m, amb Buxus, Corylus i Fraxinus, sobre sòl, 29 VIII 2014.
Helvella cupuliformis Dissing et Nannf. - Pla de Beret (9), sobre sòl, vora rierol, 23 VIII 2014 (LE 304007).
Helvella elastica Bull. - Plaça dels Arbres (15), bosc mixt (Abies alba i Betula pendula), sobre sòl, 25 VIII 2014 (LE 304013).
Helvella fibrosa (Wallr.) Korf - Espot, sobre Santa Maria (12), a la pineda de Pinus uncinata, sobre sòl vora rierol, 24 VIII 2014 (LE 304009) (Fig. 6d); Coll del Bac (21), sota Buxus sempervirens de mida gairebé arbòria, 29 VIII 2014.
Helvella lacunosa Afzel. - Vora Borda de Perosa (4), prat subalpí a la riba d’un rierol, sobre sòl bàsic, 23 VIII 2014 (LE 304002).
Helvella macropus (Pers.) P. Karst. - Espot, sobre Santa Maria (12), en un bedollar (Betula pendula), vora un rierol, 24 VIII 2014 (LE 304010); Plaça dels Arbres (15), bosc d’Abies alba , sobre sòl, 25 VIII 2014.
Pezizaceae
Adelphella babingtonii (Berk. et Broome) Pfister, Matočec et I. Kušan - Vora borda de la Perosa (4), riba d’un rierol, sobre branques caigudes d’Abies sp., 23 VIII 2014 (LE 304003).
Peziza succosella (Le Gal et Romagn.) M. M. Moser ex Aviz.-Hersh. et Nemlich - Espot, sobre Sta Maria (12), sota Betula pendula, a la riba d’un rierol, sobre sòl, 24 VIII 2014 (LE 304011).
Peziza cf. succosella (Le Gal et Romagn.) M.M. Moser ex Aviz.-Hersh. et Nemlich - Pista a Montgarri (6), bosc d’Abies alba, sobre sòl, 23 VIII 2014 (LE 304004); Pardines (22), 1461 m, amb Buxus, Corylus i Fraxinus, sobre sòl, 29 VIII 2014.
Pyronemataceae
Scutellinia barlae (Boud.) Maire - Circ dels Estanyets (11), prat aigualós, vora un rierol, sobre sòl bàsic, 24 VIII 2014 (LE 304008).
Tarzetta cupularis (L.) Lambotte - Borda de Tariou (3), marge d’un rierol, sobre sòl, 23 VIII 2014 (LE 304000); Pardines (18), sobre sòl, 29 VIII 2014.
Trichophaea woolhopeia (Cooke et W. Phillips) Boud.Pla de Beret (9), marge d’un rierol, sobre sòl, 23 VIII 2014 (LE 304005); Pardines (18), 1325 m, sobre sòl, 29 VIII 2014.
Sordariomycetes
Hypocreales
Hypocreaceae
Hypocrea leucopus P. Karst. - Port del Cantó (2), sota Pinus sylvestris, sobre sòl bàsic, 22 VIII 2014 (LE 303999).
Bsidiomycota
Agaricomycetes
Agaricaceae
Lycoperdon excipuliforme (Scop.) Pers. - Davant de Montgarri (8), 23 VIII 2014; Circ dels Estanyets (11), 24 VIII 2014; Núria, vora Santuari (16), prat, sobre sòl àcid, 27 VIII 2014; Pardines (22), 27 VIII 2014.
Clavariaceae
Hodophilus foetens (W. Phillips) R. Heim - Port del Cantó (2), prat, sobre sòl bàsic, 22 VIII 2014.
Entolomataceae
Entoloma anatinum (Lasch) Donk - Port de la Bonaigua (13), prat alpí, entre gramínies i Calluna vulgaris, sobre sòl àcid, 24 VIII 2014 (JVG 1140824-19, LE 312421; JVG 1140824-21, LE 312422); Plaça dels Arbres (15), clariana en el bosc mixt d’Abies alba i Betula pendula, 25 VIII 2014 (JVG 1140825-3, LE 312423) (Fig. 3a); Núria, vora Santuari (16), prat, entre gramínies, sobre sòl acid, 27 VIII 2014 (JVG 1140827-16, LE 312424).
Entoloma aranense F. Caball. et Vila - Núria, principi Coma de l’Embut (16a), prat, sobre sòl basic, 27 VIII 2014 (JVG 1140827-14, LE 312428) (Fig. 3b); *ibid., vora santuari, prat bàsic, 8 VIII 2014 (JVG 1140808-3). Comentaris: genèticament molt proper a Entoloma coracis, una espècie de recent descripció (Crous et al., 2021), de la qual se’n separa pels colors més pàŀlids, el port menys esvelt i un hàbitat estrictament alpí o subalpí.
Entoloma asprellum (Fr.) Fayod - Montgarri (4, 5, 8), prat, sobre sòl àcid, 23 VIII 2014 (JVG 1140823-15, LE 312425); Núria (16a), vora Santuari, prat, sobre sòl àcid, 27 VIII 2014 (JVG 1140827-8, LE 312426); *ibid., 30 VIII 2013 (JVG 1130830-14) (Fig. 3c); *ibid., Ras de l’Ortigar, 2220 m, vora Salix retusa, 8 VIII 2014 (JVG 1140808-7); *Alp, Baga de Saltèguet, 1760 m, en un prat bàsic, 8 VII 2014 (JVG 1140708-14, JVG 1140708-16); *Toses, Coll de la Creueta, 2090 m, en un prat, entre gramínies, 5 VIII 2014 (JVG 1140805-18). Comentaris: molt abundant a tota la zona del Pirineu, gairebé sempre en prats bàsics alpins o subalpins. Es tracta d’un tàxon molt variable macroscòpicament, no sempre fàcil d’identificar i que pot ser confós, per exemple, amb Entoloma glaucobasis
Entoloma azureopallidum Corriol - Circ dels Estanyets (11a), prat alpí, sobre sòl bàsic, entre Dryas octopetala, 24 VIII 2014 (JVG 1140824-4, LE 312505) (Fig. 3d). Comentaris: nou per Catalunya.
Entoloma brunneoserrulatum Eyssart. et Noordel. - Port del Cantó (2), prat bàsic, entre l’herba, 22 VIII 2014 (JVG 1140822-11); Núria, principi Coma de l’Embut (16a), prat bàsic, entre l’herba, 27 VIII 2014 (JVG 1140827-21). Comentaris: l’aresta bruna de les làmines permet la seva separació respecte Entoloma serrulatum i tàxons propers. Entoloma brunneoserrulatum pot fructificar tant en prats com en boscos.
Entoloma catalaunicum (Singer) Noordel. - Port del Cantó (2), al prat, al marge i clarianes del bosc de Pinus sylvestris,
178 Butlletí ICHN 85 (4), 2021
J. VILA ET AL.
PRINCIPALS






312420)
Morozova;
sobre sòl bàsic, 22 VIII 2014 (JVG 1140822-2, LE 312420) (Fig. 3e). Comentaris: tot i l’epitet específic, no es gaire freqüent al nostre país. Ben caracteritzat pel bonic to rosat del píleu i per l’estípit amb colors més o menys blavosos. Una coŀlecció trobada per sobre dels 2600 m d’altitud al Pirineu de Lleida no ha estat conservada (S. Català, comunicació personal).
Entoloma chalybaeum (Pers.) Noordel. - Vora borda de la Perosa (4), prat subalpí, sobre sòl bàsic, 23 VIII 2014 (JVG 1140823-4, LE 312429; JVG 1140823-5, LE 312507); vora Pont de Marimanya (7), prat supalpí, sobre sòl bàsic, 23 VIII
2014 (JVG 1140823-10, LE 312430); Pla de Beret (9), prat sobre sòl bàsic, 23 VIII 2014 (JVG 1140823-17, LE 312431); Núria, principi Coma de l’Embut (16a), prat bàsic, entre l’herba, 27 VIII 2014. Comentaris: molt abundant en prats, es caracteritza pel color blau fosc del basidioma. Entoloma rivipollense, de recent descripció (Vila et al., 2021) pot presentar un color semblant però es separa pels caràcters microscòpics i l’hàbitat en boscos.
Entoloma clandestinum (Fr.) Noordel. - Port del Cantó (2), prat sobre sòl bàsic, 22 VIII 2014 (JVG 1140822-9, LE 312450); vora borda de la Perosa (4), prat subalpí, so-
179 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA
Figura 3. a) Entoloma anatinum (LE 312423) Foto: Morozova; b) Entoloma aranense (LE 312428) Foto: Morozova; c) Entoloma asprellum (JVG 1130830-14) Foto: Vila; d) Entoloma azureopallidum (LE 312505) Foto: Morozova; e) Entoloma catalaunicum (LE
Foto:
f) Entoloma exile (LE 312434) Foto: Morozova.
a b c d e f
RESULTATS DE LA CAMPANYA MICOLÒGICA RUSSO-CATALANA ALS PIRINEUS CATALANS OCCIDENTALS
GEA, FLORA ET FAUNA
bre sòl bàsic, 23 VIII 2014; Davant de Montgarri (8), 23 VIII 2014; Port de la Bonaigua (10, 13), prat alpí, entre gramínies i Calluna vulgaris, sobre sòl àcid, 23 VIII 2014, 24 VIII 2014 (JVG 1140824-16, LE 312451); Bosc del Gerdar de Sorpe (14), clariana amb prat, 24 VIII 2014 (JVG 1140824-25, LE 312452); Núria, vora Santuari (16), prat, sobre sòl àcid, 27 VIII 2014.
*Entoloma cuboidoalbum Noordel. et Hauskn. - Val de Margalida, Bossòst, 1200 m, entre l’herba, sota Abies alba i Betula pendula, 5 IX 2007 (JVG 1070905-3).
Entoloma exile (Fr.) Hesler - Plaça dels Arbres (15), clariana en bosc mixt d’Abies alba i Betula pendula, 25 VIII 2014 (JVG 1140825-2, LE 312434; JVG 1140825-6, LE 312435) (Fig. 3f).
Entoloma formosum (Fr.) Noordel. - Circ dels Estanyets (11), prat, sobre sòl bàsic, 24 VIII 2014 (JVG 1140824-10, LE 312437); Port de la Bonaigua (13), prat alpí, entre gramínies i Calluna vulgaris, sobre sòl àcid, 24 VIII 2014 (JVG 1140824-20, LE 312478).
Entoloma fridolfingense Noordel. et Lohmeyer - Port del Cantó (2), prat, sobre sòl bàsic, 22 VIII 2014 (1140822-1, LE 312436); *Sant Martí Sacalm, Coll de Malla, 980 m, prat sec bàsic, entre plantes rases i Euphrasia, 16 IX 2014 (JVG 1140916-9, JVG 1140916-10) (Fig. 4a).
Entoloma glaucobasis Huijsman ex Noordel. - Vora borda de la Perosa (4), prat subalpí, sobre sòl bàsic, 23 VIII 2014 (JVG 1140823-3, LE 312438); entrant a la Vall d’Aran, primers prats amb Gentiana lutea (7), 23 VIII 2014; davant de Montgarri (8), prat alpí, sobre sòl bàsic, 23 VIII 2014 (JVG 1140823-13, LE 312439); *Toses, Collada del Pedró, 2070 m, en un prat bàsic, entre l’herba, 5 VIII 2014 (JVG 1140805-3); *Toses, Coll de la Creueta, 2090 m, en un prat, entre l’herba, 5 VIII 2014 (JVG 1140805-23, JVG 1140805-24) (Fig. 4b); *Sant Martí Sacalm, el Far, 980 m, en una clariana herbosa, vora Quercus humilis, sòl bàsic, 6 VI 2014 (JVG 1140606-7). Comentaris: relativament freqüent en prats, sobre sòl bàsic, de l’estatge montà a l’alpí. Es separa d’Entoloma asprellum per l’aspecte més tomentós-pruïnós del píleu i per l’estípit densament fibriŀlós. Entoloma glaucodubium Corriol - Pardines (23), bosc de Corylus avellana, Buxus sempervirens, Fraxinus excelsior i Sorbus aria, sobre sòl bàsic, 29 VIII 2014 (JVG 1140829-7, JVG 1140829-10) (Fig. 4c). Comentaris: descrita recentment (Corriol, 2016), sembla estrictament associada a boscos de Corylus avellana, un fet inusual entre les espècies d’Entoloma del subgènere Cyanula. A part de l’hàbitat, s’identifica per l’aspecte del píleu, que presenta un disc central de color rosat, bru rosat o bru vermellós, sobre un fons molt més pàŀid. Nou per Catalunya.
Entoloma griseocyaneum (Fr.) P. Kumm. - Vora borda de la Perosa (4), prat subalpí, sobre sòl bàsic, 23 VIII 2014 (JVG 1140823-5, LE 312440); entrant a la Vall d’Aran, primers prats amb Gentiana lutea (7), 23 VIII 2014; Port de la Bonaigua (10, 13), prat alpí, entre gramínies i Calluna vulgaris, sobre sòl àcid, 23 VIII 2014, 24 VIII 2014; Plaça dels Arbres (15), clariana en bosc mixt d’Abies alba i Betula pendula, 25 VIII 2014; Núria, principi de la Coma de l’Embut (16a), prat bàsic, entre l’herba, 27 VIII 2014.
Entoloma incanum (Fr.) Hesler - Port del Cantó (2), sota Pinus sylvestris, sobre sòl bàsic, 22 VIII 2014; Circ dels Estanyets (11), prat, sobre sòl bàsic, 24 VIII 2014; Núria, principi de la Coma de l’Embut (16a), prat bàsic, entre l’herba, 27 VIII 2014. Comentaris: fàcil d’identificar pel vistós color verd de l’estípit i per la olor, que recorda la del formatge blau. A Catalunya l’hem observat desde l’estatge alpí (exemplars de mida reduïda) fins a la zona mediterrània.
Entoloma infula (Fr.) Noordel. - Port de la Bonaigua (10, 13), prat alpí, entre gramínies i Calluna vulgaris, sobre sòl àcid, 23 VIII 2014, 24 VIII 2014; Núria, vora Santuari (16), prat, sobre sòl àcid, 27 VIII 2014.
Entoloma juncinum (Kühner et Romagn.) Noordel. - Plaça dels Arbres (15), clariana en el bosc mixt d’Abies alba i Betula pendula, 25 VIII 2014 (JVG 1140825-17, LE 312442).
*Entoloma linkii (Fr.) Noordel. - Davant de Montgarri, alt. 1650 m, sobre fusta en descomposició d’Abies alba, 4 IX 2007 (JVG 1070904-10).
Entoloma mediterraneense Noordel. et Hauskn. - Pardines (23), bosc de Corylus avellana, Buxus sempervirens, Fraxinus excelsior i Sorbus aria, sobre sòl bàsic, 29 VIII 2014 (JVG 1140829-8). Comentaris: abundant en tot tipus d’hàbitats mediterranis (boscos de Pinus halepensis, Quercus ilex, brolles de Q. coccifera, Cistus, etc.), fructifica de forma molt més esporàdica en hàbitats més humits, com boscos de Corylus o prats montans. Genèticament relacionat amb Entoloma glaucodubium (Dima et al., 2022, en premsa).
*Entoloma minutigranulosum O.V. Morozova, Noordel., Brandrud et Dima - Toses, Collada del Pedró, 2070 m, en un prat bàsic, entre l’herba, 5 VIII 2014 (JVG 1140805-5) (Fig. 4d); Sant Martí Sacalm, el Far, 980 m, en una clariana, vora Quercus humilis, sòl bàsic, 6 VI 2014 (JVG 1140606-3). Comentaris: l’epitet específic fa referència a l’aspecte finament granulós del píleu, un caràcter diferencial d’aquesta espècie. De recent descripció (Dima et al., 2021), és una espècie no citada anteriorment a Catalunya.
*Entoloma montanum Noordel., J.B. Jordal, Lorås, Eidissen, E. Larss. et Dima - Núria, Queralbs, Ras de l’Ortigar, 2220 m, entre Dryas octopetala i Salix retusa, 27 VIII 1997 (JVG 970827-14) (Fig. 4e). Comentaris: nou per Catalunya.
Entoloma mougeotii (Fr.) Hesler - Pla de Beret (9), sobre molses, a la riba d’un rierol, 23 VIII 2014 (JVG 1140823-23, LE 312445); Plaça dels Arbres (15), clarianes en bosc mixt d’Abies alba i Betula pendula), 25 VIII 2014 (JVG 114082516, LE 312446); *les Planes d’Hostoles (abans de Coll de Condreu), 870 m, en un prat bàsic, entre l’herba, 6 VI 2014 (JVG 1140606-1); *Vidrà, Collfred, 1320 m, en un prat bàsic, entre l’herba, 25 IX 2013 (JVG 1130925-14).
Entoloma muscoalpinum Vila, F. Caball. et Català - Núria (16), prat vora el santuari, 2030 m, entre l’herba, 27 VIII 2014 (JVG 1140827-1) (Fig. 5). Comentaris: descrit a Vila et al. (2014) a partir de diverses mostres d’exemplars joves trobades també a l’àrea de Núria, tot i que a més altitud (2220 m). La present coŀlecció mostra exemplars perfectament desenvolupats. Entoloma korhonenii presenta basidiomes més foscos i amb un característic aspecte fibriŀlós-subesquamós. Un article sobre aquest grup està en fase de redacció.
180 Butlletí ICHN 85 (4), 2021
J. VILA ET AL.






Entoloma poliopus (Romagn.) Noordel. - Pardines (23), bosc de Corylus avellana, Buxus sempervirens, Fraxinus excelsior i Sorbus aria, sobre sòl bàsic, 29 VIII 2014 (JVG 1140829-5). Comentaris: fructifica en boscos o brolles, desde l’estatge subalpí a la zona mediterrània. No l’hem observat en prats. Els exemplars més típics tenen el píleu grisenc i l’estípit blavós, però es tracta d’un tàxon molt variable, no sempre fàcil d’identificar.
Entoloma porphyrogriseum Noordel. - Vora borda de la Perosa (4), prat subalpí, sobre sòl bàsic, 23 VIII 2014 (JVG 1140823-1, LE 312506); Circ dels Estanyets (11), prat, sobre sòl bàsic, 24 VIII 2014 (JVG 1140824-12); Plaça dels Arbres (15), clariana en el bosc mixt d’Abies alba i Betula pendula, 25 VIII 2014 (JVG 1140825-1, LE 312432; JVG 114082511, LE 312433) (Fig. 4f); Pardines (19), prat, 29 VIII 2014; *Toses, vora Collada de Toses, 1900 m, prat bàsic, entre l’herba, 8 VII 2014 (JVG 1140708-23); *Toses, Coll de la Creueta, 2090
5 VIII 2014 (JVG
181 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA
m, en un prat, entre l’herba,
b
d
f
Figura 4. a) Entoloma fridolfingense (JVG 1140916-10) Foto: Vila; b) Entoloma glaucobasis (JVG 1140805-23) Foto: Vila; c) Entoloma glaucodubium (JVG 1140829-7) Foto: Vila; d) Entoloma minutigranulosum (JVG 1140805-5) Foto: Vila; e) Entoloma montanum (JVG 970827-14) Foto: Vila; f) Entoloma porphyrogriseum (LE 312433) Foto: Morozova.
a
c
e
PRINCIPALS RESULTATS DE LA CAMPANYA MICOLÒGICA RUSSO-CATALANA ALS PIRINEUS CATALANS OCCIDENTALS


1140805-25). Comentaris: freqüent en prats alpins i subalpins, no sembla estar present en zones més baixes. Té el píleu densament fibriŀlós i de color molt fosc, a vegades gairebé negre, que contrasta molt amb el color blanquinós de les làmines joves.
Entoloma querquedula (Romagn.) Noordel. - Circ dels Estanyets (11), prat, sobre sòl bàsic, 24 VIII 2014 (JVG 1140824-5, LE 312427; JVG 1140824-17, LE 312510). Comentaris: forma part del difícil grup d’Entoloma serrulatum, amb diverses espècies que poden tenir reflexos olivacis en el píleu (Dima et al., 2022, en premsa).
Entoloma sericellum (Fr.) P. Kumm. sensu lato - Port de la Bonaigua (10, 13), prat alpí, entre gramínies i Calluna vulgaris, sobre sòl àcid, 23 VIII 2014, 24 VIII 2014; (JVG 1140824-22, LE 312459); Plaça dels Arbres (15), clariana en el bosc mixt d’Abies alba i Betula pendula, 25 VIII 2014; Núria, vora Santuari (16), prat, sobre sòl àcid, 27 VIII 2014 (JVG 1140827-4, LE 312460).
Entoloma sericeum Quél. - Port del Cantó (2), al prat, al marge i clarianes del bosc de Pinus sylvestris, sobre sòl bàsic, 22 VIII 2014; vora Pont de Marimanya (7), prat subalpí, sobre sòl bàsic, 23 VIII 2014; davant de Montgarri (8), prat alpí, sobre sòl bàsic, 23 VIII 2014 (JVG 1140823-12, LE312457); Pla de Beret (9), sobre molses, a la riba d’un rierol, 23 VIII 2014; Port de la Bonaigua (10, 13), prat alpí, entre gramínies i Calluna vulgaris, sobre sòl àcid, 23 VIII 2014, 24 VIII 2014;
Circ dels Estanyets (11), prat, sobre sòl bàsic, 24 VIII 2014 (JVG 1140824-14, LE 312458); Núria, vora Santuari (16), prat, sobre sòl àcid, 27 VIII 2014. Comentaris: sens dubte la Nolanea més abundant en prats montans, subalpins o alpins (excepte comunitats de Salix o Dryas, on hi fructifica Entoloma sericeoalpinum, vegeu Vila et al., 2021), tant sobre sòl àcid com bàsic. En l’àrea mediterrània es veu reemplaçada per altres espècies com Entoloma juncinum, E. minutisporum o, fins i tot, E. llimonae (Vila et al., 2013)
Entoloma serrulatum (Fr.) Hesler - Port de la Bonaigua (10, 13), prat alpí, entre gramínies i Calluna vulgaris, sobre sòl àcid, 23 VIII 2014, 24 VIII 2014; Circ dels Estanyets (11a), prat alpí, sobre sòl bàsic, entre Dryas octopetala, 24
VIII 2014 (JVG 1140824-3, LE312461; JVG 1140824-13, LE 312462); Núria, vora Santuari (16), prat sobre sòl àcid, 27
VIII 2014 (JVG 1140827-5, LE 312463).
Entoloma turci (Bres.) M.M. Moser - Port del Cantó (2), prat en clariana vora Pinus sylvestris, sobre sòl bàsic, 22
VIII 2014 (JVG 1140822-3, LE 312464; JVG 1140822-7, LE 312465; JVG 1140822-8, LE 312466; JVG 1140822-10, LE 312467) (Fig. 6a); *Sant Martí Sacalm, el Far, 980 m, en una clariana, vora Quercus humilis, sòl bàsic, 6 VI 2014 (JVG 1140606-9); *El Brull, Pla de la Calma, prat àcid amb Erica scoparia, Calluna i molses rases, 7 X 2014 (JVG 114100725, JVG 1141007-27).
Entoloma undatum (Gillet) M.M. Moser sensu lato - Davant de Montgarri (8), prat alpí, sobre sòl bàsic, 23 VIII 2014; Pla de Beret (9), sobre molses, a la riba d’un rierol, 23 VIII 2014; sobre Santa Maria (12), bosc a la riba d’un rierol, 24 VIII 2014 (JVG 1140824-15, LE 312468); Pardines, vora Coll del Bac (21), en el prat, 29 VIII 2014.
Entoloma viiduense Noordel. et Liiv - Port de la Bonaigua (13), prat alpí, entre gramínies i Calluna vulgaris, sobre sòl àcid, 24 VIII 2014 (JVG 1140824-19); *El Brull, Pla de la Calma, prat àcid amb Erica scoparia i Pteridium, 26 IX 2014 (JVG 1140926-30, JVG 1140926-31, JVG 1140926-32, JVG 1140926-34) (Fig. 6b).
Entoloma violaceoserrulatum Noordel. - Núria, principi Coma de l’Embut (16a), prat bàsic, entre l’herba, 27 VIII 2014 (JVG 1140827-24, LE 312469).
Entoloma weholtii Noordel. - Vora borda de la Perosa (4), prat subalpí, sobre sòl bàsic, 23 VIII 2014 (JVG 1140823-8, LE 312470); Pla de Beret (9), sobre molses, a la riba d’un rierol, 23 VIII 2014 (JVG 1140823-16, LE 312471); Plaça dels Arbres (15), clariana en el bosc mixt d’Abies alba i Betula pendula, 25 VIII 2014 (JVG 1140825-7, LE 312511); Núria, vora Santuari (16) prat, sobre sòl àcid, 27 VIII 2014 (JVG 1140827-11, LE 295070); *Alp, Baga de Saltèguet, 1800 m, prat bàsic, entre l’herba, 8 VII 2014 (JVG 1140708-12).
Rhodophana nitellina (Fr.) Papetti - Pardines (23), en poblament dens de Buxus sempervirens, 29 VIII 2014.
Hygrophoraceae
Cuphophyllus fornicatus (Fr.) Lodge, Padamsee et VizziniPardines (19), prat, 29 VIII 2014.
Cuphophyllus pratensis (Fr.) Bon - Circ dels Estanyets (11), prat, sobre sòl bàsic, 24 VIII 2014; Queralbs, Núria, vora
182 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA
J. VILA ET AL.
Figura 5. a) Locis tipus d’Entoloma muscoalpinum, Ras de l’Ortigar (Núria) Foto: Vila; b) Entoloma muscoalpinum (JVG 1140827-1) Foto: Vila.
a b





Santuari (16), prat, sobre sòl àcid, 27 VIII 2014; Pardines (19), prat, 29 VIII 2014.
Cuphophyllus virgineus (Wulfen) Kovalenko - Vora Pont de Marimanya (7), prat supalpí, sobre sòl bàsic, 23 VIII 2014; Davant de Montgarri (8), prat alpí, sobre sòl bàsic, 23 VIII 2014; Pla de Beret (9), sobre molses, a la riba d’un rierol, 23 VIII 2014; Núria, vora Santuari (16), prat, sobre sòl àcid, 27 VIII 2014; Pardines (20), Buxus sempervirens matollar dens, 29 VIII 2014.
Gliophorus irrigatus (Pers.) A.M. Ainsw. et P.M. Kirk - Pardines, prop Coll del Bac (21), Buxus sempervirens matollar dens, 29 VIII 2014.
Gliophorus psittacinus (Schaeff.) Herink - Núria, principi Coma de l’Embut (16a), prat bàsic, entre l’herba, 27 VIII 2014; Pardines (20), Buxus sempervirens matollar dens, 29 VIII 2014.
Hygrocybe conica (Schaeff.) P. Kumm. - Port del Cantó (2), prat, sobre sòl bàsic, 22 VIII 2014; vora borda de la Perosa (4), prat subalpí, sobre sòl bàsic, 23 VIII 2014; Port de la Bonaigua (10), prat alpí, entre gramínies i Calluna vulgaris, sobre sòl àcid, 23 VIII 2014; Plaça dels Arbres (15), clariana en el bosc mixt d’Abies alba i Betula pendula, 25 VIII 2014; Núria, vora Santuari (16), prat, sobre sòl àcid, 27 VIII 2014.
183 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA
Figura 6. a) Entoloma turci (LE 312465) Foto: Morozova; b) Entoloma viiduense (JVG 1140926-31) Foto: Vila; c) Geoglossum fallax (LE304016) Foto: Fedosova; d) Helvella fibrosa (LE 304009) Foto: Fedosova; e) Hygrocybe turunda (LE 295071) Foto: Morozova.
OCCIDENTALS a b d c e
PRINCIPALS RESULTATS DE LA CAMPANYA MICOLÒGICA RUSSO-CATALANA ALS PIRINEUS CATALANS
GEA, FLORA ET FAUNA
Hygrocybe turunda (Fr.) P. Karst. - Port de la Bonaigua (13), prat alpí, entre gramínies i Calluna vulgaris, sobre sòl àcid, 24 VIII 2014 (JVG 1140824-18, LE 295071) (Fig. 6e); Plaça dels Arbres (15), clariana en el bosc mixt d’Abies alba i Betula pendula, 25 VIII 2014 (JVG 1140825-13, LE 295072).
Hymenogastraceae
Meottomyces dissimulans (Berk. et Broome) Vizzini - Pardines, vora Coll del Bac (20), en una comunitat densa de Buxus sempervirens, 29 VIII 2014. Comentaris: a la localitat estudiada és una espècie molt abundant, que hem observat durant força anys seguits, tant a la primavera (maig) com a l’estiu o principis de la tardor.
Lyophyllaceae
Leucocybe connata (Schumach.) Vizzini, P. Alvarado, G. Moreno et Consiglio - Núria, vora Santuari (16), prat sobre sòl àcid, 27 VIII 2014 (JVG 1140827-26).
Rugosomyces carneus (Bull.) Bon - Circ dels Estanyets (11, 11a), prat sobre sòl bàsic, entre Dryas octopetala, 24 VIII 2014 (JVG 1140824-8, LE 295073); Núria, vora Santuari (16), prat, entre gramínies, sobre sòl bàsic, 27 VIII 2014.
Marasmiaceae
Crinipellis scabella (Alb. et Schwein.) Murrill - Port del Cantó (2), prat, sobre sòl bàsic, 22 VIII 2014 (LE 295074); pista a Montgarri (3), prat amb Eryngium bourgatii, Trifolium i algunes Betula, sobre roca calcària, 24 VIII 2014.
Physalacriaceae
Rhizomarasmius epidryas (Kühner ex A. Ronikier) A. Ronikier et Ronikier - Circ dels Estanyets (11a), sobre Dryas octopetala, 24 VIII 2014 (LE295075).
Tricholomataceae
Dermoloma cuneifolium (Fr.) Singer ex Bon - Pardines (19), prat, 29 VIII 2014.
Melanoleuca brevipes (Bull.) Pat. - Núria, vora Santuari (16), prat sobre sòl àcid, 27 VIII 2014.
Ripartites tricholoma (Alb. et Schwein.) P. Karst. - Pardines (21), en formació densa de Buxus sempervirens, 29 VIII 2014.
Strophariaceae
Panaeolus fimicola (Pers.) Gillet - Port de la Bonaigua (10), prat alpí, entre gramínies i Calluna vulgaris, sobre sòl àcid, 23 VIII 2014; Plaça dels Arbres (15), clariana en el bosc mixt d’Abies alba i Betula pendula, 25 VIII 2014; Núria, vora Santuari (16), prat pasturat, sobre sòl àcid, 27 VIII 2014.
Panaeolus semiovatus (Sowerby) S. Lundell et Nannf. - Núria, vora Santuari (16), prat pasturat, sobre sòl àcid, 27 VIII 2014.
Boletales
Boletaceae
Chalciporus amarellus (Quél.) Bataille - Port del Cantó (2), sota Pinus sylvestris, sobre sòl bàsic, 22 VIII 2014 (LE 312441).
Geastrales
Geastraceae
Geastrum campestre Morgan - Pista a Montgarri (3), Prat amb Eryngium bourgatii, Trifolium i algunes Betula, sobre roca calcària, 23 VIII 2014.
Geastrum fimbriatum Fr. - Port del Cantó (2), prat prop d’un bosc de Pinus sylvestris, roques calcàries o gresos vermells, 22 VIII 2014.
Geastrum triplex Jungh. - Pardines, prop del Coll del Bac (20, 21), matollar dens de Buxus sempervirens 29 VIII 2014.
Conclusions
Presentem un catàleg amb 226 tàxons dels quals, 81, han estat confirmats mitjançant estudi microscòpic o anàlisi genètica (ITS).
Un dels grups més comuns de fongs que es troben als prats és el gènere Entoloma. Un nombre significatiu d’espècies d’aquest gènere s’havien esmentat o descrit en publicacions anteriors (Vila et al., 1997, 1998, 2001, 2006, 2013, 2014; Vila & Caballero, 2007, 2009; Caballero & Vila, 2013).
Un factor important que pot explicar l’alta diversitat d’aquest gènere és la presència de roca calcària a la majoria de les localitats que vam visitar. Es van identificar un total de 38 espècies d’Entoloma, 26 de les quals pertanyen al subgènere Cyanula
Entoloma azureopallidum, E. glaucodubium, E. minutigranulosum i E. montanum no havien estat citats prèviament a Catalunya.
La majoria de les espècies identificades creixien en comunitats de prats de muntanya, rarament en terres baixes, principalment sobre roca calcària. Entre aquestes espècies hi ha Entoloma catalaunicum, que hem trobat a prop de la localitat del tipus. Entoloma fridolfingense, E. glaucobasis, E. griseocyaneum, E. incanum, E. porphyrogriseum, E. querquedula, E. violaceoserrulatum, E. weholtii, etc. també havien estat trobats durant les prospeccions en hàbitats calcaris de la terra baixa, a la Rússia europea i a Europa del Nord (Brandrud et al., 2017; Morozova et al., 2015, 2016).
Entoloma anatinum, E. chalybaeum, E. exile, E. formosum, E. sericellum, E. sericeum, E. turci, E. undatum també poden créixer sobre sòls àcids. Representants del subgènere Nolanea, com Entoloma juncinum i del subgènere Rhodopolia, es troben principalment als boscos.
Al mateix temps, van ser retrobades quatre espècies descrites recentment dels Pirineus: Entoloma aranense (d’Era Val d’Aran), E. muscoalpinum (Vall de Núria) i E. glaucodubium i E. azureopallidum (del vessant francés dels Pirineus). Entoloma aranense i E. azureopallidum també les hem trobat al Nord-Oest del Caucas (dades no publicades).
També hem comprovat la presència d’espècies rares o interessants d’altres grups taxonòmics: Bryoglossum gracile, Hodophilus foetens, Hygrocybe turunda, Rhizomarasmius epidryas, Rhodophana nitellina.
184 Butlletí ICHN 85 (4), 2021
J. VILA ET AL.
Agraïments
Agraïm els recursos rebuts de l’IEC (Programa Biodiversitat dels Fongs i Líquens dels PPCC), i els dels Fons de Recerca Salvador Llimona i Ramón Vila. L’estudi d’Olga Morozova és inclòs en el marc d’un dels projectes de recerca (no. ААААА19-119020890079-6) del Komarov Botanical Institute de la Russian Academy of Sciences, amb un ajut de la Russian Foundation for Basic Research (project No20-04-00349).
Bibliografia
Ballarà, J. 1995. Alguns ascomicets interessants dels Pirineus catalans. Revista Catalana de Micologia, 18: 1-8.
Ballarà, J. 1996. Estudis sobre Cortinarius subalpins de Catalunya: 1. Espècies recoŀlectades a l’Hylocomio-Pinetum catalaunicae. Revista Catalana de Micologia, 19: 47-66.
Ballarà, J. 1997. Nou estudi d’espècies fúngiques interessants dels estatges alpí i subalpí dels Pirineus catalans. Revista Catalana de Micologia, 20: 1-24.
Ballarà, J. 1998. Alguns Cortinarius interessants dels Pirineus catalans. Revista Catalana de Micologia, 21: 137-150.
Ballarà, J., & Escànez, Ll. 1999. Nou estudi de fongs alpins interessants dels Pirineus catalans. Revista Catalana de Micologia, 22: 39-46.
Ballarà, J. 1999. Alguns Cortinarius interessants dels Pirineus catalans. II. Revista Catalana de Micologia, 22: 47-70.
Bon, M., & Ballarà, J. 1995. Aportació a l’estudi de la micoflora alpina dela Pirineus (1ª part). Revista Catalana de Micologia, 18: 39-50.
Bon, M., & Ballarà, J. 1996. Aportació a l’estudi de la micoflora alpina dels Pirineus (2ª part). Revista Catalana de Micologia, 19: 139-156.
Brandrud, T. E., Bendiksen, E., Noordeloos, M. E., Dima, B., & Morozova, O. 2017. Entoloma-arter funna i Jämtland och Medelpad. Svensk Mykologisk Tidskrift, 38 (3): 25-35.
Caballero, F., & Vila, J. 2013. Entoloma nuevos o interesantes de la Península Ibérica (3). Adiciones y correcciones. Fungi non Delineati LXVI (Studies on Entoloma): 63-85, 136-145 (iconografía). Edizioni Candusso, Alassio.
Codina, J., & Font i Quer, P. 1931. Introducció a l’estudi dels macromicets de Catalunya. Cavanillesia, 3: 100-189.
Corriol, G. 2016. Algunos Entoloma inéditos del subgénero Leptonia Errotari, 13: 33-50.
Crous, P. W., Cowan, D. A., Maggs-Kölling, G., Yilmaz, N., Thangavel, R., Wingfield, M. J., Noordeloos, M. E., Dima, B., Brandrud, T. E., Jansen, G. M., Morozova, O. V., Vila, J., Shivas, R. G., Tan, Y. P., Bishop-Hurley, S., Lacey, E., Marney, T. S., Larsson, E., Le Floch, G., Lombard, L., Nodet, P., Hubka, V., Alvarado, P., […], Romero, M., Rooney-Latham, S., Roux, J., Sandoval-Denis, M., Smith, M. T. H., Steinrucken, T. V., Svetasheva, T. Y., Tkalčec, Z., Van Der Linde, E. J., V.D. Vegte, M., Vauras, J., Verbeken, A., Visagie, C. M., Vitelli, J. S., Volobuev, S. V., Weill, A., Wrzosek, M., Zmitrovich, I. V., Zvyagina, E. A., & Groenewald, J. 2021. Fungal Planet description sheets: 11821283. Persoonia, 46: 313-528.
Dima, B., Brandrud, T. E., Corriol, G., Jansen, G. M., Jordal, J. B., Khalid, A. N., Larsson, E., Lorås, J., Morozova, O. V., Naseer, A., Noordeloos, M. E., Rossi, W., Santamaria, S., Sarwar, S., Sesli, E., Usman, M., Afshan, N. S., Ahmad, I., Banerjee, A., Banerjee, K., Bendiksen, E., Rodrigues Da Silva Colombo, D., De Kesel,
A., Dovana, F., Ferisin, G., Hussain, S., Islam, S., Jesus, A. L., Kaygusuz, O., Krisai-Greilhuber, I., Mahammad, S., Mishra, D. K., Nath, P. S., De Oliveira Da Paixã, S. C., Panja, B., Papp, V., Amorim Pires-Zottarelli, C. L., Radnóti, A., Rana, D., Saha, R., Türkekul, I., & Haelewaters, D. 2021. Fungal Systematics and Evolution: FUSE 7. Sydowia, 73: 271-340.
Dima, B., Morozova, O. V., Vila, J., Brandrud, T. E., Jordal, J. B., Bendiksen, E., Lorås, J., Eidissen, S. E., Jansen, G. M., Corriol, G., Krisai-Greilhuber, I., Nuytinck, J., Reschke, K., Weholt, Ø., Biketova, A., Nagy, L. G., Larsson, E., Papp, V., Kovács, G. M., & Noordeloos, M. E. 2022. An ITS/LSU phylogeny of the European species of Entoloma subgenus Cyanula in global perspective. Persoonia (en premsa).
Esteve-Raventós, F. 1978. Estudios sobre el genero Inocybe (Agaricales) en la Península Ibérica e Islas Baleares IV: nueva aportación al estudio del material recogido por R. Singer en Cataluña. Lazaroa, 19: 29-42.
Esteve-Raventós, F., & Vila, J. 1997. Algunos Inocybe de la zona alpina de los Pirineos de Catalunya. I. Revista Catalana de Micologia, 20: 177-186.
Esteve-Raventós, F., & Vila, J. 1998. Algunos Inocybe de la zona alpina de los Pirineos de Catalunya. II. Revista Catalana de Micologia, 21: 185-201.
Esteve-Raventós, F., Vila, J., & Llistosella, J. 1999. Algunos Entoloma interesantes o poco conocidos de los Pirineos de Catalunya. Revista Catalana de Micologia, 22: 135-143.
Esteve-Raventós, F., Vila, J., & Llistosella, J. 2001. Algunos hongos interesantes del piso subalpino de los Pirineos de Cataluña. Revista Catalana de Micologia, 23: 79-90.
Font i Quer, P. 1937. Resultats del Pla Quinquennal Micològic a Catalunya 1931-1935. Arxius de l’Escola Superior d’Agricultura de Barcelona nova ser., 3: 523-536.
Galán, R., Raitviir, A., & Vila, J. 1997. On some noteworthy alpine or subalpine Hyaloscyphaceae (Leotiales) found in the Pyrenees. Revista Catalana de Micologia, 20: 137-146.
Gorris, M., Gràcia, E., Vila, J., & Llimona, X. 1999. Alguns mixomicets, principalment quionòfils, nous o poc citats als Pirineus catalans. Revista Catalana de Micologia, 22: 23-34.
Knudsen, H., & Vesterholt, J. (Eds.) 2012. Funga Nordica. Agaricoid, boletoid, clavarioid, cyphelloid and gastroid genera Copenhagen: Nordsvamp. 1083 p.
Llimona, X. 1997. Rolf Singer, 1906-1994; nota sobre la seva vida, amb especial atenció a la seva activitat a Barcelona. Collectanea Botanica, 23: 163-169.
Llimona, X. 2009. La Criptogàmia i la Reial Acadèmia: La recerca en Micologia (Fongs i Líquens). Memòria de la Reial Acadèmia de Ciències i Arts de Barcelona, 1021. Vol. LXIV núm. 4. Barcelona 94 p.
Llistosella, J., Martí, O., Aguasca, M., & Martín, M. P. 1994. Aportació a la flora micològica del Parc Nacional d’Ordesa i Monte Perdido (Pirineus centrals). Butlletí Societat Catalana de Micologia, 16-17: 99-118.
Llistosella, J., Aguasca, M., & Vila, J. 1996. Basidiomicets de les torberes dels Pirineus catalans. Primera contribució. Revista Catalana de Micologia, 19:67-72.
Llistosella, J., Vila, J., Rocabruna, A., Tabarés, M., & Llimona, X. 1998. Alguns fongs interessants del Parc Nacional d’Aigüestortes i Estany de Sant Maurici. Revista Catalana de Micologia, 21: 151-157.
Llorens-van-Waveren, L., & Llistosella, J. 2004. Contribució a la flora dels fongs del Parc Natural del Cadí-Moixeró (Catalunya). I. El gènere Entoloma (Fr.) P. Kumm. Revista Catalana de Micologia, 26: 165-176.
185 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA
PRINCIPALS RESULTATS DE LA CAMPANYA MICOLÒGICA RUSSO-CATALANA ALS PIRINEUS CATALANS OCCIDENTALS
GEA, FLORA ET FAUNA
Llorens-van-Waveren, L., & Llistosella, J. 2005. Fongs de Gréixer. Revista Catalana de Micologia, 27: 131-151.
Llorens-van-Waveren, L., & Llistosella, J. 2006. El gènere Entoloma a Catalunya. I. Notes florístiques. Revista Catalana de Micologia, 28: 87-91.
Morozova, O.V., Malysheva, E. F., Popov, E. S., Psurtseva, N. V., Fedosova, A. G., & Kovalenko, A. E. 2015. Macromycetes of the Izborsk-Maly Valley, rare and new to the Pskov Region. Novosti Sistematiki Nizaikh Rastenii, 49: 186-203.
Morozova, O. V., Alexandrova, A. V., Popov, E. S., & Malysheva, E. F. 2016. New data on the agaricoid basidiomycetes of the Tver Region. Novosti Sistematiki Nizaikh Rastenii, 50: 174-186.
Morozova, O. V., Noordeloos, M. E., & Vila, J. 2014. Entoloma subgenus Leptonia in boreal-temperate Eurasia: Towards a phylogenetic species concept. Persoonia, 32 (1): 141-169.
Muntañola-Cvetkovic, M., Hoyo, P., & Llimona, X. 1997. Fongs mitospòrics dels estatges montà, subalpí i alpí de Catalunya. Revista Catalana de Micologia, 20: 199-212.
Noordeloos, M. E. 1992. Entoloma s.l. Fungi Europaei, vol. 5. Giovanna Biella, Saronno, Italy. 760 pp.
Noordeloos, M. E. 2004. Entoloma s.l. Fungi Europaei, vol. 5a. Edizione Candusso, Italy. 618 pp.
Quadrada, R., Llistosella, J., & Vila, J. 1997. Basidiomicets de les torberes dels Pirineus catalans. II. El gènere Galerina. Revista Catalana de Micologia, 20: 213-220.
Rocabruna, A., Tabarés, M., Ballarà, J., & Vila, J. 1994. Primera aportació al coneixement de la micoflora dels Pirineus i PrePirineus de Catalunya. Butlletí Societat Catalana de Micologia, 16-17: 47-86.
Rocabruna, A., Vila, J., Tabarés, M., & Ballarà, J. 1996. Aportación a la micoflora de los Pirineos y Pre-Pirineos de Cataluña. II. Algunas especies asociadas al abeto (Abies alba). Revista Catalana de Micologia, 19: 139-153.
Singer, R. 1935. Étude systématique sur les Melanoleuca d’Europe et clé des espèces observées en Catalogne. Cavanillesia, 7: 122-132.
Singer, R. 1936a. Notes sur quelques Basidiomycètes. Revue Mycologique (Paris), 1(2): 75-84.
Singer, R. 1936b. Notes sur quelques Basidiomycètes. II série. Revue Mycologique (Paris), 1(6): 279-293.
Singer, R. 1936c. Les rússules de Catalunya i clau per a llur determinació. Cavanillesia, 8: 144-159.
Singer, R. 1947. Champignons de la Catalogne observés en 1934 (Suppl. I, Suppl. II). Collectanea Botanica (Barcelona), 1(3): 199-246.
Tabarés, M., Llistosella, J., & Rocabruna, A. 2004. Fongs nous o poc citats de Catalunya. VI. Revista Catalana de Micologia, 26: 43-58.
Vidal, J. M., & Vila, J. 1994. Algunos hongos hipogeos de Andorra. Butlletí Societat Catalana de Micologia, 16-17: 213-220.
Vidal, J. M. 1994. Algunos hongos hipogeos interesantes para la micoflora catalana. Butlletí Societat Catalana de Micologia, 1617: 221-248.
Vidal, J. M. 1997. Algunos hongos hipogeos nuevos o poco citados de Cataluña (Zygomycotina, Ascomycotina y Basidiomycotina). Revista Catalana de Micologia, 20: 25-62.
Vila, J., & Caballero, F. 2007. Entoloma nuevos o interesantes de la Península Ibérica. Fungi non delineati. Pars XXXVIII. Edizione Candusso, Italy. 64 p.
Vila, J., & Caballero, F. 2009. Entoloma nuevos o interesantes de la Península Ibérica (2). Fungi non delineati. Pars XLV. Edizione Candusso, Italy. 100 p.
Vila, J., & Llimona, X. 1999. Els fongs del Parc Natural del Cap de Creus i Serra de Verdera (Girona). II. Aproximació al component fúngic del Cistion. Revista Catalana de Micologia, 22: 95-114.
Vila, J., Caballero, F., Carbó, J., Alvarado, P., Català, S., Higelmo, M.A., & Llimona, X. 2014. Preliminary morphologic and molecular study of the Entoloma rusticoides group (Agaricales-Basidiomycota). Revista Catalana de Micologia , 35: 65-99.
Vila, J., Carbó, J., Caballero, F., Català, S., Llimona, X., & Noordeloos, M. E. 2013. A first approach to the study of the genus Entoloma subgenus Nolanea s.l. using molecular and morphological data. Fungi non delineati. Pars LXVI (studies on Entoloma): 3-62, 93-135 (iconography). Edizioni Candusso, Alassio.
Vila, J., Llistosella, J., & Llimona, X. 1997. Contribució al coneixement dels fongs de l’estatge alpí dels Pirineus de Catalunya. I. Revista Catalana de Micologia, 20: 221-232.
Vila J., Llistosella, J., & Llimona, X. 1998. Contribució al coneixement dels fongs de l’estatge alpí dels Pirineus de Catalunya. II. Revista Catalana de Micologia, 21: 93-113.
Vila, J., Esteve-Raventós, F., Llistosella, J., & Llimona, X. 2001. Contribución al conocimiento de los hongos del piso alpino de los Pirineos de Cataluña. III. Revista Catalana de Micologia, 23: 51-65.
Vila, J., Caballero, F., & Tabarés, M. 2006. Alguns fongs de primavera interessants trobats als Pirineus catalans. Revista Catalana de Micologia, 28: 119-124.
Vila, J., Noordeloos, M. E., Reschke, K., Moreau, P.-A., Battistin, E., Ribes, M. Á., Marulli, U., Corriol, G., Polemis, E., Loizides, M., & Dima, B. 2021. New species of the genus Entoloma (Basidiomycetes, Agaricales) from Southern Europe. Österreichische Zeitschrift für Pilzkunde, 29: 123–153.
186 Butlletí ICHN 85 (4), 2021
J. VILA ET AL.
NOTA BREU
Primeros registros de Nigrotipula nigra nigra (Linnaeus, 1758) para la Península Ibérica (Diptera: Tipulidae)
First records of Nigrotipula nigra nigra (Linnaeus, 1758) (Diptera: Tipulidae) for the Iberian Peninsula
Jorge Mederos*, Adrià Miralles-Núñez** & Eduard Marquès-Mora***
* Museu de Ciències Naturals de Barcelona. Laboratori de Natura. Departament d’artròpodes. Passeig Picasso s/n. 08003 Barcelona.
** Servicios Depec. Responsable d’Entomologia. C/ Nena Casas, 71. 08017 Barcelona.
*** Institució Alt Empordanesa per l’Estudi i Defensa de la Natura (IAEDEN). C/Sant Vicenç, 30, 1r. 17600 Figueres.
Autor per a la correspondencia: Jorge Mederos. A/e: mederos@gmail.com
Rebut: 25.10.2021. Acceptat: 21.11.2021. Publicat: 30.12.2021 Butlletí de
El género Nigrotipula Hutson & Vane-Wright, 1969 está representado por solo cuatro especies: Nigrotipula achlypoda (Alexander, 1966), Nigrotipula bathroxantha (Alexander, 1961) y Nigrotipula xanthocera (Alexander, 1936) que se citan exclusivamente de la India, mientras que Nigrotipula nigra nigra (Linnaeus, 1758) posee una distribución más amplia en la región paleártica, especialmente en el Paleártico occidental (Oosterbroek, 2021). Esta especie se ha registrado en praderas costeras, reportándose de marismas (con larvas en el lodo orgánico más o menos saturado de los márgenes de lagos y pantanos), pastizales húmedos y costeros, así como también asociado a carrizales y hábitats semihúmedos (Kramer & Withers, 2007; Autio & Salmela, 2010; Salmela, 2010; Boardman, 2017; Tillier & Dehalleux, 2019). Oosterbroek (2021) reúne toda la información disponible sobre esta especie en el Catalogue of the Craneflies of the World. En el presente trabajo, se aportan los primeros registros de N. nigra nigra en la Península Ibérica, en la comarca de l’Alt Empordà, en Cataluña.
Durante la segunda quincena de agosto de 2021, el tercer autor fotografió un macho de la familia Tipulidae (Fig. 1) en áreas periféricas al Parc Natural dels Aigüamolls de l’Empordà (Girona). La coloración totalmente negra del espécimen, así como la coloración oscura y venación de las alas (Fig. 2a), no correspondía a ninguna de las especies citadas en la Península Ibérica. Poco después, se visitaron áreas cercanas a la localidad anterior y se observaron y capturaron nuevos ejemplares, confirmándose la identidad de la especie como N. nigra nigra. La observación de la actividad de los especímenes en el área se prolongó hasta la primera mitad de octubre de 2021. Para la identificación del material se consultó el Catalogue of the Craneflies of the World (Oosterbroek, 2021). Los ejemplares fueron capturados mediante manga entomológica y posteriormente conservados en etanol al 70 % o en seco. Los especímenes fueron estudiados con un microscopio estereoscópico Motic SMZ-168, las imá-
genes en laboratorio se tomaron con un microscopio óptico Kyowa Unilux-12 83-483D, con apilado de imágenes mediante cámara iPhone XR y posteriormente procesadas con Helicon Focus 7. El material se encuentra depositado en la colección general del Museu de Ciències Naturals de Barcelona (MCNB), con el número de registro correspondiente antecedido por el acrónimo MZB y que será mencionado al especificar el material examinado.
Familia Tipulidae Latreille, 1802
Subfamilia Tipulinae Latreille, 1802
Nigrotipula nigra nigra (Linnaeus, 1758) (Figs. 1, 2)
Material examinado: 1 ♂, Castelló d’Empúries, Girona, Cataluña, España, 23/08/2021, E. Marquès leg. MZB 20211542 (EtOH 70 %); 2 ♂♂ idem, MZB 2021-1543, MZB 2021-1544 (seco); 1 ♀, Peralada, Girona, Cataluña, España, 25/08/2021, E. Marquès leg. MZB 2021-1545 (seco); 1 ♂ idem MZB 2021-1546 (seco); 1 ♂ 1 ♀ (cópula), Pau, Girona, Cataluña, España, 15/09/2021, E. Marquès leg. MZB 20211828 (EtOH 70 %).
En el caso de los especímenes recolectados en el Empordà, las dos localidades donde se encontraron se ubican en antiguas lagunas de origen aluvial, actualmente dedicadas a la obtención de forraje para ganado y donde la gramínea dominante en estos prados es Paspalum distichum. Los prados están rodeados por unos canales de drenaje donde se desarrolla vegetación dominada por árboles de ribera y una densa vegetación arbustiva. Son prados inundables, ya sea por precipitación o por inundación artificial mediante el bombeo de agua de los canales adyacentes en los períodos estivales. Dada la cercanía de los puntos de muestreo a los límites del Parc Natural dels Aiguamolls de l’Empordà, no sería descartable la presencia de la especie dentro del mismo. Sería de interés una mayor prospección de la zona para establecer la extensión real de la población, ya que se sitúa en una de las
187 Butlletí ICHN 85 (4), 2021
BREU
NOTA
edition)
la Institució Catalana d’Història Natural, 85 (4): 187-190. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print
DOI: 10.2436/20.1502.01.91

áreas más meridionales de su distribución en el Paleártico occidental, que se extiende desde los países del norte de Europa, como Suecia (Salmela, 2010), hasta el sur, como Grecia (Oosterbroek, 2009). Las fechas donde se ha detectado la especie en Cataluña (agosto hasta mediados de octubre) se extiende un poco más del período de vuelo citado de poblaciones en áreas próximas como Francia o Italia (Oosterbroek, 2008; Quindroit, 2020), por lo que es probable que la especie presente una fenología similar a estas regiones. Durante el mes de agosto se observaron casi exclusivamente especímenes macho, y en septiembre se observó la primera cópula de la especie. Con el presente registro se eleva a 7 los géneros de
la familia Tipulidae citados para la Península Ibérica y a 165 el número de especies.
Agradecimientos
Nuestro agradecimiento al equipo de conservadoras y técnicos del Departament d’artròpodes del MCNB por facilitar el estudio de este interesante material. A Eulalia Eiroa (Universidad de Santiago de Compostela) y Miguel Carles-Tolrá por la revisión, observaciones y apoyo brindado durante la redacción del manuscrito.
188 Butlletí ICHN 85 (4), 2021 NOTA BREU J. MEDEROS ET AL.
e
Figura 1. a) Localización de las áreas muestreadas dentro de los términos municipales de la provincia de Girona. b-c) Vistas generales de dos de las áreas en el momento del muestreo. Habitus del macho de Nigrotipula nigra nigra fotografiado en una de las localidades muestreadas: d) en vista dorsal; e) lateral.
a b c d

Bibliografía
Autio, O. & Salmela, J. 2010. The semi-aquatic fly fauna (Diptera) of wetlands of the Aland Islands. Memoranda Societatis pro Fauna et Flora Fennica, 86: 43-53.
Boardman, P. 2017. Cranefly report for Shropshire & elsewhere, 2016. Bulletin of the Dipterists Forum 83; Cranefly News. Dipterists Forum Cranefly Recording Scheme Newsletter, 32: 3-4.
Kramer, J. & Withers, P. 2007. Craneflies (Diptera) from the Pierre Verots Nature Reserve, St Jean de Thurigneux, Ain, France. Dipterists Digest (2nd series), 13: 157-163.
Oosterbroek, P. 2008. Tipulidae. P. 337-344. In: Ziegler, J. (ed.), Diptera Stelviana. Vol. 1. Studia Dipterologica. Supplement 16. Müncheberg, Germany.
Oosterbroek, P. 2009. New distributional records for Palaearctic Limoniidae and Tipulidae (Diptera: Craneflies), mainly from the collection of the Zoological Museum, Amsterdam. In: Lantsov, V. (ed.), Crane flies. History, taxonomy and ecology (Diptera: Tipulidae, Limoniidae, Pediciidae, Trichoceridae, Ptychopteridae, Tanyderidae). Memorial volume dedicated to Dr. Charles Paul Alexander (1889-1981), Dr. Bernhard Mannheims (1909-1971) and Dr. Evgeniy Nikolaevich Savchenko (1909-
189 Butlletí ICHN 85 (4), 2021 NOTA BREU PRIMEROS REGISTROS DE NIGROTIPULA NIGRA NIGRA (LINNAEUS, 1758) PARA LA PENÍNSULA IBÉRICA
a b c d e f
Figura 2. Nigrotipula nigra nigra: a) venación alar; b) ovipositor en vista lateral; c) hipopigio en vista lateral; bomba espermática y edeago: d) en vista lateral; e) dorsal; f) gonostilos externo e interno.
NOTA BREU
1994). Zoosymposia 3: 179-197.
Oosterbroek, P. 2021. Catalogue of the Craneflies of the World (CCW). Disponible en: http://ccw.naturalis.nl [Fecha de consulta: 10 octubre, 2021].
Quindroit, C. 2020. Une premiere liste des Tipuloidea des Pays de la Loire et addition d une espece de Limoniidae a la faune de France (Diptera). L’Entomologiste. 76: 5-48.
Salmela, J. 2010. Cranefly (Diptera, Tipuloidea & Ptychopteridae) fauna of Limhamn limestone quarry (Sweden, Malmo). Diversity and faunistics viewed from a NW European perspective. Norwegian Journal of Entomology, 57: 123-135.
Tillier, P. & Dehalleux, A. 2019. Les Tipulidae d’Ile-de-France: donnees recentes et synthese des connaissances regionales (Diptera: Tipulidae). L’Entomologiste, 75: 143-176.
190 Butlletí ICHN 85 (4), 2021
MEDEROS ET AL.
J.
Sobre la gairebé absència de Rosmarinus officinalis (Lamiaceae) al Pla d’Urgell i algunes anomalies fenològiques interessants
Antoni Mayoral Arqué*
* Secció de Botànica de l’Institut d’Estudis Ilerdencs. Pl. Catedral s/n. 25007 Lleida. A/e: amayoral@xtec.cat
Rebut: 26.10.2021; Acceptat: 28.11.2021; Publicat: 30.12.2021
Resum
S’ha realitzat un estudi fenològic de Rosmarinus officinalis durant prop de set anys (octubre 2014-juny 2021) a la comarca del Pla d’Urgell i altres contrades properes (depressió de l’Ebre, W Catalunya). La floració d’aquesta espècie al llarg dels mesos més freds, la seva localització als vessants S de les zones enlairades dels tossals més elevats i la feble exposició als vents més freds i secs del NW, ens porten a suggerir les temperatures mínimes molt baixes amb fortes gelades que es donen a l’hivern en situacions d’inversió tèrmica, com el principal factor que explicaria la seva absència a les zones més planes i baixes de la resta de la comarca. La segona part d’aquest article tracta de la floració inusual de Stipa offneri i altres espècies, detectada entre novembre de 2014 i juny de 2015, període en què les estacions meteorològiques del Pla d’Urgell també varen enregistrar importants anomalies.
Paraules clau: Plantes vasculars, fenologia, anomalies, meteorologia, depressió de l’Ebre.
Abstract
About the near absence of Rosmarinus officinalis (Lamiaceae) in Pla d’Urgell and some interesting phenological anomalies
A phenological study of Rosmarinus officinalis has been carried out for almost seven years (October 2014-June 2021) in the region of Pla d’Urgell and other nearby areas (Ebro Valley, W Catalonia). The flowering of this species during cold winter months, its location on the S slopes of elevated areas of the highest hills and its little exposure to NW colder and drier winds lead us to suggest that the very low minimum temperatures with severe frosts that occur in winter in situations of thermal inversion are the main factor that would explain its absence in flatter and lower areas of the rest of the region. The second part of this article deals with the unusual flowering of Stipa offneri and other species, detected between November 2014 and June 2015, a period in which the meteorological stations of Pla d’Urgell also recorded significant anomalies.
Key words: Vascular plants, phenology, anomalies, meteorology, Ebro Valley.
Introducció
En la nostra tesi de llicenciatura (Mayoral, 1986) fèiem referència a la quasi absència del romer (Rosmarinus officinalis L.) al Pla d’Urgell. Sembla ser que aquesta gairebé inexistència podria estar relacionada amb el factor continentalitat, manifestat sobretot per la més o menys distància al mar i la inversió tèrmica que es dona a les parts més fondes, amb fortes i freqüents gelades i la formació de boires persistents. Relacionat amb el que acabem d’esmentar, trobem l’estudi (Terradas, 1973, 1986) realitzat en una altra zona de la depressió de l’Ebre (els Monegros), el qual va detectar inversió tèrmica fins i tot en desnivells de només 20 m: generalment es donaven temperatures mínimes més altes al cim dels turons i el nombre d‘hores de gelada era més elevat a les petites valls. En aquest mateix treball, s’esmenta també un altre fet que podria condicionar la distribució de la vegetació: el paper, com a aixopluc de plantes termòfiles, dels rius i barrancs que hi van a parar. En aquests indrets, les turbulències evitarien l’estancament de l’aire fred i serien llocs arrecerats,
on l’acció del cerç seria menys intensa. A la tardor de l’any 2014, tot just quan iniciàvem les primeres observacions del comportament del romer, ens va sorprendre una anomalia fenològica, concretament la floració fora del seu temps habitual d’algunes espècies. Va ser llavors quan vam decidir fer un seguiment conjunt del romer i d’aquestes altres plantes durant un període prou llarg, una mica més de set anys. El treball s’ha localitzat al Pla d’Urgell, per bé que en l’apartat del romer s’han fet algunes observacions en comarques properes, circumstància que ens va permetre constatar una altra anomalia en la vegetació dels guixos de la Serra de BellmuntAlmenara.
Metodologia
Llocs d’estudi
El Pla d’Urgell, des de la construcció del canal homònim, ha experimentat moltes modificacions que han comportat que quedin molts pocs indrets on es desenvolupa la vegetació
191 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA GEA, FLORA ET FAUNA
Butlletí de la Institució Catalana d’Història Natural, 85 (4): 191-198. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition) DOI: 10.2436/20.1502.01.92
adaptada a les condicions climatològiques que s’hi donen. Les observacions i seguiments s’han centrat en zones molt properes a Mollerussa (vegeu fig.1), concretament al Tossal de l’Infern (31TCG2206, 07), que forma part de l’EIN Tossals de Torregrossa i a l’indret conegut com la Serra (31TCG2208, 2108, 2109, 2008, 2009, 1909) que ocupa part dels termes de Sidamon, Torregrossa, Fondarella, Mollerussa i Miralcamp. Aquests tossals, molt lligats a la dinàmica fluvial del Corb (Calvet, 1977), són relleus tabulars que s’orienten en sentit ESE-WNW i estan formats per graves cantelludes plistocèniques dipositades sobre argiles amb intercalacions de gresos del Catià (Oligocè). Sobresurten entre 15 i 90 m de la resta dels indrets i els seus sòls pertanyen al tipus pètric Calcisòl, concretament a la sèrie Seana (Boixadera et al., 2009), els quals estan caracteritzats per un pH que va de 7,9 a 8,3 i per ser molt permeables; l’element més destacat és l’horitzó petrocàlcic Bkm, format per nòduls de graves i sorres molt cimentades per Ca CO3. La vegetació d’aquests indrets (Conesa et al., 1994) pertany al domini climàcic del carrascar (Quercetum rotundifoliae), les poques flotes d’alzina que hi queden en donen testimoni (vegeu fig. 4). La flora és eminentment mediterrània amb alguns components estèpics. La sèrie regressiva del carrascar comporta en primer lloc l’aparició del coscollar o garriga (Quercetum cocciferae) que és substituït per la brolla de romaní i maleïda (Rosmarino officinalis-Linetum suffruticosi), la manca del romer en la majoria dels casos li dona una aspecte més aviat de timoneda i en el seu si hi destaca l’abundància de Stipa offneri. La degradació d’aquesta comunitat comporta l’aparició del llistonar (Ruto angustifoliae-Brachypodietum ramosi), molt estès en els vessants N. La part S d’aquests tossals sol estar ocupada per l’espartar (Lygeo sparti-Stipetum parviflorae), caracteritzat sobretot per Stipa parviflora. Als espais oberts que deixen els llistonars i els espartars s’hi fan, respectivament, les associacions de caire terofític Clypeolo jonthlaspi-Psiluretum incurvi i Vulpio unilateralis-Minuartietum campestris. L’etapa
final de la sèrie correspon a la comunitat nitrohalòfila de siscall i botja pudent (Salsolo vermiculatae-Artemisietum herbae-albae). El clima d’aquestes contrades (Mayoral, 1986) és mediterrani continental, semiàrid, amb amplitud tèrmica accentuada, fortes gelades i freqüents boires persistents. Les precipitacions anuals estan al voltant de 400 mm, la primavera i la tardor solen ser les estacions on s’acumula més pluja. Les mitjanes anuals de T estan al voltant de 14 ºC. Pel que fa als vents (Busqué et al., 1989), el dominant és el seré o ponent (W); altres vents a destacar són el morella o garbí (SW), el llevant (E) que sol portar les pluges, la refrescant marinada (SE) que acostuma a bufar sobretot a l’estiu i el cerç o mestral (NW), molt fred i sec a l’hivern.
Espècies estudiades
Pel que fa al romer, s’ha estudiat la ubicació i la fenologia de l’única població d’aquesta espècie al Pla d’Urgell, situada al vessant S del Tossal de l’Infern (vegeu fig. 5). També s’ha tingut en compte un sol exemplar que es troba a la Serra (al terme de Torregrossa). El seguiment de la floració, tant del romer com de les altres espècies, s’ha realitzat entre l’octubre de 2014 i juny de 2021. En el cas del romer, també s’ha analitzat el comportament del que es localitza al Tossal Gros (terme d’Arbeca), indret amb les mateixes característiques geològiques i pedològiques, però, en aquest cas, forma part d’una brolla més termòfila, el Cytiso fontanesii-Cistetum clusii (=Genisto biflorae-Cistetum clusii). Igualment, s’ha tingut en compte la floració de les poblacions dels Gypsophiletalia de prop de la Sentiu de Sió, amb unes diferències pel que fa al substrat ben clares. Quant a les espècies que van florir de manera anòmala, se n’ha fet un seguiment durant el mateix període de temps a les zones del Tossal de l’Infern i la Serra. Aquestes plantes són les següents: Stipa offneri Breistr., S. parviflora Desf., Thymus vulgaris L. subsp. vulgaris, Genista scorpius (L.) DC., Lithodora fruticosa (L.) Griseb., Heliant-
192 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA A. MAYORAL ARQUÉ
Figura 1. La major part de l’estudi s’ha efectuat en els tossals d’aquesta zona del Pla d’Urgell. Font del mapa: Institut Geològic i Cartogràfic de Catalunya (2021).
SOBRE LA GAIREBÉ ABSÈNCIA DE ROSMARINUS OFFICINALIS (LAMIACEAE) AL PLA D'URGELL
Taula 1. Síntesi dels índexs de floració dels tàxons estudiats al llarg dels anys i mesos del període octubre 2014-juny 2021.

hemum marifolium (L.) Mill. subsp. marifolium, H. violaceum (Cav.) Pers. (=H. pilosum (L.) Pers.), Teucrium capitatum L. subsp. capitatum i T. gnaphalodes L’Hér.
Avaluació de la fenologia de floració
Tant en el romer com les altres espècies, per tal de quantificar la floració, s’ha utilitzat un índex amb dos valors: percentatge d’individus florits de cada espècie i grau de floració individual. El percentatge es valora de la manera següent: - ,
cap individu florit; 1, menys de cinc exemplars florits; 2, cinc o més individus florits, però no s’arriba al 25 %; 3, entre el 25 % i el 50 %; 4, més del 50 % de les mates, florides. El grau de floració individual s’ha establert de la següent manera: 1, cap exemplar arriba a les 10 flors; 2, menys del 25 % dels individus té més de 10 flors; 3, del 25 % al 50 % dels exemplars presenta més de 10 flors; 4, més del 50 % dels individus presenta més de 10 flors. Així, per exemple, una espècie que experimenti un índex 3.2, tindrà entre el 25 i el 50 % de mates florides i menys del 25 % d’elles presentarà més de 10 flors. Com es pot veure, la segona xifra, el grau de floració individual, ens ajuda a matisar millor la intensitat de la floració, sobretot quan aquesta no és massa elevada, com és el cas de les anomalies, a més, en el treball de camp és fàcil d’utilitzar. Es van seleccionar superfícies de 50 m2 on s’havien observat les anomalies fenològiques; per tal d’establir el percentatge de floració, s’hi va comptar el nombre de mates totals i les florides de cada espècie. Es va visitar com a mínim una vegada al mes cada indret i s´han efectuat les mitjanes dels resultats de les observacions mensuals de les diferents poblacions. Quan la mitjana d’alguna espècie no arriba a l’índex de floració mínim -1.1-, es fa la mitjana només amb els valors dels inventaris en què ha experimentat floració i es posa entre parèntesis, així es poden constatar les floracions esporàdiques. La síntesi d’aquests resultats es presenta a la taula 1. En els mesos de juliol, agost i setembre, no es va registrar cap floració dels tàxons estudiats, per aquest motiu no s’han fet constar per tal de no complicar més la taula.
D’altra banda, s’han utilitzat les dades, gràfics i taules disponibles a la xarxa del Servei Meteorològic de Catalunya
193 Butlletí ICHN 85 (4), 2021
FLORA ET FAUNA
GEA,
2014 2015 2016 2017 O N D G F M A M J O N D G F M A M J O N D G F M A M J O N D Stipa offneri - 3.2 3.3 3.3 1.1 - - 2.2 - - - - - - - 2.2 4.4 1.1 - - - - - - 2.2 4.4 1.1 - -Stipa parviflora - 2.2 3.3 2.2 - - - 3.3 2.2 - - - - (1.1) - 2.2 4.4 3.2 - - - - - - 2.2 4.4 3.2 (1.1) -Thymus vulgaris - 2.2 2.2 2.2 3.2 1.1 2.2 3.3 - - - - 1.1 - 2.2 4.3 4.4 1.1 - - - - - 2.2 4.3 4.3 1.1 - -Genista scorpius - 1.1 2.2 1.2 2.2 2.2 3.3 - - - - - (1.1) - 3.3 4.4 - - - - - - - 2.2 4.4 - - - -Helianthemum marifolium - 3.1 1.1 - - - 1.1 2.2 - - - - (1.1) - 1.1 2.2 4.2 2.1 - - - - - 2.1 2.2 4.2 2.1 - -Helianthemum violaceum - 3.1 2.1 1.1 - - - 2.1 - - - - - - 1.1 3.1 4.1 - - - - - - 1.1 3.1 4.1 - - -Lithodora fruticosa - 1.1 1.1 - - 1.1 3.3 2.2 - - - - (1.1) - 3.3 4.4 3.2 - - - - - - 2.1 4.4 3.2 - - -Teucrium capitatum - - 1.2 - - - - 1.1 2.2 - - - - - - - 2.2 4.3 - - - - - - 1.1 2.2 4.3 - -Teucrium gnaphalodes - - 1.3 - - - 1.1 3.3 2.2 - - - - - - 2.2 4.3 3.3 - - - - - - 2.2 4.3 3.3 - -Rosmarinus officinalis - 3.2 3.3 3.4 4.4 4.4 2.2 - - - 2.2 3.3 4.3 4.3 4.4 2.2 - - - 2.2 2.3 2.3 3.4 4.4 2.2 - - - 3.3 3.3 2018 2019 2020 2021 G F M A M J O N D G F M A M J O N D G F M A M J O N D G F M A M J Stipa offneri - (1.1) - 3.2 4.4 1.1 - - - - - (1.1) 2.2 4.4 1.1 - - - - - - 2.2 4.4 1.1 - - - - - - 3.2 4.4 1.1 Stipa parviflora (1.1)(1.1) - 2.2 4.4 3.4 - - - - - - 2.2 4.4 3.2 - - - - - - 2.2 4.4 3.2 - - - - - - 2.2 4.4 3.2 Thymus vulgaris - 1.1 2-2 4.4 4.3 1.1 - - - - - 1.2 4.3 4.4 1.2 - - - - - 3.3 4.3 4.4 1.2 - - - - 1.1 2.2 4.3 4.4 1.1 Genista scorpius - 1.1 1.1 4.4 - - - - - - - 2.2 4.3 - - - - - - - 2.2 4.4 - - - - - - 1.1 2.2 4.4 -Helianthemum marifolium - - 1.1 2.2 4.2 2.1 - - ( 1.1) (1.1) - 2.1 2.2 4.2 2.1 - - - - - - - - - - - - - - 2.1 3.2 4.2 2.1 Helianthemum violaceum - - 1.1 3.1 4.1 - - - (1.1) - - 1.1 3.1 4.1 - - - - - - 1.1 3.1 4.1 - - - - - - 1.1 3.1 4.1Lithodora fruticosa - 2.1 2.1 4.4 3.2 - - - - - - 3.3 4.4 2.2 - - - - - - 2.2 4.4 3.2 - - - - - 1.1 2.2 4.4 3.2Teucrium capitatum - - - 1.1 2.2 4.3 - - - - - - 1.1 2.2 4.3 - - - - - - - 2.2 4.3 - - - - - - 1.1 2.2 4.3 Teucrium gnaphalodes - - - 3.3 4.3 3.3 - - - - - - 2.2 4.3 3.3 - - - - - - 2.2 4.3 3.3 - - - - - - 2.2 4.3 3.3 Rosmarinus officinalis 3.3 4.4 4.4 3.4 - - - - 3.4 4.4 4.4 4.4 2.3 - - - - 2.2 3.4 4.4 4.4 2.2 - - 1.1 2.2 3.3 3.4 4.3 4.4 2.3 - -
Figura 2. Localització de les subpoblacions de romer «1» i Tyrimnus leucographus «2» al Tossal de l’Infern. Font de l’ortofoto: Institut Cartogràfic i Geològic de Catalunya (2021).



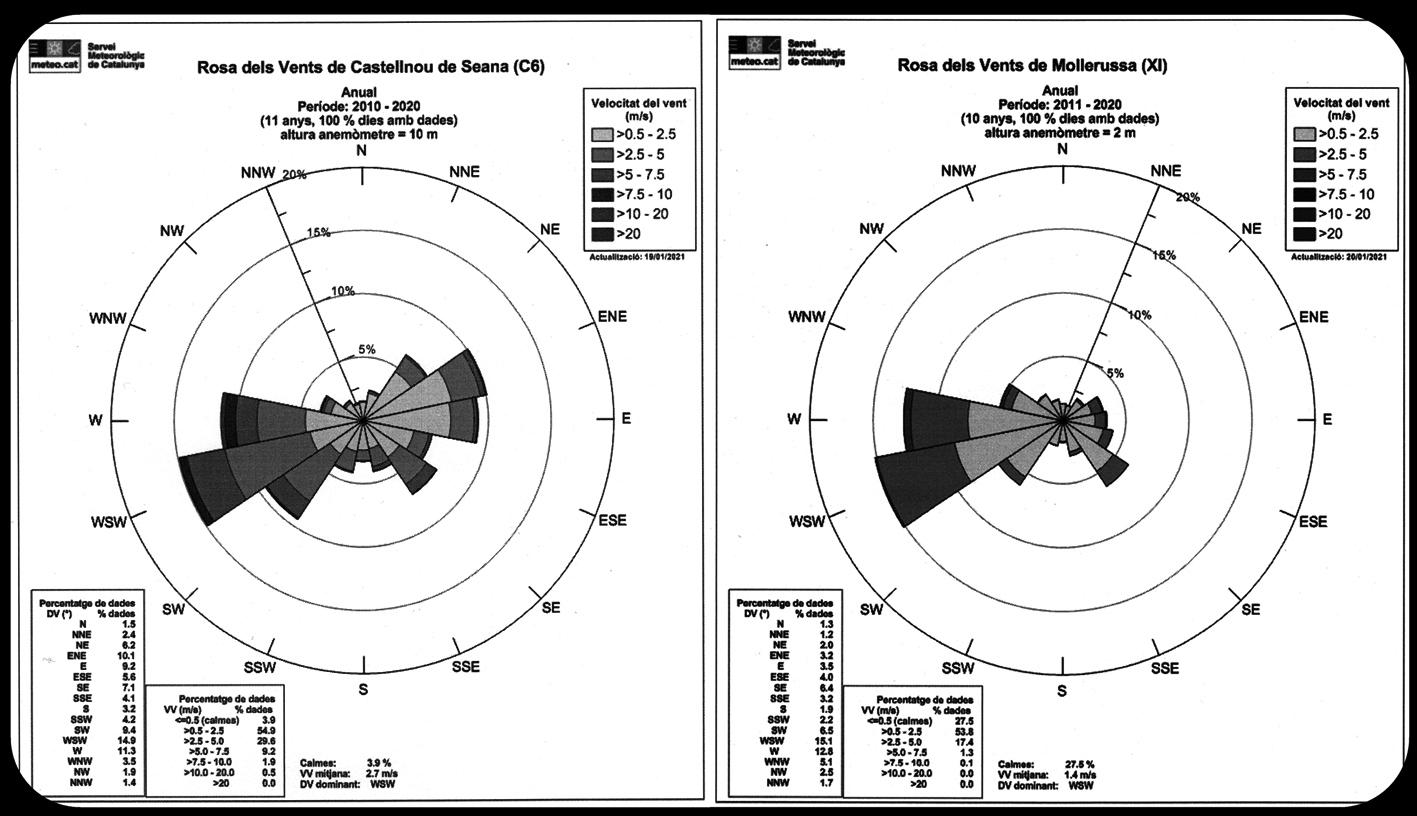
Resultats
Distribució local del romer
(Meteocat), concretament les de les estacions automàtiques del Pla d’Urgell, prenent Mollerussa (XI), la més propera al Tossal de l’Infern i la Serra, com a referència més important. S’han elaborat taules que contenen els resums de l’estació de Mollerussa des de l’any 2014 al 2020 i elements d’altres estacions que hem cregut rellevants. S’han comparat les dades meteorològiques i fenològiques per tal de veure si es dona alguna relació.
La població de R. officinalis del Tossal de l’Infern està dividida en dues subpoblacions d’un centenar i al voltant de 40 exemplars, separades per uns 30 m, les quals se situen (vegeu fig. 2) a la part SW del turó (31TCG2207), en els entrants que ha obert l’erosió. La major part dels seus individus es troben uns 5 m per davall de la plataforma que culmina el tossal (300 m), ben arrenglerats i orientats cap al S (gairebé 180º). Aquesta disposició i l’orientació del tossal en sentit ESE-WNW (vegeu fig. 2), afavoreixen la seva exposició als vents de component W (seré o ponent), SW (morella o garbí) i SE (marinada), però, en canvi, possibiliten que estiguin a resguard dels vents del NW (cerç o mestral), E (llevant), N i NE. Entre la part més alta del tossal –un dels punts més elevats de la comarca– i la base, hi ha un desnivell aproximat de 15 m i uns 70 m respecte a les parts més fondes. L’únic exemplar de la Serra, es localitza prop de l’Observatori (terme de Torregrossa, 31TCG1909, 279 m), mirant cap al S i en una petita depressió ocasionada per l’extracció de graves que fa que no sigui gaire afectat pels vents. Al Tossal Gros, orientat en sentit E-W (31TCG2501, 342 m) el romer es troba a
194 Butlletí ICHN 85 (4), 2021
FLORA ET FAUNA
GEA,
Figura 3. Flors de romer afectades per les gelades. Tossal de l’Infern (gener de 2019).
Figura 4. Tossal de l’Infern: els vestigis de carrascar i la brolla on destaca Stipa offneri
Figura 5. La població de romer del Tossal de l’Infern se situa a les parts altes del vessant S.
A. MAYORAL ARQUÉ
Figura 6. Roses dels vents de Castellnou de Seana (2010-2020) i Mollerussa (2011-2020). Meteocat (2021a,b)
Taula 2. Dades meteorològiques enregistrades a l’estació de Mollerussa (2014-2020). PPT=precipitació anual, TMM= T mitjana anual, TXM= T màxima mitjana; TNM= T mínima mitjana, TXX= T màxima absoluta, TNN=T mínima absoluta (entre parèntesis la data en què es van assolir), Vm= velocitat mitjana anual vent i direcció dominant (Mollerussa, 2m; Castellnou, 10 m), Hr= humitat relativa mitjana, I. solar= mitjana irradació solar global diària. Elaboració pròpia a partir de Meteocat (2014-20a,b).
Taula 3. Estació de Mollerussa. Temperatures màximes (TXM) i mínimes (TNM) mitjanes mensuals i dies amb gelades (T≤ 0) dels anys 2014, 2015 i 2020. Elaboració pròpia a partir de Meteocat (2014-20b).
Taula 4. Precipitacions mensuals i anuals (mm) enregistrades a les quatre estacions del Pla d’Urgell durant l’any 2015. Elaboració pròpia a partir de Meteocat (2014-20b).
les parts més altes i als vessants S i N, en canvi, Globularia alypum L., una espècie termòfila que sol tenir la floració plena al febrer, hi és prou abundant, però, no creix al vessant N. Tanmateix, un sol exemplar situat en una zona propera al Tossal de l’Infern (els Colls, 31TCG2306, 295 m) creix en orientació NNE. Pel que fa a la Serra de Bellmunt-Almenara, orientada en sentit ESE-WNW, la població de romer que s’ha tingut en compte –situada entre els termes de la Sentiu de Sió i Bellcaire d’Urgell (31TCG2428, 2528, 330 m)– es troba sobre els guixos que afloren a la vall invertida de l’anticlinal, formant part de l’associació Ononidetum tridentatae.
Pel que fa als resultats del seguiment fenològic del romer (vegeu la taula 1), hem pogut constatar que la seva floració va des del mes de novembre a l’abril, amb un màxim situat, aproximadament, entre mitjans de febrer i la primera meitat de març. Només en una ocasió (2020) es va observar a finals d’octubre algun individu amb molt poques flors. És important destacar l’elevada floració que assoleix al gener, precisament durant aquest mes, algun any, per exemple el 2019, vam poder observar flors músties i groguenques per l’efecte de les gelades (vegeu fig. 3); tanmateix, amb la millora de les condicions meteorològiques, la major part dels exemplars
195 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA
2014 2015 2016 2017 2018 2019 2020 PPT (mm) 425,7 199,7 353,1 290,0 506,4 437,7 455,7 TMM (ºC) 14,7 14,4 14,3 14,2 14,6 14,6 14,7 TXM (ºC) 21,6 21,6 21,1 21,7 21,4 21,8 21,5 TNM (ºC) 8,7 7,8 8,1 7,2 8,4 7,7 8,5 TXX (ºC) 35.8 (16/7) 41,4 (7/7) 37,8 (10/7) 38,2 (3/8) 38,7 (2/8) 42,1 (29/6) 39,4 (8/8) TNN (ºC) -6,2 (31/12) -5,6 (2/1) -6,1 (17/1) -9,1 (18/1) -6,8 (9/2) -5,7 (11/1) -4,0 (27/12) Vm vent (2 m) 1,6 W 1,4 W 1,4 W 1,3 W 1,4 W 1,5 W 1,3 W Vm vent (10 m) 2,8 SW 2,7 SW 2,7 W 2,6 W 2,6 W 2,8 W 2,6 W Hr mitjana 68 % 66 % 66 % 70 % 73 % 69 % 75 % I. solar MJ/m2 16,5 16,6 16,6 17,2 16,2 17,1 16,3 Dies gelada 25 57 35 71 48 60 34
G F M A M J J A S O N D TNM (ºC)- 2014 2,6 1,7 3,3 8,6 9,0 14,0 15,5 16,7 15,3 9,7 5,9 1,2 TNM (ºC)- 2020 1,3 2,6 4,7 8,0 12,6 14,1 16,8 16,9 12,5 6,3 4,7 1,5 TXM (ºC)- 2014 11,7 12,9 17,7 22,5 24,4 29,8 30,4 30,6 27,9 24,6 16,9 9,7 TXM (ºC)- 2020 8,4 16,3 16,8 20,4 26,4 28,5 33,4 32,9 27,9 20,8 15,6 10,1 Dies Gelada- 2014 5 8 3 0 0 0 0 0 0 0 1 8 Dies Gelada- 2015 19 15 5 2 0 0 0 0 0 0 6 10 Dies Gelada- 2020 9 9 1 0 0 0 0 0 0 2 4 9
CODI I NOM EMA G F M A M J J A S O N D Any C6 Castellnou de Seana 11,2 16,7 12,4 8,0 4,0 25,6 32,2 24,8 13,8 2,9 12,7 3,5 167,8 V8 el Poal 10,3 15,8 15,6 10,4 3,8 33,4 33,9 33,3 15,1 6,7 20,9 3,4 202,6 WC Golmés 11,0 18,4 11,6 5,7 4,9 23,0 35,0 18,8 18,1 5,5 13,5 2,8 168,3 XI Mollerussa 10,2 20,1 21,7 5,2 5,0 15,1 45,6 22,2 21,6 5,0 24,6 3,4 199,7
LA GAIREBÉ ABSÈNCIA DE ROSMARINUS OFFICINALIS (LAMIACEAE) AL PLA D'URGELL
SOBRE

s’havien recuperat i estaven ben florits al febrer. Els resultats de les poblacions de romer del Tossal Gros i dels guixos de la Sentiu de Sió-Bellcaire d’Urgell, són congruents amb el suara esmentat.
Anomalies fenològiques
Els resultats de les plantes que van florir de manera inusual també són recollits a la taula 1. Val a dir que la gran majoria d’aquestes són mates –sobretot camèfits i algun hemicriptòfit (les Stipa) i un nanofaneròfit (G. scorpius)– la major part, típiques dels Rosmarinetalia i totes tenen la seva normal floració a la primavera, entre març i juny; un aspecte que queda ben reflectit a la taula que ens ocupa. L’anomalia, en primer lloc, queda manifestada per la floració durant el desembre de 2014 de totes les plantes considerades i també al novembre amb l’excepció del gènere Teucrium. Aquesta floració conjunta en els mesos esmentats, no es repeteix en els altres anys, només es detecten floracions esporàdiques dels heliantems. Durant el gener de 2015, els representants del gènere Stipa, enregistren els índexs de floració més elevats de tots els anys per aquest mateix mes, aspecte que també es compleix –i fins i tot continua al febrer– en el cas de Genista i Thymus. L’altra cara de l’anomalia es fa palesa en els mesos d’abril, maig i juny de 2015, tots els tàxons mostren, per aquest període de temps, els índexs de floració més baixos de
tots els anys estudiats. Stipa offneri és l’espècie que destaca més en aquest descens: tots els anys, experimenta la plenitud de la seva floració (4.4) durant el maig, l’excepció es dona el 2015 amb un clar descens (2.2). Hem de ressaltar que el romer no experimenta variacions destacades dels seus índexs en els anys 2014 i 2015.
Discussió
La població de R. officinalis del Tossal de l’Infern i l’exemplar de la Serra, són les dues úniques localitats del Pla d’Urgell. La ubicació dels individus, en tractar-se d’organismes que viuen en unes condicions prou extremes, ens pot donar una informació molt interessant: els nostres exemplars estan ben arrenglerats en direcció S, se situen a les parts altes dels turons, però eviten les zones de màxima altitud, busquen les petites depressions que han excavat l’erosió diferencial o bé l’acció antròpica, són sotmesos (vegeu figs. 2 i 6 i taula 2) als vents més freqüents i intensos de component W-SW, però queden poc exposats als del grup del cerç o mestral (WN, NW, NNW), els quals són els més freds i secs als mesos hivernencs. Curiosament, aquesta especial disposició, també la segueixen uns pocs exemplars de dues plantes termòfiles –Cistus clusii Dunal subsp. clusii i G. alypum– que es troben al Tossal de Montperler, vers Bellpuig d’Urgell (31TCG3710, 336 m). En el mateix sentit, hem de destacar (vegeu fig. 7) la detecció l’any 2018 al Tossal de l’Infern de Tyrimnus leucographus (L.) Cass., una espècie rara a Catalunya que se sol localitzar en contrades més properes a la línia litoral i que recentment ha estat indicada a les Garrigues (Molero et al., 2006). T. leucographus, al Tossal de l’Infern, s’encara també cap al S, però a diferència del romer, es fa a les parts baixes dels entrants produïts per l’erosió (vegeu fig. 2), fins i tot en una petita fondalada, només oberta vers el migdia. Aquest vegetal floreix de maig a juliol, la seva fenologia no ha de suportar els freds hivernencs i per tant no li cal pujar als llocs més enlairats del tossal. L’exemplar de G. alypum de la zona dels Colls, tot i que un sol individu no és massa significatiu, també l’hem localitzat a les parts altes del turó, en un vessant on no toquen els vents del grup del cerç. És ben clar que el romer i les espècies termòfiles, en situar-se a les parts superiors dels tossals, poden disminuir l’efecte de la inversió tèrmica. Nosaltres mateixos, de bon matí, en el nostre trajecte diari al lloc de treball, hem pogut verificar sovint aquest fenomen quan la carretera N-II passa per la vall del riu Corb: durant les calmes anticiclòniques dels mesos hivernencs, un desnivell d’uns 10 m, provocava en el termòmetre del nostre cotxe, descensos de 0,5 a 1 ºC. Els resultats del nostre estudi sobre el romer semblen confirmar les conclusions de Terradas (1973, 1986) sobre els efectes de la inversió tèrmica i el paper de les turbulències dels rius importants i barrancs associats en la distribució de la vegetació. En el nostre territori, el tipus de substrat no condiciona la distribució del romer, aquest se situa a les parts altes dels turons i busca el vessant S per tal de disminuir el fred i les fortes gelades que provoca la inversió tèrmica, alhora que s’aixopluga dels vents més gèlids. Tal com passa amb algunes plantes termòfiles, es fa
196 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA
MAYORAL ARQUÉ
Figura 7. Tyrimnus leucographus a les parts baixes del vessant S del Tossal de l’Infern.
A.
SOBRE
cada vegada més rar a mesura que ens anem allunyant de la influència marítima i fins i tot no creix en altituds superiors als 400 m, com és el cas de la Serra de Bellmunt-Almenara. Reapareix quan aquest anticlinal s’acosta al riu Segre (Bellcaire d’Urgell, la Sentiu de Sió), on les turbulències podrien fer minvar l’estancament de l’aire fred. Tots aquests aspectes, ens porten a suggerir les temperatures mínimes originades per la inversió tèrmica com el factor que limita la seva distribució al Pla d’Urgell i terres properes. Quan es produeixen fortes gelades, les flors del romer queden afectades, esdevenen grogues i pansides, tanmateix els individus es recuperen i tornen a florir, fora dels microclimes que es donen en els tossals estudiats, en les zones més fondes i continentals, les intenses gelades podrien afectar molt més el desenvolupament d’aquesta floració.
No sabem la causa que pot tenir a veure amb l’anomalia de la floració de determinats tàxons detectada entre desembre de 2014 i els primers mesos de 2015. En la caracterització climàtica de 2014 (Meteocat, 2014) s’esmenta que ha estat un dels anys més càlids d’ençà que es tenen dades i s’assenyalen les temperatures molt elevades d’abril, octubre, novembre i gener. Les temperatures màximes i mínimes mitjanes excepcionalment elevades d’octubre i novembre d’aquest any –arriben a superar les de 2020 que és també un dels més càlids de la sèrie estudiada (vegeu taules 2 i 3)– podrien haver desencadenat el fet anterior, encara que la major part d’aquesta floració va tenir lloc de desembre de 2014 a febrer de 2015, període que va experimentar una elevada concentració de gelades (vegeu taula 3). El descens de l’índex de floració de les espècies estudiades durant la primavera de 2015, podria tenir relació amb la molt escassa precipitació que s’hi va donar. En l’informe de 2015 (Meteocat, 2015), es destaca l’anomalia pluviomètrica del Pla d’Urgell, la qual va quedar reflectida en la totalitat de les seves estacions. Si analitzem de manera més acurada la precipitació de 2015 (vegeu la taula 4), ens sorprèn que el mes amb la precipitació més elevada sigui juliol quan generalment és el més sec; la xifra de maig que sol esdevenir un dels més plujosos, és força baixa, la resta de mesos primaverals i els mesos de tardor són també prou secs. Cal fer un seguiment del comportament d’aquestes plantes per tal de comprovar si pot ser útil en la predicció d’anys excepcionalment secs. Els representants del gènere Stipa són els que han experimentat més intensament aquesta anomalia, S. offneri la més sensible, no tant S. parviflora, una planta amb connotacions més estèpiques; no hem notat cap irregularitat a les altres quatre espècies (S. iberica Martinovský, S. barbata Desf., S. capillata L., S. lagascae Roem. & Schult.) presents al nostre territori. L’anomalia no ha estat condicionada per l’orientació, a excepció de S. parviflora que té predilecció pels vessants S, les altres espècies han florit tant en terreny pla com en diferents vessants. Durant la primavera de 2015, hem pogut constatar també una disminució del grau floració de les espècies no afectades per l’anomalia. Tot i que l’estudi i el seguiment s’han centrat al Tossal de l’Infern i la Serra, en el Tossal Gros observàrem que les espècies considerades també la van presentar, però amb menys intensitat, no la vam detectar a la zona de Bellcaire i la Sentiu. En aquest indret, sí
que vàrem observar un comportament irregular de Gypsophila struthium L. subsp. hispanica (Willk.) G. López en els guixos afectats per un incendi succeït l’estiu de 2014; de finals de desembre d’aquest mateix any fins gener de 2015, moltes de les mates d’aquesta planta van rebrotar i florir abastament. Aquest fenomen no es va donar a les zones no cremades properes. En aquest lloc, aquest tàxon floreix sobretot a l’estiu (Guàrdia & Ninot, 1991). Sembla ser que l’incendi pot tenir a veure amb el comportament anormal de Gypsophila, però, han pogut influir també les anomalies meteorològiques de 2014-2015? Cal perseverar en l’estudi del comportament d’aquest endemisme.
Un altre detall interessant és que al llarg dels anys estudiats, el ritme fenològic del romer no s’ha vist alterat per les irregularitats meteorològiques i les plantes que han experimentat l’anomalia, en certa manera, l’han intentat imitar. El canvi climàtic que ja estem patint, podrà esperonar el desenvolupament del romer i les plantes termòfiles a les parts més baixes i fredes del Pla d’Urgell? No ho sabem, però en aquesta comarca, l’acció antròpica, d’ençà la construcció del Canal d’Urgell, ha provocat la desaparició en molts indrets de la natura adaptada a les condicions climatològiques que s’hi donen. Són cada cop menys, cal vetllar per la preservació d’aquestes zones privilegiades per petites que siguin, la informació que ens donen els organismes que s’hi fan, és massa valuosa i no ens podem permetre perdre-la.
Agraïments
Les consideracions sobre l’escrit fetes per Llorenç Sáez i Pere Aymerich ens han estat molt valuoses.
Bibliografia
Boixadera, J., Danés, R. & Herrero, C. 2009. Mapa de sòls de Catalunya 1:25.000 Mollerussa 389-1-1 (65-29). Institut Geològic de Catalunya, Departament d’Agricultura, Alimentació i Acció Rural, Institut Cartogràfic de Catalunya. Barcelona.
Busqué, J., Calvet, J., Martí, E., Sau, T. & Martín-Vide, J. 1989. La geografia dels vents a Catalunya. Revista Catalana de Geografia, segona època, 4: 62-73.
Calvet, J. 1977. Contribución al conocimiento geomorfológico de la Depresión Central Catalana. Tesi doctoral inèdita. Universitat de Barcelona. 331p.
Conesa, J.A., Mayoral, A., Pedrol, J. & Recasens, J. 1994. El paisatge vegetal dels espais d’interès natural de Lleida: àrea meridional. Institut d’Estudis Ilerdencs. Lleida. 194 p.
Guàrdia, R. & Ninot, J.M. 1991. Models de comportament fenològic als prats secs i a les brolles de l’àrea meridional de la Noguera. Butlletí de la Institució Catalana d’Història Natural, 59: 117-127.
Institut Cartogràfic i Geològic de Catalunya. 2021. Mapa del Pla d’Urgell i ortofotomapa del Tossal de l’Infern. Disponible a: http://www.icc.cat/vissir3/ [ Data de consulta: 13 setembre 2021]
Mayoral, A. 1986. Estudis sobre la vida vegetal al Pla d’Urgell. Tesi de llicenciatura inèdita. Universitat de Barcelona. 186 p.
Meteocat. 2014. Dades de la XEMA: introducció, llistat, mapa de localització i caracterització climàtica. Disponible a https:// static-m.meteo.cat/wordpressweb/wp-content/uploads/2014/11/
197 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA
LA GAIREBÉ ABSÈNCIA DE ROSMARINUS OFFICINALIS (LAMIACEAE) AL PLA D'URGELL
GEA, FLORA ET FAUNA
18151719/EMAs_2014.pdf [Data de consulta: 10 juliol 2021].
Meteocat. 2015. Dades de la XEMA: introducció, llistat. mapa de localització i caracterització climàtica. Disponible a https:// static-m.meteo.cat/wordpressweb/wp-content/uplads/2014/11/ 19085959/EMAs2015 [Data de consulta 10 juliol 2021].
Meteocat. 2014-2020a. Resums anuals (2014-20) de la EMA de Mollerussa. Disponible a https: // www.meteo.cat/ wpweb/climatologia/serveis-i-dades-climatiques/anuaris-dedades-meteorologiques/xarxa-destacions-meteorologiquesautomatiques/ [ Data de consulta: 10 juliol 2021].
Meteocat. 2014-2020b. Taules de dades mensuals 2014-20 (Mollerussa, Castellnou, el Poal, Golmés). Disponible a https:// www.meteo. cat/wpweb/climatologia/serveis-i-dades-climatiques/anuarisde-dades-meteorologiques/xarxa-destacions-meteorologiquesautomatiques/ [Data de consulta: 10 juliol 2021].
Meteocat. 2021a. Rosa dels vents climàtica anual (Castellnou de Seana). Disponible a https:// static-m.meteo.cat/content/ climatologia/roses-vent/C6_rosa_clim_0.png [Data de consulta: 10 juliol 2021].
Meteocat. 2021b. Rosa dels vents climàtica anual (Mollerussa). Disponible a https://static-m.meteo.cat/ content/climatologia/rosesvent/XI_2_rosa_clim_0.png [ Data de consulta: 10 juliol 2021].
Molero, J., Sáez, L., Vallverdú, J. & Valle, L.G. 2006. Noves aportacions al coneixement de la flora vascular de les comarques meridionals de Catalunya. III. Orsis, 21: 19-39.
Terradas, J. 1973. Clima y economía hídrica en comunidades vegetales de Los Monegros. Tesi doctoral inèdita. Universitat de Barcelona.
Terradas, J. 1986. El paisatge vegetal dels Monegros: assaig d’interpretació. Orsis, 2: 71-95.
198 Butlletí ICHN 85 (4), 2021
A. MAYORAL ARQUÉ
Aportacions al coneixement de la flora vascular del sector oriental del massís del Montseny (Catalunya, NE de la península Ibèrica)
Pere Barnola Echenique*, Josep Maria Panareda Clopés** & Josep Gesti Perich***
* C/ Santa Rosa, 46. 08470 Sant Celoni.
** C/ Lluís Montané, 2, 3, 3. 08470 Sant Celoni.
*** C/ Sant Ignasi, 42. 17430 Santa Coloma de Farners.
Autor per la correspondència: Josep Gesti. A/e: josepgesti@gmail.com
Rebut: 07.11.2021; Acceptat: 28.11.2021; Publicat: 30.12.2021
Resum
Aportem dades sobre 43 tàxons de plantes vasculars (13 autòctons i 30 aŀlòctons) observats al massís del Montseny i que no hi havien estat citats anteriorment. Així mateix, donem informació sobre la distribució de 57 tàxons més (36 autòctons i 21 aŀlòctons), dels quals en constatem la presència al sector oriental i/o a la zona basal d’aquest massís, on no havien estat indicats fins el moment.
Paraules clau: flora vascular, massís del Montseny, Catalunya, península Ibèrica.
Abstract
Contributions to the knowledge of the vascular flora of the eastern sector of the Montseny massif (Catalonia, NE Iberian Peninsula)
We provide data on 43 vascular plant taxa (13 native and 30 non-native) observed in the Montseny massif, not previously reported in this area. We also provide information on the distribution of 57 more taxa (36 native and 21 non-native) in the eastern sector and in the basal zone of this massif, where they had not been indicated so far.
Key words: vascular flora, Montseny massif, Catalonia, Iberian Peninsula.
Introducció
El Montseny és un espai amb una llarga història de prospeccions botàniques que es remunten més de tres segles enrere (vegeu-ne una síntesi a Saez et al., 2017), la qual cosa ha fornit una quantitat molt elevada d’informació sobre la seva flora. Atenent el recompte de citacions en cadascun dels quadrats UTM de 10 km de costat (Font, 2021) que l’atenyen, el Montseny forma part de les àrees més ben prospectades de Catalunya, amb diversos quadrats que disposen de més de 18.000 citacions de plantes vasculars. Això no obstant –com és natural en qualsevol espai d’aquestes dimensions– aquest coneixement és desigual en els diferents sectors que conformen el massís. Així, per exemple, algunes localitats «clàssiques» com ara Santa Fe del Montseny, les Agudes o el turó de l’Home, acumulen un nombre molt elevat de citacions. Per contra, algunes àrees de la part baixa han estat poc prospectades, probablement perquè no han despertat tant d’interès entre els científics com els estatges superiors que tenen una singularitat biogeogràfica evident.
Al present treball aportem dades sobre la flora del municipi de Riells i Viabrea (al sector oriental del Montseny), una
d’aquestes àrees comparativament poc estudiades. En el decurs del projecte de catalogació de la flora vascular d’aquest municipi (http://flora.riellsiviabrea.cat), impulsat pel grup de Flora Catalana del Baix Montseny, hem pogut localitzar alguns tàxons dels quals no ens consten indicacions anteriors al massís. Així mateix, per altres espècies que ja eren citades al Montseny, aquest treball ens ha permès constatar la seva presència en àrees d’on no eren conegudes, ja sigui al sector oriental o bé a la part baixa d’aquest espai.
Material i mètodes
Les dades que aportem corresponen a plantes vasculars (autòctones i aŀlòctones creixent fora de conreu) observades al municipi de Riells i Viabrea durant les prospeccions fetes pels autors en els darrers tres anys i també en campanyes portades a terme per un de nosaltres (JMPC) entre 1999 i 2016. Aquest municipi, situat dins del sector oriental del massís del Montseny, ocupa una extensió aproximada de 27 km2 amb altituds que van des dels 80 fins als 1.312 metres sobre el nivell del mar.
199 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA GEA, FLORA ET FAUNA
Butlletí de la Institució Catalana d’Història Natural, 85 (4): 199-207. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition) DOI: 10.2436/20.1502.01.93
GEA,
La informació s’organitza en tres apartats:
Tàxons no indicats prèviament al massís del Montseny: hem revisat, entre d’altres, la informació continguda als catàlegs florístics del massís (Bolòs et al., 1986; Sáez et al., 2017), en publicacions que contenen dades procedents d’aquest espai (Barnola & Romo, 1989; Gutiérrez & Sáez, 1996; Gutiérrez, 1998, 1999; Barnola et al., 2014; Pié & Vilar, 2004; Panareda & Nuet, 2005; Mercadé & Pérez-Haase, 2013; Pié et al., 2014; Nuet; 2015; Sáez et al., 2015; Mercadal, 2019; González & Nuet, 2020; etc.), al Banc de Dades de Biodiversitat de Catalunya (Font, 2021), i a les bases de dades d’herbaris de l’Institut Botànic de Barcelona (BC), del Centre de Documentació de Biodiversitat Vegetal de la Universitat de Barcelona (BCN) i de la Universitat de Girona (HGI). Hi incloem alguns tàxons dels quals només hi havia indicacions referides a quadrats UTM de 10 km de costat (ORCA, 1985-2014; Bolòs, 1998) que cobreixen parcialment el massís, però sense localitats concretes que permetessin confirmar que eren citacions del Montseny.
Tàxons ja coneguts al Montseny, però no indicats prèviament al sector oriental del massís: en base a les mateixes fonts assenyalades a l’apartat precedent i incloent també aquells tàxons dels quals només constaven referències genèriques de quadrats UTM parcialment inclosos al massís.
Tàxons no indicats prèviament a la zona basal del massís: entesa com la que va fins als 500 m d’altitud, coincidint amb la definida pels dos intervals altitudinals basals (<250 m i 250-500 m) reconeguts al catàleg de Sáez et al. (2017).
Les localitats concretes on hem observat els tàxons es recullen a l’apèndix. S’hi presenten en ordre alfabètic segons el seu nom científic i, per cadascun, donem la localització, el quadrat UTM d’1 km de costat (totes les coordenades pertanyen a la zona 31T, sistema de referència ETRS89), l’altitud, l’hàbitat, el plec d’herbari (si n’hi ha) i la data d’observació. D’aquells tàxons dels quals n’hem observat moltes localitats en llistem només 5, que són representatives dels diferents aspectes tractats en aquest treball (localitats a cotes baixes de la zona basal, distribució en diferents àrees del sector oriental, etc.).
Resultats i discussió
Tàxons no indicats prèviament al massís del Montseny
A la columna «Mon» de la Taula 1 assenyalem 43 tàxons (13 autòctons i 30 aŀlòctons) que hem observat a Riells i Viabrea i dels quals no ens consten citacions anteriors al massís del Montseny.
Taula 1. Nous tàxons observats al Montseny. Nat: grau de naturalització local (C: casual; E: establert), Mon: novetat per al massís del Montseny; Est: novetat per al sector oriental del massís; Bas: novetat per l’àrea basal del massís (entre claudàtors s’indica l’altitud mínima observada). Els noms dels tàxons aŀlòctons van precedits d’un asterisc (*)
Tàxon Família Nat Mon Est Bas Observacions
Agrimonia eupatoria L. subsp. grandis Rosaceae + + (Andrz. ex Asch. & Graebn.) Bornm
Agrostis capillaris L. subsp. capillaris Poaceae + [220 m]
*Alcea rosea L. [Althaea rosea (L.) Cav.] Malvaceae C + + + [110 m]
Allium ampeloprasum L. Amaryllidaceae +
Aristolochia paucinervis Pomel [A. longa L. Aristolochiaceae + Indicada al quadrat UTM DG51 subsp. paucinervis (Pomel) Batt.] (ORCA)
Avenella flexuosa (L.) Drejer Poaceae + [485 m] [Deschampsia flexuosa (L.) Trin.]
*Bassia scoparia (L.) A.J. Scott Amaranthaceae C + [Kochia scoparia (L.) Schrad.]
*Berberis aquifolium Pursh Berberidaceae C + [90 m]
Bolboschoenus maritimus (L.) Palla Cyperaceae + [95 m]
[Scirpus maritimus L.]
Brachypodium distachyon (L.) P. Beauv. Poaceae +
*Brassica napus L. Brassicaceae C + + [95 m]
Bromus lanceolatus Roth Poaceae + + + [100 m]
Campanula erinus L. Campanulaceae +
*Campsis radicans (L.) Seem. Bignoniaceae C + + [95 m]
*Carpobrotus edulis (L.) N.E. Br. Aizoaceae C + + + [240 m]
*Cedrus deodara (Lamb.) G. Don. Pinaceae C + + + [250 m]
Centaurium pulchellum (Swartz) Druce Gentianaceae + [270 m]
*Cerastium tomentosum L. Caryophyllaceae C +
*Cercis siliquastrum L. Fabaceae C +
Cirsium monspessulanum (L.) Hill Asteraceae +
*Cotula australis (Sieber ex Spreng.) Hook. f. Asteraceae E + +
Cruciata glabra (L.) Ehrend. Rubiaceae + [250 m]
200 Butlletí ICHN 85 (4), 2021
FLORA
P. BARNOLA ECHENIQUE ET AL.
ET FAUNA
Tàxon Família Nat Mon Est Bas Observacions
*Cyperus alternifolius L. subsp. Cyperaceae C + + + [125 m] flabelliformis (Rottb.) Kük
*Dichondra micrantha Urb. Convolvulaceae E + + + [85 m]
*Echinochloa colonum (L.) Link Poaceae E + + + [85 m]
Elymus caninus (L.) L. Poaceae +
Elymus pungens (Pers.) Melderis subsp. Poaceae + + + [120 m] campestris (Godr. & Gren.) Melderis
*Epilobium brachycarpum C. Presl Onagraceae E + + + [85 m]
Epilobium tetragonum L. Onagraceae + [175 m]
Equisetum × moorei Newman Equisetaceae + + + [220 m]
Equisetum ramosissimum Desf. Equisetaceae + [85 m]
Eragrostis minor Host Poaceae + + + [85 m]
*Eriobotrya japonica (Thunb.) Lindl. Rosaceae C + + + [100 m]
*Eschscholzia californica Cham. Papaveraceae C +
*Eucalyptus camaldulensis Dehnh. Myrtaceae C + [175 m]
*Eucalyptus globulus Labill. Myrtaceae C + [210 m]
*Euphorbia serpens Kunth Euphorbiaceae E + + [90 m]
*Gazania rigens (L.) Gaertn. Asteraceae C + +
Glebionis coronaria (L.) Cass. ex Spach Asteraceae + + + [120 m] [Chrysanthemum coronarium L.]
Heteropogon contortus (L.) Beauv. Poaceae + + + [155 m] Indicada al quadrat UTM DG41 ex Roem. & Schultes (ORCA)
Iris pseudacorus L. Iridaceae + [100 m]
*Lamium galeobdolon (L.) Crantz subsp. Lamiaceae C + + + [145 m] argentatum (Smejkal) J. Duvig.
Lathyrus niger (L.) Bernh. Fabaceae + [375 m]
*Ligustrum lucidum W.T. Aiton Oleaceae E +
Lotus angustissimus L. subsp. suaveolens Fabaceae + + + [320 m] (Pers.) O. Bolòs & Vigo
Lupinus angustifolius L. Fabaceae + [80 m]
*Malva arborea (L.) Webb & Berthel. Malvaceae C + + + [315 m] [Lavatera arborea L.]
*Melia azedarach L. Meliaceae C + + + [105 m]
Melilotus officinalis (L.) Lam. Fabaceae + Indicada als quadrats UTM DG42, 51 i 62 (ORCA)
*Morus alba L. Moraceae E + + + [85 m]
Myosotis sylvatica Hoffm. subsp. Boraginaceae + [380 m]
teresiana (Senn.) O. Bolòs & Vigo
Narcissus tazetta L. Amaryllidaceae + + [110 m]
*Nigella damascena L. Ranunculaceae C + + + [120 m]
*Nothoscordum gracile (Aiton) Stearn Amaryllidaceae C + + + [115 m] Indicada al quadrat UTM DG51 [incl. N. × borbonicum Kunth] (ORCA)
*Oenothera oehlkersi Kappus Onagraceae E + + + [120 m]
Onopordum acanthium L. Asteraceae + + [85 m]
*Osteospermum ecklonis (DC.) Norl. Asteraceae C + + + [105 m] [Dimorphotheca ecklonis DC.]
Oxalis acetosella L. Oxalidaceae + [500 m]
*Panicum capillare L. Poaceae E + + + [80 m]
Papaver somniferum L. subsp. Papaveraceae + + + [85 m] setigerum (DC.) Arcang.
*Passiflora caerulea L. Passifloraceae C + + [95 m]
Phalaris paradoxa L. Poaceae + + + [120 m]
*Phyla canescens (Kunth) Greene Verbenaceae E + [Lippia filiformis Schrad.]
*Physalis ixocarpa Brot. ex Hornem. Solanaceae E +
*Pittosporum tobira (Thunb.) W.T. Aiton Pittosporaceae C + + + [85 m]
Plantago major L. subsp. Plantaginaceae + [380 m] intermedia (Gilib.) Lange
*Plumbago auriculata Lam. Plumbaginaceae C + + + [125 m]
201 Butlletí ICHN 85 (4), 2021
GEA, FLORA ET FAUNA
APORTACIONS AL CONEIXEMENT DE LA FLORA VASCULAR DEL SECTOR ORIENTAL DEL MASSÍS DEL MONTSENY
GEA, FLORA ET FAUNA
Tàxon Família Nat Mon Est Bas Observacions
*Populus deltoides Marshall Salicaceae C + + + [220 m]
*Prunus domestica L. Rosaceae C + [105 m]
*Prunus dulcis (Mill.) D.A. Webb Rosaceae C +
Pseudoturritis turrita (L.) Brassicaceae + [330 m]
Al-Shehbaz [Arabis turrita L.]
*Punica granatum L. Lythraceae C + [165 m]
*Pyracantha fortuneana (Maxim.) H.L. Li Rosaceae E + + + [100 m]
*Pyrus communis L. Rosaceae C + [95 m]
Ranunculus sardous Crantz subsp. Ranunculaceae + + + [100 m] trilobus (Desf.) Rouy & Fouc
Reseda luteola L. subsp. luteola Resedaceae + [85 m]
Rosa pouzinii Tratt. Rosaceae + Indicada al quadrat UTM DG52 (ORCA)
Scolymus hispanicus L. Asteraceae +
Scorzonera angustifolia L. Asteraceae +
Serratula tinctoria L. Asteraceae + [175 m]
Setaria adhaerens (Forssk.) Chiov. Poaceae + + + [85 m] [S. verticillata (L.) P. Beauv. subsp. aparine (Steudel) Asch.]
*Silene pseudoaction Desf. Caryophyllaceae C + + + [105 m]
*Sinapis arvensis L. Brassicaceae E + + + [110 m]
*Solanum tuberosum L. Solanaceae C +
Sparganium erectum L. Typhaceae + [100 m]
*Tagetes patula L. Asteraceae C + + + [95 m]
Tilia platyphyllos Scop. Malvaceae + [215 m]
Torilis arvensis (Huds.) Link subsp. arvensis Apiaceae + [90 m]
*Trachycarpus fortunei (Hook.) H. Wendl. Arecaceae C + + + [85 m]
*Tradescantia fluminensis Velloso Commelinaceae C + + + [125 m]
Tragopogon porrifolius L. Asteraceae + + [110 m] Indicada a Sant Feliu de Buixalleu (Llensa, 1945: “incierta”)
Trifolium ligusticum Balb. ex Loisel. Fabaceae + + + [240 m]
*Ulmus pumila L. Ulmaceae C + [95 m]
Urospermum picroides (L.) Scop. Asteraceae + Indicada al quadrat UTM DG62 ex F.W. Schmidt (ORCA)
Veronica hederifolia L. Plantaginaceae + [90 m]
Veronica serpyllifolia L. subsp. serpyllifolia Plantaginaceae + [325 m]
Vicia peregrina L. Fabaceae + + [485 m]
*Wisteria sinensis (Sims) Sweet Fabaceae C + + + [245 m]
*Yucca aloifolia L. Asparagaceae C + + + [115 m]
*Ziziphus jujuba Mill. Rhamnaceae C +
Entre les espècies autòctones cal destacar la localització d’alguns tàxons amb àrees de distribució poc extenses a Catalunya, com ara Trifolium ligusticum (que es circumscriu a l’extrem nord-oriental del país), Lotus angustissimus subsp. suaveolens (citat principalment en àrees del litoral de la meitat oriental) o l’híbrid Equisetum × moorei, de distribució mal coneguda, que ja havia estat indicat en diferents punts de la comarca de la Selva (Vilar, 1987).
Pel que fa a les plantes aŀlòctones, una gran majoria són espècies d’ús ornamental, que hom pot veure cultivades als jardins i zones verdes de les extenses urbanitzacions del municipi, i que apareixen també fora de cultiu en espais antropitzats propers (vores de carrers i de camins, solars, descampats, talussos, herbassars, etc.). Cal advertir que una part important d’aquestes plantes introduïdes no semblen plenament establertes al territori, sinó que hi tenen un comporta-
ment casual (Blackburn et al., 2011, Pyšek et al., 2004; Richardson et al., 2000, 2011), de manera que el manteniment de les seves poblacions depèn de la introducció repetida de propàguls des dels punts on són cultivades. Molts d’aquests tàxons aŀlòctons són plantes àmpliament distribuïdes a Catalunya, on fa temps que hi són conegudes. Això no obstant, també n’hi ha alguns d’introducció més recent o que han estat poc reportats al territori català. Aquest és el cas d’Epilobium brachycarpum, documentat a la Terra Alta (Cardero et al., 2004), a l’Alt Urgell (Aymerich, 2013, 2016b) i a la Selva (Gesti & Vilar, 2019); d’Oenothera oehlkersi (tàxon d’origen probablement híbrid que alguns autors inclouen dins d’Oentohera glazioviana Micheli), que s’ha citat a la Garrotxa (Verloove & Sánchez Gullón, 2008), al Ripollès (Aymerich, 2016a), al Pallars Sobirà (Aymerich, 2017) i a la província de Barcelona (Dietrich, 1997); d’ Osteospermum ecklonis ,
202 Butlletí ICHN 85 (4), 2021
P. BARNOLA ECHENIQUE ET AL.
APORTACIONS AL CONEIXEMENT DE LA FLORA VASCULAR DEL SECTOR ORIENTAL DEL MASSÍS DEL MONTSENY
indicada en alguns punts del litoral i prelitoral (C. Gómez & H. Álvarez in BC; Gesti, 2020); i de Silene pseudoatocion i Tagetes patula, ambdues àmpliament indicades a l’extrem meridional del país però, en canvi, escassament reportades a l’àrea nordoriental (vegeu distribució a Font, 2021).
Taula 2. Noves aportacions a la flora vascular del massís del Montseny.
Aquests nous 43 taxons suposen un increment d’un 2,6 % de tàxons en relació a les dades disponibles anteriorment al Montseny (Taula 2). En termes relatius, l’aportació més rellevant (12,9 %) correspon a les plantes aŀlòctones, que passen dels 232 tàxons citats prèviament als 262 coneguts actualment.
Montseny Dades bibliogràfiques i d’herbaris
Tàxons no indicats prèviament al sector oriental del massís del Montseny
A banda dels 43 tàxons que ja hem indicat com a nous pel Montseny (i que, per tant, també ho són pel sector est), n’aportem 29 més (15 autòctons i 14 aŀlòctons) que ja eren coneguts al massís però no del seu sector oriental (Taula 1, columna «Est»).
En molts casos es tracta d’espècies que són força rares al conjunt del Montseny, ja siguin autòctones com ara Allium ampeloprasum, Aristolochia paucinervis, Brachypodium
Taula 3. Noves aportacions
Tàxons no indicats prèviament a la zona basal del massís del Montseny
A la Taula 1 (columna «Bas») llistem 76 tàxons (38 autòctons i 38 aŀlòctons) que hem pogut observar a l’àrea basal del Montseny (fins als 500 m d’altitud) i que no hi havien estat citats. Hi són bona part dels tàxons que hem donat com a novetats pel Montseny i també tot un seguit de plantes citades del massís, però que no havien estat reportades per sota dels 500 m. Algunes són plantes amb el seu òptim a terra baixa o que estan vinculades a ambients que són més freqüents en la zona basal (conreus, indrets antropitzats, zones humides de plana, etc.): Bolboschoenus maritimus, Bromus lanceolatus,
distachyon, Campanula erinus, Cirsium monspessulanum, Scolymus hispanicus, Scorzonera angustifolia, Urospermum picroides o Vicia peregrina, o bé aŀlòctones com Eschscholzia californica, Euphorbia serpens, Ligustrum lucidum, Physalis ixocarpa, Phyla canescens, etc.
En conjunt, aquests 72 tàxons suposen un augment d’un 5,4 % en relació als coneguts anteriorment en aquest sector (Taula 3). De nou, és en el grup de les plantes aŀlòctones on hi ha hagut una aportació relativa més rellevant (22,6 %), tot passant dels 195 tàxons citats prèviament als 239 actuals.
Equisetum ramosissimum, Eragrostis minor, Heteropogon contortus, Iris pseudacorus, Lupinus angustifolius, Veronica hederifolia, etc. D’altres, en canvi, són espècies més pròpies de la muntanya mitjana que arriben a cotes baixes aprofitant els ambients que els són favorables (obagues, riberes fluvials, fondals...): Avenella flexuosa, Cruciata glabra, Myosotis sylvatica subsp. teresiana, Pseudoturritis turrita, Serratula tinctoria, Tilia platyphyllos, Veronica serpyllifolia, etc.
En fan part també un gran nombre de plantes aŀlòctones casuals que tenen el seu origen en els conreus (Brassica napus, Prunus domestica, Punica granatum, Pyrus communis, etc.) o, més sovint, en les abundants zones ajardinades de
203 Butlletí ICHN 85 (4), 2021
FLORA ET FAUNA
GEA,
Noves aportacions Increment Total Nombre de tàxons autòctons 1.413 13 0,92 % 1.426 Nombre de tàxons aŀlòctons 232 30 12,93 % 262 Nombre total de tàxons 1.645 43 2,61 % 1.688
Sector Est Dades bibliogràfiques i d’herbaris Noves aportacions Increment Total Nombre de tàxons autòctons 1.141 28 2,45 % 1.169 Nombre de tàxons aŀlòctons 195 44 22,56 % 239 Nombre total de tàxons 1.336 72 5,39 % 1.408
a la flora vascular del sector oriental del massís del Montseny.
Àrea basal Dades bibliogràfiques i d’herbaris Noves aportacions Increment Total <250 m Nombre de tàxons autòctons 717 27 3,77 % 744 Nombre de tàxons aŀlòctons 154 38 24,68 % 192 Nombre total de tàxons 871 65 7,46 % 936 250-500 m Nombre de tàxons autòctons 996 23 2,31 % 1.019 Nombre de tàxons aŀlòctons 152 6 3,95 % 158 Nombre total de tàxons 1.148 29 2,53 % 1.177
Taula 4. Noves aportacions a la flora vascular de l’àrea basal del massís del Montseny.
GEA, FLORA ET FAUNA
les urbanitzacions de Riells i Viabrea (Berberis aquifolium, Campsis radicans, Passiflora caerulea, etc.).
Prenent en consideració els mateixos dos intervals altitudinals que s’analitzaven al catàleg de 2017, es pot observar com és al de cotes per sota de 250 m on han aparegut més novetats, amb 65 nous tàxons (que suposen un increment del 7,5 % en relació als coneguts prèviament). També és l’interval que concentra la major part de les noves espècies aŀlòctones identificades a l’àrea basal (38 tàxons, +24,7 %).
Pel que fa a l’interval comprès entre els 250 i els 500 m, s’hi han localitzat un nombre més petit de novetats (29 tàxons, +2,5 %) i l’increment d’espècies aŀlòctones és menys acusat (6 tàxons, +4,0 %).
Agraïments
Volem agrair a Carlos Gómez haver-nos facilitat la clau de determinació del gènere Oenothera que prepara per la seva tesi doctoral i haver atès diversos dubtes sobre espècies aŀlòctones. També a Roser Guàrdia i Josep Vicens (BCN), a Neus Ibáñez (BC) i a Lluís Vilar (HGI) per haver-nos facilitat la consulta i els llistats dels plecs d’herbari de les respectives coŀleccions.
Bibliografia
Aymerich, P. 2013. Sobre algunes espècies aŀlòctones a l’alt Segre (NE de la península Ibèrica), noves o molt rares per a la flora catalana. Orsis, 27: 195-207.
Aymerich, P. 2016a. Contribució al coneixement de la flora aŀlòctona del nord i el centre de Catalunya. Orsis, 30: 11-40.
Aymerich, P. 2016b. Notes florístiques de les conques altes dels rius Segre i Llobregat. IV. Orsis, 30: 133-165.
Aymerich, P. 2017. Notes sobre flora aŀlòctona a Catalunya. Butlletí de la Institució Catalana d’Història Natural, 81: 97-116.
Barnola, P. & Romo, À. M. 1989. Addicions a la flora vascular del Montseny. II Trobada d’Estudiosos del Montseny. Monografies de la Diputació de Barcelona, 18: 107-108.
Barnola, P., Vilar, L. & Pié, G. 2014. Aportacions a la flora vascular del Baix Montseny. Aportacions a la flora vascular. VIII Trobada d’Estudiosos del Montseny. Diputació de Barcelona. Monografies del Montseny, VIII: 119-122.
Blackburn, T. M., Pyšek, P., Bacher, S., Carlton, J. T., Duncan, R.P., Jarošík, V., Wilson, J. R. & Richardson, D. M. 2011. A proposed unified framework for biological invasions. Trends in Ecology & Evolution, 26 (7): 333–339.
Bolòs, O. de, Nuet, J. & Panareda, J. M. 1986. Flora vascular del Montseny. P. 41-92. In: Terrades, J. & Miralles, J. (eds.). El patrimoni biològic del Montseny. Catàlegs de flora i fauna, 1. Diputació de Barcelona. Barcelona.
Bolòs, O. de. 1998. Atlas corològic de la flora vascular dels Països Catalans: Primera compilació general. Barcelona: Institut d’Estudis Catalans. Secció de Ciències Biològiques.
Cardero, S., Royo, F. & Torres, L. 2004. Novetats florístiques per a les Terres de l’Ebre. Butlletí de la Institució Catalana d’Història Natural, 72: 99-103.
Dietrich, W. 1997. Oenothera L. P. 90-100. In: Castroviejo, S., Aedo, C., Benedí, C., Laínz, M., Munoz Garmendia, F., Nieto; G. & Paiva, J. (eds.). Flora iberica, Vol. VIII. HaloragaceaeEuphorbiaceae. Real Jardín Botánico-CSIC. Madrid.
Font, X. 2021. Mòdul de Flora i Vegetació. Banc de Dades de Biodiversitat de Catalunya. Generalitat de Catalunya i Universitat
de Barcelona. Disponible a: http://biodiver.bio.ub.es/biocat/ homepage.html [Data de consulta: 13 setembre 2021].
Gesti, J. & Vilar, L. 2019. Aportacions al coneixement de la flora de les Guilleries orientals i àrees properes (nord-est de Catalunya). Butlletí de la Institució Catalana d’Història Natural, 83: 195-198.
Gesti, J. 2020. Catàleg de la flora vascular de Santa Coloma de Farners (la Selva, nord-est de Catalunya). Miconia, 4: 69-105.
González, V. & Nuet, J. 2020. Projecte FitoCAT, 40: Plantes interessants de Sant Llop de Viabrea (la Selva). Miconia, 4: 295-314.
Gutiérrez, C. & Sáez, L. 1996. Aportacions al coneixement de la flora vascular del Montseny. Folia Botanica Miscellania, 10: 67-75.
Gutiérrez, C. 1998. Novetats florístiques per al Montseny i el Montnegre. Butlletí de la Institució Catalana d’Història Natural, 66: 59-62.
Gutiérrez, C. 1999. Novetats a la flora vascular del Montseny. III i IV Trobades d’Estudiosos del Montseny. Diputació de Barcelona. Monografies, 27: 23-25.
Llensa, S. 1945. Inventario razonado de la Flora de Hostalrich y su comarca. Anales de la Escuela de Peritos Agrícolas y Superior de Agricultura y de los Servicios Técnicos de Agricultura, 5: 121-290.
Mercadal, G. 2019. Els prats de dall de la terra baixa catalana. Caracterització geobotànica, valoració agroambiental i estudi de les relacions fitosociològiques entre els prats dalladors de l’Europa occidental. Tesi Doctoral. Universitat de Girona. Girona. 2.306 p.
Mercadé, A. & Pérez-Haase, A. 2013. Aportació al coneixement florístic del Montseny. Butlletí de la Institució Catalana d’Història Natural, 77: 148-152.
Nuet, J. 2015. Notes sobre la flora del Montseny (Catalunya), 1. Butlletí de la Institució Catalana d’Història Natural, 79: 55-57.
ORCA. 1985-2014. Atlas corològic de la flora vascular dels Països Catalans. Institut d’Estudis Catalans. Secció de Ciències Biològiques. Barcelona.
Panareda, J. M. & Nuet, J. 2005. Novetats florístiques al Montseny des del 1986. VI Trobada d’Estudiosos del Montseny. Diputació de Barcelona. Monografies, 41: 63-66.
Pié, G. & Vilar, L. 2014. Corologia de la flora vascular d’interès de conservació al Parc Natural del Montseny. Butlletí de la Institució Catalana d’Història Natural, 78: 65-74.
Pié, G., Vilar, L. & Barnola, P. 2014. Estudis corològics al Parc Natural del Montseny. Aportacions a la flora vascular. VIII Trobada d’Estudiosos del Montseny. Monografies del Montseny, VIII: 114118.
Pyšek, P., Richardson, D. M., Rejmánek, M., Webster, G. L., Williamson, M. & Kirschner, J. 2004. Alien plants in checklists and floras: towards better communication between taxonomists and ecologists. Taxon, 53 (1): 131–143.
Richardson, D. M., Pyšek, P., Rejmanek, M., Barbour, M. G., Panetta, F. D. & West, C. J. 2000. Naturalization and invasion of alien plants: concepts and definitions. Diversity Distributions, 6 (2): 93–107.
Richardson, D. M., Pyšek P. & Carlton J. T. 2011. A compendium of essential concepts and terminology in biological invasions. P. 409420. In: Richardson, D. M. (ed.). Fifty years of invasion ecology: the legacy of Charles Elton. Blackwell Publishing. Oxford.
Sáez, L., Galán de Mera, A., Pyke, S., Pié, G. & Carnicero, P. 2015. New data on vascular plants from Montseny massif (northeastern Iberian Peninsula). Orsis, 29: 205-230.
Sáez, L., Pié, G. & Carnicero, P. 2017. Catàleg de la flora vascular del massís del Montseny. Tres segles d’investigació botànica (1716-2016). Sèrie Territori i Parcs Naturals, 1. Coŀlecció Estudis. Diputació de Barcelona. 238 p.
Verloove, F. & Sánchez Gullón, E. 2008. New records of interesting xenophytes in the Iberian Peninsula. Acta Botanica Malacitana, 33: 147-67.
Vilar, L. 1987. Flora i vegetació de la Selva. Tesi doctoral. Universitat Autònoma de Barcelona. Bellaterra. 608 p.
204 Butlletí ICHN 85 (4), 2021
P. BARNOLA ECHENIQUE ET AL.
APORTACIONS
Apèndix. Localitats (totes corresponents a Riells i Viabrea) dels tàxons citats al text. Els noms dels tàxons al·lòctons van precedits d’un asterisc (*).
Agrimonia eupatoria subsp. grandis (Andrz. ex Asch. & Graebn.) Bornm.: a can Perearnau, DG5825, 540 m, herbassar en un antic camp abandonat (HGI 24247, Barnola, Panareda & Gesti, 5-VII-2020).
Agrostis capillaris subsp. capillaris: prop can Mora, DG6224, 250 m, herbassar, 29-V-1999; vora riera de Riells, DG6123, 270 m, herbassar, 10-III-2012; entorn de can Salvà, DG6319, 220 m, herbei, 20-III-2013; prop de Roca Negra, DG5923, 450 m, herbassar vora pista, 31-XII-2019; prop de can Plana, DG5922, 310 m, herbassar, 21-VI-2020.
*Alcea rosea L.: al sot Gran (urbanització Bosc de la Batllòria), DG6119, 110 m, herbassar ruderal, P. Alboquers, 15-VI-2020.
Allium ampeloprasum L.: sobre la riera de Sant Llop, DG6220, 140 m, herbassar al marge del camí, 13-VIII-2020.
Aristolochia paucinervis Pomel: Boscos de can Plana, DG5922, 350 m, vorada forestal, 21-VI-2020.
Avenella flexuosa (L.) Drejer: sobre can Merlet, DG5925, 485 m, talús de pista forestal, 5-VII-2020.
*Bassia scoparia (L.) A.J. Scott: c. la Rajolaria, DG6319, 95 m, marge de conreu, 13-X-2010.
*Berberis aquifolium Pursh: carrer de les Aigües, DG6319, 90 m, talús, 16-II-2020; a l’avinguda de la Moreneta, DG6219, 130 m, marge, 26-II-2020; Bosc de la Batlloria, DG6219, 135 m, sot humit, 26-II-2020; a can Pujol, DG6021, 240 m, marge, 8-IX-2020.
Bolboschoenus maritimus (L.) Palla: marge de la riera de Riells, DG6319, 95 m, indret enclotat, 9-VIII-1999.
Brachypodium distachyon (L.) P. Beauv.: entorn de can Salvà, DG6319, 110 m, clariana de bosc, 3-VI-2020; sobre can Pagès, DG6320, 140 m, clariana bosc, 3-VI-2020; carretera de Sant Llop, c. can Pla, DG6221, 215 m, vorada, 3-VI-2020; entre can Riboies i can Marxant, DG6320, 150 m, prat sec, 3-VI-2020; sobre can Planes, DG5921, 335 m, prat sec en una vorada de pista forestal, 21-VI-2020.
*Brassica napus L.: sota can Riboies, DG6320, 95 m, marge de conreu, 16-II-2020.
Bromus lanceolatus Roth: a l’urbanització Bosc de la Batllòria, DG6219, 100 m, descampat, 27-VIII-2020; a can Salvà, DG6319, 125 m, parcel·la no urbanitzada (BCN 163135, Barnola & Gesti, 10-VII-2020).
Campanula erinus L.: a la Torre de Montfort, DG5926, 730 m, pradell terofític, 8-VIII-2020.
*Campsis radicans (L.) Seem.: a la urbanització can Salvà, DG6319, 95 m, vorada entre el carrer i el bosc, 27-VIII-2020; a can Pujol, DG6021, 240 m, herbassar ruderal, 8-IX-2020.
*Carpobrotus edulis (L.) N. E. Br.: a can Pujol, DG6021, 240 m, talús, 8-IX-2020.
*Cedrus deodara (Lamb.) G. Don.: al sot de Rocablanca, DG6022, 250 m, vorada de pista forestal, 8-IX-2020.
Centaurium pulchellum (Swartz) Druce: c. can Mora, DG6224, 250 m, herbassar, 29-V-1999; sobre can Plana, DG5921, 335 m, vorada de pista forestal, 21-VI-2020.
*Cerastium tomentosum L.: Fogueres de Montsoriu, DG6025, 625 m, herbei en un talús, 10-VII-2021.
*Cercis siliquastrum L.: a can Salvà, DG6319, 110 m, marge de camp, 13-VIII-2020.
Cirsium monspessulanum (L.) Hill: al torrent dels Vimeners, DG5625, 985 m, herbassar molt humit de ribera, 16-VII-2020; al torrent dels Dos Sotets, c. els Vimeners, DG5626, 990 m, herbassar humit, 16-VII-2020.
*Cotula australis (Spreng.) Hook. f.: a Fogueres de Montsoriu, DG6025, 625 m, prat en zona verda, 26-III-2021.
Cruciata glabra (L.) Ehrend.: riera de Riells, c. ca n’Iscle, DG6023, 300 m, prat humit, 9-IV-1999; c. cal Bord Nou, DG6124, 350 m, bosc de fondalada, 9-IV-1999; c. can Manuel, DG6024, 380 m, herbassar, 9-IV-1999; c. can Mora, DG6224, 250 m, herbassar, 29-V-1999; sobre can Pelegrí, DG5925, 490 m, vorada forestal, 5-VII-2021.
*Cyperus alternifolius subsp. flabelliformis (Rottb.) Kük.: al Bosc de la Batllòria, DG6219, 125 m, torrent, 26-II-2020.
*Dichondra micrantha Urb.: a Viabrea, c. via de ferrocarril, DG6218, 95 m, prat en una zona verda, 3-X-2020; a can Bassas, DG6221, 150 m, prat calcigat, 5-IX-2020; c. estació de ferrocarril, DG6319, 85 m, descampat, 3-X-2020; Bosc de la Batllòria, DG6219, 130 m, herbassar ruderal, 9-V-2021.
*Echinochloa colonum (L.) Link: a cal Gall, DG6221, 140 m, camp abandonat, 5-IX-2020; a Viabrea, entre la carretera C-35 i les vies del ferrocarril, DG6218, 95 m, camp sense cultivar, 12-IX-2020; a ca n’Hoste, DG6320, 120 m, camp llaurat, 12-IX-2020; a l’aparcament de l’estació de ferrocarril de Viabrea, DG6419, 85 m, herbassar ruderal, 26-IX-2020; al polígon industrial de Viabrea, DG6219, 95 m, descampat, 3-X-2020; deixalleria de Viabrea, DG6319, 90 m, herbassar ruderal humit, 3-X-2020.
Elymus caninus (L.) L.: sota can Salvà, DG6319, 95 m, marge, 16-II-2020; sota can Riboies, DG6320, 95 m, marge, 16-II-2020.
Elymus pungens (Pers.) Melderis subsp. campestris (Godr. & Gren.) Melderis: a ca n’Iscle, DG6123, 310 m, marge de camp, 21-XII-2019; sota can Riboies, DG6320, 120 m, marge, 16-II-2020.
*Epilobium brachycarpum C. Presl: vora la riera de Sant Llop, DG6319, 85 m, runam, 16-IX-2019.
Epilobium tetragonum L.: sot sota el carrer del Xaloc (Residencial Riells II), DG6221, 175 m, vorada forestal, 9-VI-2020.
Equisetum × moorei Newman: al sot de ca n’Amat, DG5824, 560 m, verneda, 13-III-2021; riera de Breda, DG6122, 220 m, verneda, 5-IX2020.
Equisetum ramosissimum Desf.: vora l’estació de ferrocarril de Viabrea, DG6419, 85 m, talús, 16-II-2020; Bosc de Batllòria, DG6219, 100 m, talús, 26-II-2020; torrent de can Berenguer, c. can Ganyes, DG6120, 155 m, vorada de pista forestal, 28-V-2020; a can Castells, DG6220, 105 m, vorada de camí, 13-VIII-2020; c. can Plana, DG5922, 295 m, marge d’herbassar humit, 28-X-2020.
Eragrostis minor Host: carril bici de la carretera de Breda, DG6319, 95 m, herbassar calcigat, 12-IX-2020; aparcament de l’estació de ferrocarril de Viabrea, DG6419, 85 m, herbassar ruderal, 26-IX-2020.
*Eriobotrya japonica (Thunb.) Lindl.: vora riera del sot Gran, DG6119, bardissa, 100 m, 26-II-2020; urbanització de can Plana, DG6220, clariana forestal, 125 m, 26-II-2020; a can Merlet, DG5925, vora de camí, 490 m, 5-VII-2020; sobre el torrent de can Dansa, DG6118, talús sobre el torrent, 100 m, 5-VII-2020; a can Bassas, DG6221, 150 m, vora de camí, 5-IX-2020.
205 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA
AL CONEIXEMENT DE LA FLORA VASCULAR DEL SECTOR ORIENTAL DEL MASSÍS DEL MONTSENY
GEA, FLORA ET FAUNA
*Eschscholzia californica Cham.: entre els carrers de Mestral i de Xaloc (Residencial Riells II), DG6221, 185 m, herbassar ruderal, 9-VI2020.
*Eucalyptus camaldulensis Dehnh.: sota can Guillem, DG6120, 175 m, vorada forestal, 26-IX-2020.
*Eucalyptus globulus Labill.: c. riera de Riells, sota ca n’Iscle, DG6023, 310 m, clariana de bosc, 9-IV-1999; vora can Bord Vell, DG6124, 350 m, vora camí, 9-IV-1999; prop can Mora, DG6224, 250 m, vora masia, 29-V-1999; vora la riera de Riells, DG6123, 270 m, marge d’antic camp, 10-III-2012; Residencial Riells II, DG6221, 210 m, marge, 20-IX-2012.
*Euphorbia serpens Kunth: carrer de les Aigües, DG6319, 90 m, vora de camí (BCN 161746, Panareda & Barnola, 13-VIII-2020); polígon industrial de Viabrea, DG6219, 95 m, descampat, 3-X-2020.
*Gazania rigens (L.) Gaertn.: a Fogueres de Montsoriu, DG6025, 595 m, solar (BCN 163128, Barnola & Gesti, 10-VII-2021).
Glebionis coronaria (L.) Cass. ex Spach: sobre el torrent de can Dansa, DG6119, 120 m, herbassar ruderal humit, 8-V-2021.
Heteropogon contortus (L.) P. Beauv. ex Roem. & Schult.: talús vora cal Gall, DG6221, 155 m, prat sec, 5-IX-2020.
Iris pseudacorus L.: riera de Breda, c. can Marc, DG6320, 100 m, herbassar de ribera, 17-IV-2021.
*Lamium galeobdolon subsp. argentatum (Smejkal) J. Duvig.: a can Bassas, DG6221, 145 m, herbassar, 9-III-2021.
Lathyrus niger (L.) Bernh.: c. can Tordera, DG6024, 375 m, vorada forestal, 28-IV-2018.
*Ligustrum lucidum W.T. Aiton: avinguda de la Moreneta (Bosc de la Batllòria), DG6219, 115 m, talús, 26-II-2020; fondal c. can Guilleumes, DG6221, 140 m, vorada forestal, 5-IX-2020; Alba de Liste, DG6218, 100 m, marge de camp, 12-IX-2020; Viabrea, c. carretera de Breda, DG6319, 85 m, vorada de camí, 26-IX-2020; torrent de can Dansa, DG6019, 150 m, talús, 25-II-2021.
Lotus angustissimus subsp. suaveolens (Pers.) O. Bolòs & Vigo: sobre can Plana, DG5922, 320 m, pradell terofític humits en una vorada de pista forestal (HGI 24247, Barnola, Panareda & Gesti, 21-VI-2020).
Lupinus angustifolius L.: sota can Riboies, DG6320, 120 m, clariana bosc, 20-II-2016; c. can Joanic, DG5924, 475 m, vorada de pista forestal, 31-XII-2019; al pont de Sota Via (Viabrea), DG6419, 80 m, marge de camp, 16-II-2020; a can Pere Llop, DG5823, 560 m, herbassar obert, 13-III-2021; a can Bernat, DG5726, 785 m, clariana forestal, 23-V-2021.
*Malva arborea (L.) Webb & Berthel.: Sant Martí de Riells, DG5925, 465 m, herbassar ruderal, 31-XII-2019; c. ca n’Iscle, DG6123, 315 m, talús, 12-V-2021.
*Melia azedarach L.: Bosc de la Batllòria, DG6219, 105 m, vorada forestal, 13-XII-2020.
Melilotus officinalis (L.) Lam.: sot dels Roures, DG6122, 240 m, vora de camí ombrívol, 29-V-2010; torrent de can Dansa, DG6118, 125 m, herbassar humit, 25-II-2021; can Tries, DG6021, 255 m, prat humit, 2-IV-2021.
*Morus alba L.: Bosc de la Batllòria, DG6219, 105 m, clariana forestal, 26-II-2020; turó sobre la font de ca n’Erola, DG6222, 95 m, camp abandonat, 12-IX-2020; a Viabrea, DG6319, 85 m, descampat, 26-IX-2020; sobre el torrent de can Dansa, DG6118, 100 m, talús, 25-II2021; a Junior Park, DG6119, 160 m, vorada forestal, 8-V-2021.
Myosotis sylvatica subsp. teresiana (Senn.) O. Bolòs & Vigo: Riera de Riells, DG6024, 380 m, fondal de la riera, 9-IV-1999; sot de Can Pere Roig, DG5924, 450 m, fondal del sot, 9-IV-1999.
Narcissus tazetta L.: a can Salvà, DG6319, 110 m, herbassar en un talús, 16-II-2020.
*Nigella damascena L.: urbanització can Plana, DG6220, 110 m, marge de camí, 12-VII-2021.
*Nothoscordum gracile (Aiton) Stearn [incl. N. × borbonicum Kunth]: a Residencial Riells II, DG6220, 120 m, herbassar, N. Julià, 30-IV2021; Bosc de la Batllòria, DG6219, 115 m, herbassar, 9-V-2021.
*Oenothera oehlkersi Kappus: marge de la carretera GI552, c. ca n’Hoste, DG6320, 120 m, herbassar humit (BCN 163131, Barnola, 6-VII-2021).
Onopordum acanthium L.: sota can Salvà, 85 m, DG6319, camp abandonat, 19-IX-2016; al camí de can Perearnau, DG5925, 505 m, herbassar en un camp abandonat, 5-VII-2020; al camí de can Perearnau, DG5825, 515 m, camp abandonat, 5-VII-2020.
*Osteospermum ecklonis (DC.) Norl.: al Bosc de la Batllòria, DG6219, 105 m, herbassar al marge del carrer, 9-V-2021.
Oxalis acetosella L.: riera de Riells, DG5925, 500 m, verneda, 9-IV-1999.
*Panicum capillare L.: entre la carretera GIV-5523 i la riera de Breda, c. estació de tren, DG6419, 80 m, herbassar ruderal humit, 26-IX2020.
Papaver somniferum subsp. setigerum (DC.) Arcang.: vora de la riera de Sant Llop, DG6319, 85 m, talús de la via del tren, 16-IX-2019.
*Passiflora caerulea L.: pla de Saba, DG5824, 665 m, bardissa, 31-XII-2019; a la urbanització de can Plana, DG6219, 105 m, herbassar en un solar, 27-VIII-2020; a Residencial Riells II, DG6221, 160 m, vorada forestal, 5-IX-2020; sobre el torrent de can Dansa, a tocar de Royal Park, DG6019, 135 m, vorada forestal, 25-II-2021; can Salvà, DG6319, 95 m, solar, 10-VII-2021.
Phalaris paradoxa L.: prop Molí d’en Hoste, DG6320, 120 m, herbassar ribera riera de Riells, 5-VII-2020.
*Phyla canescens (Kunth) Greene: camí c. can Masó, DG6220, 135 m, vorada de camí, 13-VIII-2020; polígon industrial de Viabrea, DG6319, 95 m, descampat, 3-X-2020; vora les piscines (Bosc de la Batlloria), DG6219, 105 m, herbassar (BCN 163130, Barnola, 11VII-2021).
*Physalis ixocarpa Brot. ex Hornem.: al costat del camp de futbol de Viabrea, DG6319, 115 m, camp de conreu, 13-VIII-2020; carrer de les Aigües, DG6319, 90 m, camp de conreu llaurat (BCN 161744, Panareda & Barnola, 27-VIII-2020); a ca n’Hoste, DG6320, 115 m, camp de conreu llaurat, 12-IX-2020.
*Pittosporum tobira (Thunb.) W.T. Aiton: Junior Parc, DG6120, 150 m, marge vora de camí, 29-XII-2012; c. via del ferrocarril, DG6419, 85 m, talús, 16-II-2020; al Bosc de la Batllòria, DG6219, 130 m, talús en un fondal, 26-II-2020; molí d’en Bosc, DG6222, 210 m, bosc de ribera, 5-IX-2020.
Plantago major subsp. intermedia (Gilib.) Lange: sota can Fortuny de Baixt, DG6024, 380 m, clariana vial inundada periòdicament, 9-IV1999.
206 Butlletí ICHN 85 (4), 2021
P. BARNOLA ECHENIQUE ET AL.
APORTACIONS
GEA, FLORA ET FAUNA
*Plumbago auriculata Lam.: urbanització Boscos del Montseny, DG6219, 125 m, marge del carrer (BCN 163133, Barnola, 2-XI-2020).
*Populus deltoides Marshall: al Molí d’en Bosc, DG6222, 220 m, fondal humit, 5-IX-2020
*Prunus domestica L.: entre la riera de Riells i Ca n’Iscle, DG6023, 300 m, vora de camí, 9-IV-1999; sota can Fortuny de Baix, DG6024, 380 m, marge de camp, 9-IV-1999; sot de can Pere Roig, DG5924, 450 m, marge d’herbassar, 9-VI-1999; al Moli de n’Hoste, DG6320, 105 m, marge de camp, 12-IX-2020.
*Prunus dulcis (Mill.) D.A. Webb: a can Marxant, DG6320, 165 m, marge de camp, 5-V-2021; al torrent de can Dansa, DG6218, 105 m, vorada forestal, 8-V-2021; al Bosc de la Batllòria, DG6219, 145 m, marge entre el carrer i el sot, 9-V-2021.
Pseudoturritis turrita (L.) Al-Shehbaz: c. Sant Martí de Riells, DG5925, 460 m, marge de camí ombrívol, 31-XII-2019; al sot de ca n’Amat, c. can Tordera, DG6024, 330 m, herbassar humit, 31-XII-2019.
*Punica granatum L.: a can Pujol, DG6021, 255 m, marge, 8-IX-2020; a can Marxant, DG6320, 165 m, fenassar, 5-V-2021.
*Pyracantha fortuneana (Maxim.) H.L. Li: Bosc de la Batllòria, DG6219, 100 m, bardissa, 27-VIII-2020; sobre el torrent de can Dansa, c. Royal Park, DG6119, 140 m, pista forestal, 8-V-2021.
*Pyrus communis L.: entorn de can Salvà, DG6319, 95 m, alzinar esclarissat, 9-VIII-1999; a can Ganyes, DG6121, 175 m, bardissa, 28-V2020; can Tries, DG6021, 250 m, bardissa, 28-V-2020; al costat del camp de futbol de Viabrea, DG6319, 110 m, descampat, 13-VIII2020; a can Plana, DG5922, 305 m, marge de camí, 2-IV-2021.
Ranunculus sardous subsp. trilobus (Desf.) Rouy & Fouc.: a can Plana, DG5922, 310 m, prat humit, 13-III-2021; sobre el torrent de can Dansa, DG6118, 100 m, herbasar humit, 5-V-2021.
Reseda luteola L. subsp. luteola: vora la riera de Sant Llop, DG6319, 85 m, marge, 16-IX-2019.
Rosa pouzinii Tratt.: sota el coll de Castellets, DG6125, 360 m, bardissa en fondal, 21-X-2017.
Scolymus hispanicus L.: carrer de la Cadernera, DG6319, 115 m, herbassar ruderal en un talús, 3-VII-2015; a Can Salvà, DG6319, 125 m, herbassar ruderal, 10-VII-2021.
Scorzonera angustifolia L.: c. Sant Llop de Viabrea, DG6122, 240 m, brolla (BCN 161750, Panareda & Barnola, 9-VI-2020).
Serratula tinctoria L.: sobre el sot de Rocablanca, DG5922, 405 m, perxada de castanyers, 21-VI-2020; sobre la riera de Sant Llop, c. can Ganyes, DG6121, 200 m, clariana forestal, 10-IX-2020; al sot dels Roures, DG6222, 175 m, vorada forestal, 12-IX-2020.
Setaria adhaerens (Forssk.) Chiov.: c. la Batllòria, entre la carretera C-35 i les vies del ferrocarril, DG6218, 95 m, camp sense cultivar (HGI 24057, Gesti, Panareda & Barnola, 12-IX-2020); entre la riera de Sant Llop i el nucli de Viabrea, DG6319, 85 m, camp abandonat, 26-IX-2020.
*Silene pseudoatocion Desf.: urbanització can Salvà, DG6319, 105 m, herbassar ruderal en un descampat, 16-II-2020.
*Sinapis arvensis L.: prop Molí de n’Hoste, DG6320, 110 m, conreu, 20-II-2016; sobre l’estació depuradora, DG6319, 115 m, talús entre carrers, 16-II-2020; Bosc de la Batllòria, DG6219, 120 m, herbassar ruderal al marge del carrer, 9-V-2021.
*Solanum tuberosum L.: Sant Martí de Riells, DG5925, 475 m, herbassar ruderal, 31-XII-2019.
Sparganium erectum L.: a la riera de Breda, c. can Quel, DG6320, 100 m, herbassar de ribera, 12-IX-2020.
*Tagetes patula L.: a Viabrea, entre la carretera C-35 i les vies del ferrocarril, DG6218, 95 m, camp sense cultivar, 12-IX-2020.
Tilia platyphyllos Scop.: Sant Martí de Riells, DG5925, 480 m, bosc mixt de caducifolis, 5-VII-2020; al Molí d’en Bosc, DG6222, 215 m, vorada forestal, 5-IX-2020.
Torilis arvensis (Huds.) Link subsp. arvensis: entorn de can Salvà, DG6319, 90 m, herbassar, 9-VIII-1999; c. can Plana, DG5922, 295 m, marge de bosc, 21-VI-2020; c. Molí de n’Hoste, DG6320, 120 herbassar, 5-VII-2020; a can Marlet, DG5925, 485 m, herbassar, 5-VII2020; sot de Rocablanca, DG6022, 220 m, vora pista, 5-IX-2020.
*Trachycarpus fortunei (Hook.) H. Wendl.: riera de Breda, DG6419, 85 m, bosc de ribera, 16-II-2020.
*Tradescantia fluminensis Velloso: sota l’Hostal Bell-lloc, DG5925, 465 m, talús humit, 31-XII-2019; bosc de la Batllòria, DG6219, 125 m, talús del torrent, 26-II-2020.
Tragopogon porrifolius L.: al torrent de can Dansa, DG6119, 110 m, herbassar humit, 8-V-2021.
Trifolium ligusticum Balb. ex Loisel.: c. Sant Llop de Viabrea, DG6122 i DG6122, 240 m, clariana vora pista, 9-VI-2020.
*Ulmus pumila L.: a Alba de Liste, c. carretera a Sant Celoni, DG6218, 95 m, vorada, 3-X-2020.
Urospermum picroides (L.) Scop. ex F.W. Schmidt: sobre el torrent de can Berenguer, DG6021, 205 m, marge del camí, 27-V-2020; sobre can Plana, DG5921, 325 m, vora de pista forestal, 21-VI-2020; Bosc de la Batlloria, DG6219, 125 m, marge del carrer, 9-V-2021; a ca n’Iscle, DG6123, 315 m, fenassar, 12-V-2021; a Fogueres de Monsoriu, DG6025, 630 m, herbassar, 26-III-2021.
Veronica hederifolia L.: les Oliveres, DG5924, 450 m, ran de camí, 9-IV-1999; a can Merla, DG6420, 90 m, camp de conreu, 16-II-2020; a tocar de les vies del ferrocarril de Viabrea, DG6319, 90 m, herbassar ruderal, 1-III-2020; a Royal Park, DG6019, 145 m, marge de camí, 25-II-2021; a can Riboies, DG6320, 150 m, camp de conreu, 9-III-2021.
Veronica serpyllifolia L. subsp. serpyllifolia: a l’obaga de la serra de Baier, DG5924, 450 m, vorada de pista forestal, 31-XII-2019; sobre can plana, DG5921, 325 m, vorada humida de pista forestal, 21-VI-2020; sobre el sot de Rocablanca, DG5922, 405 m, vorada forestal, 21-VI-2020.
Vicia peregrina L.: vora sant Martí de Riells, DG5925, 485 m, herbassar, 26-V-2012; a Fogueres de Montsoriu, DG6025, 630 m, vora de camí, 26-III-2021; a can Badia, DG6026, 675 m, vorada, 26-III-2021.
*Wisteria sinensis (Sims) Sweet: a can Tries, DG6021, 245 m, marge de prat, 2-IV-2021.
*Yucca aloifolia L.: urbanització can Plana, DG6219, 145 m, matollar, 26-II-2020; a la urbanització can Salvà, DG6319, 115 m, talús entre el carrer i el torrent, 10-VII-2021.
*Ziziphus jujuba Mill.: c. cal Ferreric, DG6222, 170 m, bardissa, 12-IX-2020.
207 Butlletí ICHN 85 (4), 2021
AL CONEIXEMENT DE LA FLORA VASCULAR DEL SECTOR ORIENTAL DEL MASSÍS DEL MONTSENY
Gales i organismes cecidògens de Ponent de la coŀlecció Antoni Ribes
Aitor Martínez-Romero*, Antoni Ribes Escolà (†), Berta Caballero-López** & Juli Pujade-Villar*
* Universitat de Barcelona. Facultat de Biologia. Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals (BEECA). Avda. Diagonal, 643. 08028 Barcelona. Catalunya.
** Museu de Ciències Naturals de Barcelona. Laboratori de Natura. Departament d’artròpodes. Passeig Picasso s/n. 08003 Barcelona. Catalunya.
Autor per a la correspondència: Aitor Martínez-Romero. A/e: amartiro70@alumnes.ub.edu
Rebut: 19.11.2021; Acceptat: 09.12.2021; Publicat: 30.12.2021
Resum
La coŀlecció de gales d’Antoni Ribes i Escolà s’ha revisat, catalogat i fotografiat. La coŀlecció conté representants de 134 espècies, que pertanyen als següents ordres: Diptera, Hemiptera, Hymenoptera, Lepidoptera i Acariformes. Els organismes cecidògens han produït les gales sobre 56 espècies d’hostatgers vegetals de 23 famílies diferents, sent les fagàcies, les asteràcies i les salicàcies les que tenen una major representació entre les mostres. S’han identificat 10 espècies no citades anteriorment per a la península Ibèrica: 4 dípters (Asphondylia baudysi Vimmer, 1937, Lasioptera ephedricola Cockerell, 1902, Rabdophaga degeerii (Bremi, 1847) i Stefaniella cecconii (Kieffer, 1909)), 2 himenòpters (Pontania kriechbaumeri (Konow 1901) i Euura viminalis (Linnaeus 1758)) i 4 àcars (Acalitus brevitarsus (Fockeu, 1890), Aceria cephalonea (Nalepa, 1922), A. myriadeum (Murray, 1877) i A. salviae (Nalepa, 1891)). Es citen per primer cop a Catalunya 16 espècies, 13 Diptera (Asphondylia conglomerata Stefani, 1900; Lipara lucens Meigen, 1830; Oedaspis fissa Löw, 1862; Probruggmanniella phillyreae (Tavares, 1907); Psectrosema tamaricis (Stefani, 1902); Rabdophaga clavifex (Kieffer, 1891); R. rosaria (Loew, 1850); Rhopalomyia ambrosinae Gagne, 2004; R. producticeps Kieffer, 1912; R. tubifex (Bouché, 1847); Stefaniella brevipalpis Kieffer, 1898; Stefaniola bilobata (Kieffer, 1913) i Xerephedromyia ustjurtensis Fedotova, 1992), 2 Hemiptera (Hayhurstia atriplicis (Linnaeus, 1761) i Pemphigus immunis Buckton, 1896) i un Hymenoptera (Diastrophus mayri Reinhard, 1876).
Paraules clau: Antoni Ribes, Coŀlecció, gales, zoocecidis, Ponent, Lleida, Catalunya.
Abstract
Galls and cecidogenic organisms of Ponent from the Antoni Ribes collection
The gall collection of Antoni Ribes-Escolà has been reviewed, documented and photographed. It contains 134 different gall-producing species belonging to Diptera, Hemiptera, Hymenoptera, Lepidoptera and Acariformes orders. The studied collection represents 56 plant species grouped into 23 different families with Fagaceae, Asteraceae and Salicaceae being the most well represented families. It has been identified 10 species not previously reported from the Iberian Peninsula: 4 Diptera (Asphondylia baudysi Vimmer, 1937, Lasioptera ephedricola Cockerell, 1902, Rabdophaga degeerii (Bremi, 1847) and Stefaniella cecconii (Kieffer, 1909)), 2 Hymenoptera (Pontania kriechbaumeri (Konow 1901) and Euura viminalis (Linnaeus 1758)) and 4 Acari (Acalitus brevitarsus (Fockeu, 1890), Aceria cephalonea (Nalepa, 1922), A. myriadeum (Murray, 1877) and A. salviae (Nalepa, 1891)). We have also found 16 species that are first records for Catalonia: 13 Diptera (Asphondylia conglomerata Stefani, 1900; Lipara lucens Meigen, 1830; Oedaspis fissa Löw, 1862; Probruggmanniella phillyreae (Tavares, 1907); Psectrosema tamaricis (Stefani, 1902); Rabdophaga clavifex (Kieffer, 1891); R. rosaria (Loew, 1850); Rhopalomyia ambrosinae Gagne, 2004; R. producticeps Kieffer, 1912; R. tubifex (Bouché, 1847); Stefaniella brevipalpis Kieffer, 1898; Stefaniola bilobata (Kieffer, 1913) and Xerephedromyia ustjurtensis Fedotova, 1992), 2 Hemiptera (Hayhurstia atriplicis (Linnaeus, 1761) and Pemphigus immunis Buckton, 1896) and a Hymenoptera (Diastrophus mayri Reinhard, 1876).
Key words: Antoni Ribes, Collection, galls, zoocecidia, Ponent, Lleida, Catalonia.
Introducció
Els organismes cecidògens són aquells que tenen la capacitat de produir malformacions sobre òrgans i teixits vegetals mitjançant la inducció de substàncies alienes a la planta o altres accions especifiques, el que permet l’aprofitament d’aquesta estructura anormal en el seu cicle vital. Aquests organismes són, en la seva majoria, artròpodes (principal-
DOI: 10.2436/20.1502.01.94
ment himenòpters, dípters, hemípters, lepidòpters, coleòpters i àcars), tot i que també son capaços de fer gales organismes bacterians, fúngics, vírics o nematodes (Nieves-Aldrey, 1998).
Les gales presenten moltes morfologies diferents, el que comporta modificacions estructurals, tant externes com internes (llenyosa, carnosa, coloració, pilositat, nombre variable de cambres larvals,...), podent arribar en alguns casos a ser
209 Butlletí ICHN 85 (4), 2021
GEA, FLORA ET FAUNA GEA, FLORA ET FAUNA
Butlletí de la Institució Catalana d’Història Natural, 85 (4): 209-223. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition)
confoses amb fruits (Pujade-Villar, 2013). Hi ha una especificació molt gran respecte al gènere o espècie de la planta hostatgera, de manera que un mateix organisme produeix la gala en una espècie en concret o un grup d’espècies molt properes filogenèticament i sovint en el mateix òrgan (Pujade-Villar, 2013).
Les gales sorgeixen com a resultat d’un procés coevolutiu entre la planta i l’organisme inductor, per la qual cosa tenen un valor adaptatiu, tant per a l’hostatger, com pel fitòfag hoste (Price et al., 1986, 1987). Per a la planta hostatgera pot existir un doble benefici; primerament, la formació de la gala aïlla a l’inductor en l’espai i en el temps podent neutralitzar els efectes tòxics del causant de la gala sobre la planta; en segon lloc, la presència de tanins de moltes gales, afavoreix que les plantes que en posseeixen moltes, no siguin atacades pels grans herbívors (Margalef, 1974). Per a l’inductor, la gala augmenta la disponibilitat d’aliment i proporciona protecció davant del medi abiòtic i dels enemics naturals (Nieves-Aldrey, 1998).
Els processos mecànics i químics implicats en al formació de gales són encara força desconeguts (Folliot, 1977; PujadeVillar,1986; Nieves-Aldrey, 1998). Tot i així, Hearn et al. (2019) s’esclareix que entre els mecanismes presents en la formació de gales d’artròpodes es descarta la implicació de virus o microorganismes portats pels insectes, i també que s’activen alguns gens responsables de la modificació dels teixits vegetals al voltant de l’ou o la larva inductora, que posteriorment estimulen el desenvolupament de les noves estructures a través de la manipulació de les hormones de la planta o els enzims que modulen el metabolisme de l’hoste.
Pel que fa al coneixement de les gales a la península Ibèrica, a finals del segle passat i durant el primer terç de l’actual s. XXI, han estat publicats diversos estudis monogràfics, com són les gales de la família Cynipidae (Hymenoptera) de Nieves-Aldrey (2001), les gales produïdes pels Cecidomyiidae (Diptera) de Skuhravá et al. (1996, 2006) o les gales dels pugons (Hemiptera) de Nieto-Nafría et al. (2003). També s’han fet estudis generals que citen gales de diferents grups d’una àrea determinada com és el cas de Cogolludo (1921) per a España, de Salas-Remón et al. (2015) per a Murcia, Bellido et al. (2003) i Blanes-Dalmau et al. (2017) per a Catalunya, entre d’altres.
L’estudi taxonòmic de les gales ibèriques, es resumeix en la figura del portuguès Joaquín da Silva Tavares† (Cardigos, 1866 - Paris, 1931), que va elevar el coneixement, principalment dels cinípids, situant-lo al mateix nivell del altres països d’Europa (Luisier, 1932). Amb la mort de Tavares, es va iniciar un llarg període de quasi 50 anys en el que no es va publicar gairebé res sobre cecidis ibèrics, amb l’excepció dels dos treballs de l’Antoni Vilarrúbia (1936, 1956). A partir dels anys 80 els estudis de Nieves-Aldrey i del darrer autor han contribuït enormement en el coneixement d’aquesta fauna en aquest territori, principalment pel que fa a la família Cynipidae. La manca d’estudis generalistes de gales en el nostre àmbit geogràfic es veu reflectit en la deficient representació d’espècimens en les coŀleccions entomològiques ibèriques, amb les úniques excepcions del Museo de Ciencias Naturales de Madrid i el Museu de Ciències Naturals de Barcelona.
En els Països Catalans, encara que Alessandro Trotter ja havia citat algunes espècies cecidògenes, van ser els naturalistes Domènec Ventalló, Ignasi de Sagarra, Ascensi Codina, Lluís Vilarrúbia i, en un lloc destacat l’Antoni Vilarrúbia, els qui es van fer càrrec d’aquest camp que es troba a cavall entre els artròpodes i les plantes vasculars (Pujade-Villar, 2011). Dins d’aquest àmbit, Antoni Ribes i Escolà va contribuir enormement a ampliar els coneixements dels himenòpters a Catalunya.
Antoni Ribes i Escolà† (Lleida, 1968 - Torres de Segre, 2014) va ser un gran naturalista que representava una de les promeses més importants en el camp entomològic actual. Encara que no tenia estudis universitaris es va fer un investigador de renom. La seva especialitat eren els himenòpters de la superfamília Chalcidoidea, concretament la família Eulophidae, essent l’únic expert a Catalunya i Espanya on ja, en el seu segon treball (Ribes, 2011a), esmenta que existeixen 902 espècies d’aquesta superfamília a la província de Lleida citant per primera vegada 88 espècies i 15 gèneres nous per a Espanya. Coŀlaborava amb molts coŀlegues de professió resolent dubtes taxonòmics. El més reconegut és el Dr. Dick Askew, amb qui compartia una estreta relació professional; Askew és considerat el millor especialista d’eulòfids a Europa i tothom considerava que l’Antoni en seria el seu successor algun dia. Antoni Ribes i Escolà va morir prematurament a l’edat de 46 anys deixant com a llegat 13 articles publicats entre el 2009 – 2015 i el projecte en web «Microhimenòpters de Ponent»: «http://ponent.atspace.org/fauna/ins/index.htm», entre d’altres (Pujade-Villar, 2014, 2015). Part de la seva coŀlecció, els calcidoideus, han estat cedits, seguint el desig de l’Antoni, al British Museum (https://www.nhm.ac.uk/our-science/collections/entomology-collections/hymenoptera-collections. html) però les gales, rastres vegetals produïts per artròpodes i els adults dels cinipoideus, van ser legats al darrer autor, també per desig de l’Antoni.
L’objectiu d’aquest estudi és divulgar la gran feina feta per l’Antoni Ribes pel que fa a les gales i restes entomològics dels vegetals i contribuir d’aquesta forma al millor coneixement de la diversitat entomològica de Lleida.
Material i mètodes
La coŀlecció d’estructures vegetals de l’Antoni Ribes consta de 20 capses de mida mitjana (23 cm × 14 cm × 4 cm) i 8 capses de mida petita (13,5 cm × 9 cm × 2 cm) que contenen mostres recollides quasi en la seva totalitat de la regió de Ponent (veure més endavant). Aquestes mostres són principalment gales, encara que també hi ha minadors, còccids, pseudogales, pupes i plantes atacades per fitòfags degut a l’ampli espectre d’estudis en el que l’Antoni Ribes estava implicat. En total s’han revisat 880 mostres de diferents espècies vegetals i prèviament dessecades per garantir-ne la seva preservació.
Cada mostra comptava, en principi, amb una etiqueta original adjunta de l’Antoni on es referenciaven les dades següents: lloc de recoŀlecció, data de coŀlecta, codi, espècie de la planta hostatgera i la seva identificació. Quan alguna
210 Butlletí ICHN 85 (4), 2021
FLORA ET FAUNA A. MARTÍNEZ-ROMERO ET AL.
GEA,
d’aquestes dades mancava, les dades restants s’obtenien o bé consultant la base de dades informatitzada de la coŀlecció (parcialment deteriorada) o bé mitjançant la relació amb altres bosses de la capsa o bé cercant informació en línia a «Microhimenòpters de Ponent».
Tot aquest material s’ha documentat i informatitzat, de manera que cada mostra conté les dades de recoŀlecció originals i les dades taxonòmiques revisades tan de l’organisme cecidògen com de la planta hostatgera. El catàleg es pot consultar a traves de l’enllaç: «https://doi.org/10.15468/dl.n7yfq5».
Pel que fa a les localitats de les mostres, algunes d’elles corresponen a diverses localitats catalanes fora de terres lleidatanes (Albarca: 1, Prades: 2, Ripoll: 3, Sant Joan de les Abadesses: 4 i Sant Jaume de Frontanyà: 5) o d’Aragó (Candasnos: 6, Casp: 7, Penyalba: 8 i Pina de Ebro: 9), però la majoria corresponen a localitats de la província de Lleida (Abella de la Conca: 10, Bausen: 11, Barruera: 12, Bossòst: 13, Cellers: 14, Mont-rebei: 15, Pont de Suert: 16, Senet de Barravés: 17, Sort: 18, Tremp: 19 i Vielha: 20) predominant les pertanyents a la Regió de Ponent (Àger: 21, Aitona: 22, Albatàrrec: 23, Alcanó: 24, Alcoletge: 25, Alfés: 26, Algerri: 27, Aspa: 28, Bellaguarda: 29, Castelldans: 30, Castelló de Farfanya: 31, Ciutadilla: 32, Els Torms: 33, Fraga (AragóBaix Cinca): 34, Fulleda: 35, Gerb: 36, Granadella: 37, Ivars: 38, Juncosa: 39, La Granja d’Escarp: 40, Les Avellanes: 41, Llardecans: 42, Lleida: 43, Menàrguens: 44, Montoliu: 45, Omells de na Gaia: 46, Os de Balaguer: 47, Pobla de Cérvoles: 48, Serós: 49, Sant Llorenç de Montgai: 50, Sudanell: 51, Sunyer: 52, Tàrrega: 53, Torrebesses: 54, Torregrossa: 55, Torres de Segre: 56 i Sarroca de Lleida: 57).
Donat que l’interès principal residia a les gales, qualsevol artròpode trobat que acompanyés a la mostra es guardava en un tub que posteriorment s’adjuntaria a la mostra analitzada quedant registrat aquest adult. Els apèndixs i altres parts trobades també han estat indicats en el catàleg. En els casos que l’organisme trobat no fos el cecidògen (aranyes, parasitoids, etc.) aquesta informació es reflectia en les observacions de la base de dades. En aquest apartat s’indicaven els casos en els que la mostra no era una gala (minador, còccid, mostra per estudis de fitòfags,...) i les espècies vegetals que feien la funció d’hostes. Cada mostra identificada era fotografiada per tenir una referencia gràfica i utilitzar-les com a suport visual en el reconeixement del contingut de la coŀlecció, havent fotografiat 134 espècies de gales, 8 espècies de minadors i 5 espècies de còccids.
Per a la determinació de les mostres estudiades i per conèixer la distribució de les espècies identificades hem consultat les obres de: Vilarrúbia (1936), Dauphin et al. (1993), Nieves-Aldrey (1998, 2001), Nieto-Nafría et al. (2002), Carles-Tolrá (coord.) et al. (2002), Skuhravá et al. (1993, 1996, 2006), Medianero et al. (2007), Ribes-Escolà et al. (2009, 2011a,b), Salas-Remón et al. (2015), Cobo et al. (2016), Blanes-Dalmau et al. (2017), Fusu et al. (2017); i les webs següents: Ribes-Escolà (2012), Jong et al. (2014: Fauna Europaea), Ellis (2018: Plant Parasites of Europe), Noyes (2019: Universal Chalcidoidea Database), Anonimus (2019: Biodiversidad Virtual), Anònim (2020a: Global Biodiversity
Information Facility) i Anònim (2020b: Banc de dades de Biodiversitat de Catalunya).
Una part representativa de totes les mostres identificades està dipositada al Museu de Ciències Naturals de Barcelona, en forma de coŀlecció de referència i una altre coŀlecció de duplicats serà en breu dipositada al CRBA (Universitat de Barcelona). La resta del material està custodiat pel darrer autor.
Resultats
Han estat comptabilitzades un total de 21.375 gales corresponents a 134 espècies de les quals 120 han pogut ser identificades (Taula 1). Entre tot el material vegetal identificat, a més de les gales, també es va trobar un gran nombre d’altres formacions com minadors de fulles, còccids, barrinadors, pseudogales, pupes, postes, nius i material complementari d’estudis relacionats amb les interaccions planta-artròpode per part de l’Antoni.
En la majoria de mostres on es van trobar organismes adults, aquests no estaven relacionats directament amb la gala, sinó constituint el que es denomina com a fauna secundaria (paràsits o inquilins de les gales).
Totes les gales identificades són zoocecidis, produïts per espècies de la classe Arachnida (SubCl. Acari) i al Subfilum Hexapoda (Classe Insecta). Els grups més abundants (Fig. 1a), han estat els dípters (49 espècies) i els himenòpters (43), el que representa el 77 % de les espècies de gales estudiades. En me-
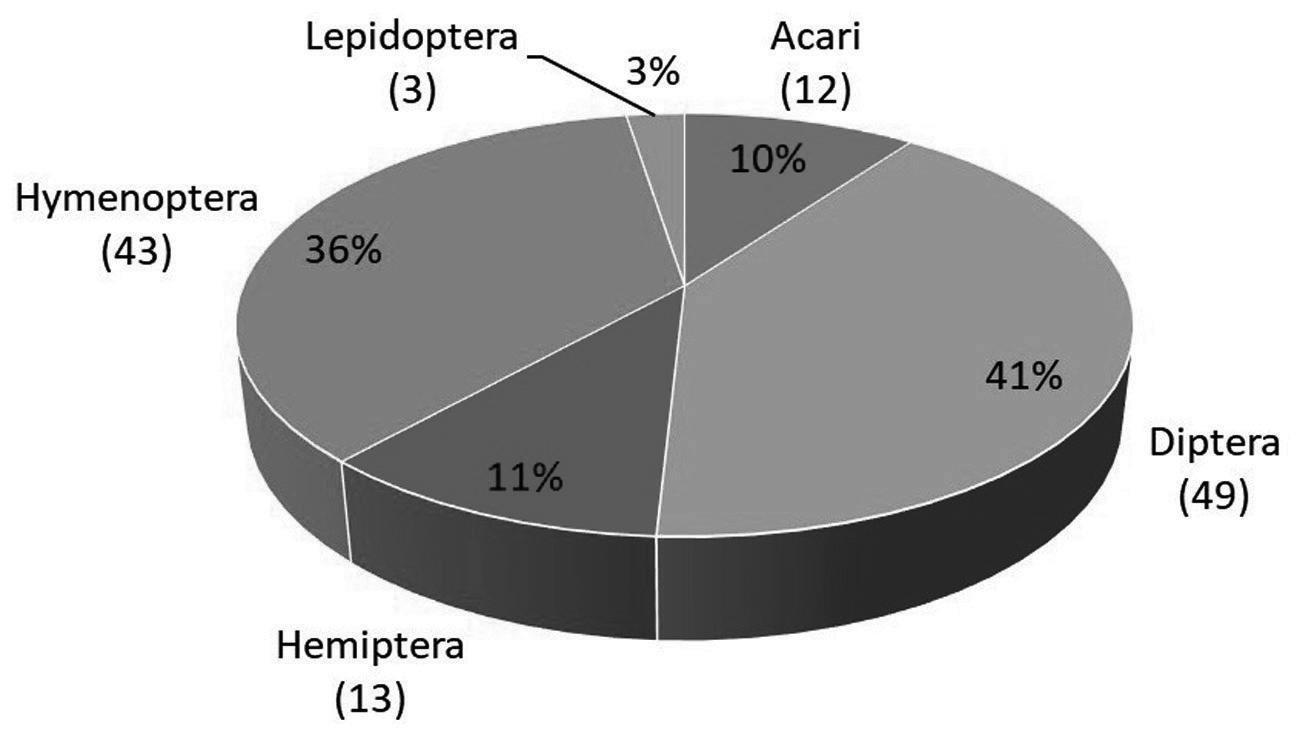
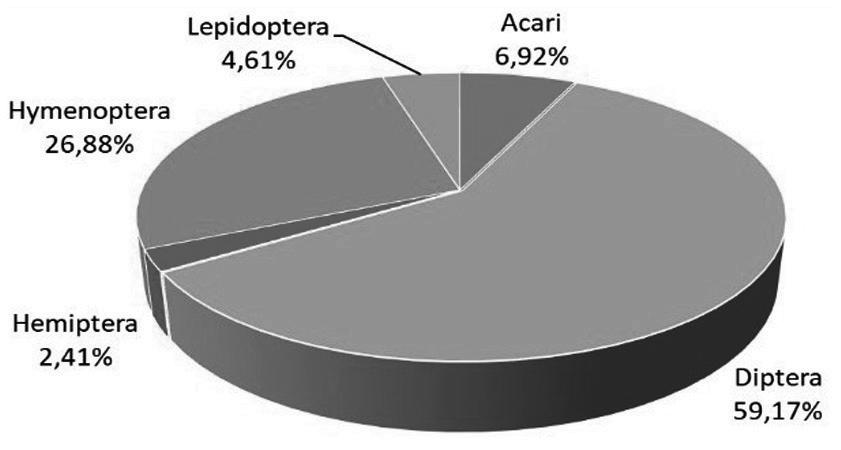
Figura 1. Dades generals de la coŀlecció Antoni Ribes: a) Percentatge d’espècies cecidògenes de cada ordre indicant el nombre d’espècies determinades; b) percentatge del nombre de gales totals estudiades per a cada ordre.
211 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA
I ORGANISMES CECIDÒGENS DE PONENT DE LA COĿLECCIÓ ANTONI RIBES
GALES
b a
GEA, FLORA ET FAUNA
Taula 1. Espècies determinades de la coŀlecció Antoni Ribes, veure el codis de les localitats a l’apartat «Material i mètodes». * primeres cites a la península Ibèrica., ** primeres cites a Catalunya, *** mostres de localitzacions només presents en el CRBA.
Macrodiplosis pustularis (Bremi, 1847)
Mikiola fagi (Hartig, 1839)
Monarthropalpus flavus (Schrank, 1776)
Probruggmanniella phillyreae (Tavares, 1907) **
Psectrosema tamaricis (Stefani, 1902) **
Rabdophaga clavifex (Kieffer, 1891) **
Rabdophaga degeerii (Bremi, 1847) *
Rabdophaga rosaria (Loew, 1850) ** 15, 17, 20
Rabdophaga salicis (Schrank, 1803) 17
Rabdophaga terminalis (Löw, 1850)
Rhopalomyia ambrosinae Gagné, 2004 ** 22, 26, 34, 40, 45, 49, 52, 56, 57
Rhopalomyia navasi Tavares, 1904 22, 34, 40, 49, 52, 56
Rhopalomyia producticeps Kieffer, 1912 ** 49, 56
Rhopalomyia santolinae Tavares, 1902 19, 22, 27***, 39, 49, 50
Rhopalomyia tubifex (Bouché, 1847) ** 26, 40, 34, 45, 49, 56, 57
Stefaniella brevipalpis Kieffer, 1898 **
6, 22, 49, 56
Stefaniella cecconii Kieffer, 1909 * 56
Stefaniola bilobata (Kieffer, 1913) ** 22, 34, 45, 49, 56
Stefaniola salsolae (Tavares, 1904)
9, 22, 27, 34, 40, 43, 45, 49, 52, 55, 56, 57
Xerephedromyia ustjurtensis Fedotova, 1992 ** 22, 55, 57
Chloropidae Lipara lucens Meigen, 1830 **
6, 14, 56
Tephritidae Dithryca guttulosa (Loew, 1869) 55
Myopites inulaedysentericae Blot, 1827 56
212 Butlletí ICHN 85 (4), 2021
Classe Subclasse Ordre Família Espècie Localitats Arachnida Acari Trombidiformes Eriophydae Acalitus brevitarsus (Fockeu, 1890) * 22 Aceria caulobia (Nalepa, 1900) 56 Aceria cephalonea (Nalepa, 1922) * 15 Aceria chondrillae (Canestrini, 1890) 15 Aceria macrochela (Nalepa, 1891) 13 Aceria macrorhyncha (Nalepa, 1899) 5 Aceria myriadeum (Murray, 1877) * 16 Aceria salviae (Nalepa, 1891) * 15 Aculus tetanothrix (Nalepa, 1889) 15, 20, 50, 56 Eriophyes laevis (Nalepa, 1889) 20 Eriophyes tiliae (Pagenstecher, 1857) 15 Stenacis triradiata (Nalepa, 1892) 25 Insecta Pterygota Diptera Cecidomyiidae Arceuthomyia valerii (Tavares, 1904) 29, 39 Arnoldiola quercus (Binnie, 1877) 15 Asphondylia baudysi Vimmer, 1937 * 15 Asphondylia conglomerata Stefani, 1900 ** 49 Asphondylia genistae (Loew, 1890) 45, 57 Asphondylia verbasci (Vallot, 1827) 22, 50 Bayeriola thymicola (Kieffer, 1888) 22, 26, 45 Braueriella phillyreae Löw, 1877 15 Contarinia ilicis Kieffer, 1898 39, 44, 47 Craneiobia corni (Giraud, 1863) 15, 16, 43, 50 Dasineura daphnes (Kieffer, 1901) 50 Dasineura rosae (Bremi, 1847) 15, 22, 56 Dasineura scorpii (Kieffer, 1909) 15, 19, 21, 27, 47 Dictyomyia navasina Tavares, 1919 19, 22, 23, 26, 45, 49, 50, 56 Dryomyia cocciferae (Marchal, 1897) 24, 35 Dryomyia lichtensteini (F. Löw, 1878) 1, 27, 36, 37, 39, 44***, 50 Etsuhoa sabinae (Kieffer, 1898) 34, 50
annulipes (Hartig, 1839) 17, 20
ephedricola Cokerell, 1902 * 22, 55, 57
eryngii (Vallot, 1829) 15, 21, 22, 26, 46, 49
22
Hartigiola
Lasioptera
Lasioptera
Lasioptera rubi (Schrank, 1803)
15, 41
20,
17
15
35
31, 35,
22, 27,
Oligotrophus juniperinus (Linnaeus, 1758)
Oligotrophus panteli Kieffer, 1898
39 Phyllodiplosis cocciferae (Tavares, 1902)
29, 42, 46, 48, 50, 56, 57
39
56
17
15
15
A. MARTÍNEZ-ROMERO ET AL.
1964) 43, 45, 46, 51, 56
Biorhiza pallida (Olivier, 1791)
Callirhytis spec. Foster, 1869
Cynips disticha Hartig, 1840 12, 15, 41, 44
Cynips quercus (Fourcroy, 1785)
Cynips quercusfolii Linnaeus, 1758
Diastrophus mayri Reinhard, 1876 **
15, 33, 41, 44, 46, 47
Diastrophus rubi (Bouché, 1834) 15, 43
Diplolepis eglanteriae (Hartig, 1840) 18, 21, 22, 56
Diplolepis mayri (Von Schlechtendal, 1877)
Diplolepis rosae (Linnaeus, 1758)
15, 22, 26, 33, 41, 44, 48, 50, 56,
4, 10, 14, 15, 19, 21, 22, 26, 44, 46, 48, 56
Neuroterus albipes (Schenck, 1863) 15, 21, 44
Neuroterus anthracinus (Curtis, 1838) 12, 13, 15, 41, 44, 47
Neuroterus numismalis (Fourcroy, 1785) 12
Neuroterus politus Hartig, 1840 15, 19, 33, 44
Neuroterus quercusbaccarum (Linnaeus, 1758) 15, 21, 29, 33, 37, 39, 44, 47
Neuroterus tricolor (Hartig, 1841) 15
Pediaspis aceris (Gmelin, 1790) 15, 46, 48
Phanacis hypochoeridis (Kieffer, 1887) 29, 33, 43***
Plagiotrochus australis (Mayr, 1882) 15, 21, 24, 27, 29, 31, 37, 39, 48, 50
Plagiotrochus burnayi Kieffer, 1901 22
Plagiotrochus coriaceus (Mayr, 1882) 57
Plagiotrochus gallaeramulorum (Fonscolombe, 1832) 22, 24***, 30, 56, 57
Plagiotrochus panteli Pujade-Villar, 1985 44, 46, 57
Plagiotrochus quercusilicis (Fabricius, 1798) 15, 21, 22, 27, 28, 32, 33, 37, 46, 47, 50, 54, 57
Timaspis cichorii (Kieffer, 1909) 56
Timaspis phoenixopodos Mayr, 1882
Trigonaspis synaspis (Hartig, 1841)
Xestophanes potentillae (De Geer, 1773) 43, 56
Eurytomidae Eurytoma gallephedrae Askew, 1998 7, 8, 34
Tenthredinidae Euura viminalis (Linnaeus, 1758) *
Pontania kriechbaumeri (Konow, 1901) * 12, 15
Pontania proxima (Serville, 1823) 12, 17, 22, 50, 56
Lepidoptera Gelechiidae Parapodia sinaica (Frauenfeld, 1859) 34, 40, 43, 56
Tortricidae Cochylimorpha peucedana (Ragonot, 1889) 19, 22, 23, 26
Cochylimorpha santolinana (Staudinger, 1871)
213 Butlletí ICHN 85 (4), 2021
Oedaspis fissa Löw, 1862 ** 23, 26, 40, 45, 56, 57 Ptiloedaspis tavaresiana Bezzi, 1920 22, 27, 34, 40, 45, 49, 52, 56, 57 Hemiptera Aphididae Aploneura lentisci (Passerini, 1856) 22, 33, 37, 50 Baizongia pistaciae (Linnaeus, 1767) 15, 29, 33, 39 Eriosoma lanuginosum (Hartig, 1839) 25 Forda formicaria Von Heyden, 1837 15, 33, 41 Geoica utricularia (Passerini, 1856) 15, 33, 50 Hayhurstia atriplicis (Linnaeus, 1761) ** 56 Pemphigus bursarius (Linnaeus, 1758) 56 Pemphigus immunis Buckton, 1896 ** 15 Pemphigus spyrothecae Passerini, 1860 56 Pemphigus vesicarius Passerini, 1862 15 Tetraneura ulmi (Linnaeus, 1758) 25 Thecabius affinis (Kaltenbach, 1843) 56 Psyllidae Psyllopsis fraxini (Linnaeus, 1758) 15 Hymenoptera Cynipidae Andricus coriarius (Hartig, 1843) 19, 41, 44 Andricus curvator Hartig, 1840 15, 32, 46 Andricus foecundatrix (Hartig, 1840) 11, 15, 19 Andricus hispanicus (Hartig, 1856) 1, 2, 3, 15, 19, 21, 32, 33, 37, 39, 41, 44, 46, 47, 48 Andricus pseudoinflator Tavares, 1901 15, 44 Andricus quercusradicis (Fabricius, 1798) 21, 46 Andricus quercusramuli (Linnaeus, 1761) 15, 37, 44, 46 Andricus quercustozae (Bosc, 1792) 15 Andricus solitarius (Fonscolombe, 1832) 15, 33***, 41, 44 Aylax minor Hartig, 1840 15, 21, 23, 25, 35, 38, 52, 53, 56, 57 Barbotinia oraniensis (Barbotin
GEA, FLORA ET FAUNA
1, 11,
13, 15, 32, 33, 39, 44, 48
44
11,
15
43
1,
1,
2
44
15
22
GALES I ORGANISMES CECIDÒGENS DE PONENT DE LA COĿLECCIÓ ANTONI RIBES
GEA,

nor nombre es troben els hemípters (13 espècies) i els àcars (12), essent molt més escasses les gales de lepidòpters (3).
Avaluant cada ordre per la seva capacitat per produir gales sobre els diferents òrgans vegetals, els dípters representen més de la meitat de les gales coŀlectades amb un 59,17 % (Fig. 1b).
La proporció entre espècies determinades d’àcars i hemípters és semblant (Fig 1a), però al comparar la proporció d’abundància de gales (Fig1b) d’ambdós grups es fa evident que el nombre de gales produïdes pels àcars (6,92 %) és més del doble de les produïdes per hemípters (2,41 %). Les ga-
les de lepidòpters estan representades per un 3 % i 4,61 % de la coŀlecció, tan s’hi ens mirem la riquesa d’espècies com l’abundància de mostres.
D’àcars s’han identificat 12 espècies pertanyents a 5 gèneres de la família Eriophyidae (Taula 1). La morfologia predominant de les gales dels àcars són pústules (Fig. 2a) i protuberàncies que es troben la superfície de les fulles. Les gales que es localitzen a tiges, flors i borrons, tenen l’aparença de deformacions coraŀloides (Fig. 2b) o piloses. Un total de quatre espècies no estaven citades fins ara a la península Ibèrica (Fig. 2c-f): Acalitus brevitarsus (Fockeu, 1890), Aceria cep-
214 Butlletí ICHN 85 (4), 2021
FLORA ET FAUNA
MARTÍNEZ-ROMERO ET AL. e f b d c a
Figura 2. Gales produïdes per eriòfids (Acariformes): a) Aceria macrorhyncha sobre Acer opalus, b) Aceria. chondrillae sobre Chondrilla juncea, c) Acalitus brevitarsus sore Alnus sp., d) Aceria cephalonea sobre Acer monspessulanum, e) Aceria myriadeum sobre Acer campestre, f) Aceria salviae sobre Salvia pratensis
A.
GALES I ORGANISMES CECIDÒGENS DE PONENT DE LA COĿLECCIÓ ANTONI RIBES

halonea (Nalepa, 1922), A. myriadeum (Murray, 1877) i A. salviae (Nalepa, 1891).
Pel que fa als dípters han estat identificades 49 espècies pertanyents a 30 gèneres de les famílies Tephritidae (4 espècies) Chloropidae (1) i Cecidomyiidae (44). Les formes i característiques de les gales de dípters són molt variades (Fig. 3a, b), trobant tant inflaments de tiges com de fruits, invaginacions com evaginacions foliars, gales unicamerals com multicamerals, erupcions pubescents sobtades, etc. La majoria de les espècies eren conegudes a la península Ibèrica, però quatre d’elles representen primeres cites de cecidòmids: Asphondylia baudysi Vimmer, 1937 (Fig. 3c), Lasioptera ep-
215 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA
Figura 3. Gales produïdes per cecidòmids (Diptera): a) Craneiobia corni sobre Cornus sanguinea, b) Bayeriola thymicola sobre Thymus sp., c) Asphondylia baudysi sobre Coronilla minima, d) Lasioptera ephedricola sobre Ephedra sp , e) Rabdophaga degeerii sobre Salix purpurea, f) Stefaniella cecconii sobre Atriplex patula, g) Asphondylia conglomerata sobre Atriplex halimus, h) Probruggmanniella phillyreae sobre Phyllyrea sp.
e b g d a h f c
GEA, FLORA ET FAUNA

216
ICHN 85 (4),
Butlletí
2021
A. MARTÍNEZ-ROMERO ET AL. i h g f e d c b a
Figura 4. Gales produïdes per cecidòmids (Diptera): a) Psectrosema tamaricis sobre Tamarix sp., b) Rabdophaga rosaria sobre Salix sp., c) Rhopalomyia ambrosinae sobre Artemisia herba-alba, d) Rhopalomyia producticeps sobre Artemisia herba-alba, e) Rhopalomyia tubifex sobre Artemisia campestris, f) Rabdophaga clavifex sobre Salix caprea, g) Stefaniella brevipalpis sobre Atriplex halimus, h) Stefaniola bilobata sobre Salsola vermiculata, i) Xerephedromyia ustjurtensis sobre Ephedra distachya
GALES I ORGANISMES CECIDÒGENS DE PONENT DE LA COĿLECCIÓ ANTONI RIBES


217 Butlletí ICHN 85 (4), 2021
GEA, FLORA ET FAUNA
Figura 6. Gales produïdes per àfids (Hemiptera): a) Pemphigus spyrothecae sobre Populus nigra, b) Forda formicaria sobre Pistacia terebinthus, c) Hayhurstia atriplicis sobre Atriplex halimus, d) Pemphigus immunis sobre Populus nigra
Figura 5. Gales produïdes per cloròpids i tefrítids (Diptera): a) Lipara lucens sobre Phragmites australis, b) Oedaspis fissa sobre Artemisia herba-alba.
b c a b a a d


218 Butlletí ICHN 85 (4), 2021
GEA, FLORA ET FAUNA
Figura 7. Gales produïdes per Hymenoptera: a) Eurytoma gallephedrae sobre Ephedra nebrodensis, b) Cynips quercusfolii (agam) sobre Quercus sp., c) Pontania kriechbaumeri sobre Salix eleagnos, d) Euura viminalis sobre Salix purpurea, e) Diastrophus mayri sobre Potentilla sp.
A. MARTÍNEZ-ROMERO ET AL. d c b a e
Figura 8. Gala produïda per Lepidoptera: Parapodia sinaica sobre Tamarix sp.
GALES I ORGANISMES CECIDÒGENS DE PONENT DE LA COĿLECCIÓ ANTONI RIBES
GEA, FLORA ET FAUNA
Taula 2. Espècies cecidògenes de la coŀlecció Antoni Ribes i família vegetal atacada, indicant l’òrgan i l’hostatger on es localitza la gala.
Família vegetal
Amaranthaceae
Anacardiaceae
Espècie hoste cecidògena
Aceria caulobia
Asphondylia conglomerata
Hayhurstia atriplicis
Stefaniella brevipalpis
Stefaniella cecconii
Stefaniola bilobata
Stefaniola salsolae
Aploneura lentisci
Baizongia pistaciae
Forda formicaria
Geoica utricularia
Apiaceae Lasioptera eryngii
Asteraceae
Betulaceae
Aceria chondrillae
Cochylimorpha peucedana
Cochylimorpha santolinana
Dictyomyia navasina
Dithryca guttulosa
Myopites inulaedysentericae
Oedaspis fissa
Phanacis hypochoeridis
Ptiloedaspis tavaresiana
Rhopalomyia ambrosinae
Rhopalomia navasi
Rhopalomyia producticeps
Rhopalomyia santolinae
Rhopalomyia tubifex
Timaspis cichorii
Timaspis phoenixopodos
Acalitus brevitarsus
Eriophyes laevis
Buxaceae Monarthropalpus flavus
Cornaceae Craneiobia corni
Cupressaceae Arceuthomyia valerii
Etsuhoa sabinae
Oligotrophus juniperinus
Oligotrophus panteli
Ephedraceae
Fabaceae
Eurytoma gallephedrae
Lasioptera ephedricola
Xerephedromyia ustjurtensis
Asphondylia baudysi
Asphondylia genistae
Dasineura scorpii
Fagaceae Andricus coriarius
Andricus curvator
Andricus foecundatrix
Andricus hispanicus
Andricus pseudoinflator
Andricus quercusradicis
Andricus quercusramuli
Andricus quercustozae
Andricus solitarius
Arnoldiola quercus
Biorhiza pallida
Callirhytis spec.
Contarinia ilicis
Cynips disticha
Cynips quercus
Cynips quercusfolii
Dryomyia cocciferae
Dryomyia lichtensteini
Òrgan atacat i espècie hostatgera vegetal
Tija de Suaeda vera Forssk. Ex. J.F.Gmel.
Tija d’Atriplex halimus L.
Fulla d’Atriplex halimus L.
Tija d’Atriplex halimus L.
Tija d’Atriplex patula L.
Borró de Salsola vermiculata L.
Borró de Salsola vermiculata L.
Fulla de Pistacia lentiscus L.
Fulla de Pistacia terebinthus L.
Fulla de Pistacia terebinthus L.
Fulla de Pistacia terebinthus L.
Tija d’Eryngium sp.
Borró de Chondrilla juncea L.
Tija de Santolina chamaecyparissus L.
Tija de Santolina rosmarinifolia L.
Borró de Santolina chamaecyparissus L.
Tija de Santolina rosmarinifolia L.
Flor de Inula sp.
Tija d’Artemisia herba-alba Asso.
Tija de Hypochaeris sp.
Borró d’Artemisia herba-alba Asso.
Borró d’Artemisia herba-alba Asso.
Tija d’Artemisia herba-alba Asso.
Tija d’Artemisia herba-alba Asso.
Tija de Santolina sp.
Tija d’Artemisia campestris L.
Tija de Cichorium intybus L.
Tija de Lactuca sp.
Fulla d’Alnus sp.
Fulla d’Alnus sp.
Fulla de Buxus sp.
Fulla de Cornus sanguinea L.
Borró de Juniperus oxycedrus L.
Borró de Juniperus sabina L.
Borró de Juniperus oxycedrus L.
Borró de Juniperus oxycedrus L.
Tija d’Ephedra nebrodensis Tineo Ex. Guss
Tija d’Ephedra sp.
Tija d’Ephedra distachya L.
Fruit de Coronilla minima L.
Fruit de Genista scorpius (L.) DC.
Borró de Genista scorpius (L.) DC.
Borró de Quercus pubescens Mill.
Fulla de Quercus sp.
Borró de Quercus sp.
Borró de Quercus sp.
Borró de Quercus sp.
Arrels de Quercus sp.
Flor de Quercus sp.
Borró de Quercus sp.
Borró de Quercus sp.
Fulla de Quercus sp.
Borró de Quercus spp.
Tija de Quercus sp.
Fulla de Quercus ilex L.
Fulla de Quercus petraea (Matt.),Liebl.
Fulla de Quercus spp.
Fulla de Quercus sp.
Fulla de Quercus coccifera L.
Fulla de Quercus sp.
219 Butlletí ICHN 85 (4), 2021
GEA, FLORA ET FAUNA
Família vegetal Espècie hoste cecidògena
Hartigiola annulipes
Macrodiplosis pustularis
Mikiola fagi
Neuroterus albipes
Neuroterus anthracinus
Neuroterus numismalis
Neuroterus politus
Neuroterus quercusbaccarum
Neuroterus tricolor
Phyllodiplosis cocciferae
Plagiotrochus australis
Plagiotrochus burnayi
Plagiotrochus coriaceus
Plagiotrochus gallaeramulorum
Plagiotrochus panteli
Plagiotrochus quercusilicis
Trigonaspis synaspis
Lamiaceae Aceria salviae
Bayeriola thymicola
Malvaceae Eriophyes tiliae
Oleaceae Braueriella phillyreae
Probruggmanniella phillyreae
Psyllopsis fraxini
Papaveraceae Aylax minor
Barbotinia oraniensis
Poaceae Lipara lucens
Rosaceae Diastrophus mayri
Diastrophus rubi
Diplolepis eglanteriae
Rosaceae Diplolepis mayri
Diplolepis rosae
Lasioptera rubi
Dasineura rosae
Xestophanes potentillae
Salicaceae
Sapindaceae
Aculus tetanothrix
Euura viminalis
Pemphigus bursarius
Pemphigus immunis
Pemphigus spyrothecae
Pemphigus vesicarius
Pontania kriechbaumeri
Pontania proxima
Rabdophaga clavifex
Rabdophaga degeerii
Rabdophaga rosaria
Rabdophaga salicis
Rabdophaga terminalis
Stenacis triradiata
Thecabius affinis
Aceria cephalonea
Aceria macrochela
Aceria macrorhyncha
Aceria myriadeum
Pediaspis aceris
Scrophulariaceae Asphondylia verbasci
Tamaricaceae Parapodia sinaica
Psectrosema tamaricis
Thymelaeaceae Dasineura daphnes
Ulmaceae Eriosoma lanuginosum
Tetraneura ulmi
Òrgan atacat i espècie hostatgera vegetal
Fulla de Fagus sylvatica L.
Fulla de Quercus faginea Lam.
Fulla de Fagus sylvatica L.
Fulla de Quercus sp.
Fulla de Quercus petraea (Matt.),Liebl.
Fulla de Quercus petraea (Matt.),Liebl.
Borró de Quercus faginea Lam.
Fulla i Flor de Quercus sp.
Fulla de Quercus sp.
Borró de Quercus spp.
Fulla de Quercus spp.
Fruit de Quercus coccifera L.
Fulla de Quercus ilex L.
Tija de Quercus coccifera L.
Borró de Quercus coccifera L.
Fulla de Quercus spp.
Fulla de Quercus sp.
Fulla de Salvia pratensis L.
Borró de Tymus sp.
Fulla de Tilia sp.
Fulla de Phillyrea sp.
Fruit de Phillyrea sp.
Fulla de Fraxinus sp.
Fruit de Papaver sp.
Fruit de Papaver sp.
Borró de Phragmites australis (Cav.) Trin. Ex Steud.
Tija de Potentilla sp.
Tija de Rubus sp.
Fulla de Rosa sp.
Fulla de Rosa sp.
Fulla de Rosa sp.
Fulla de Rubus sp.
Fulla de Rosa sp.
Pecíol de Potentilla sp.
Fulla de Salix spp.
Fulla de Salix purpurea L.
Pecíol de Populus nigra L.
Pecíol de Populus nigra L.
Pecíol de Populus nigra L.
Pecíol de Populus nigra L.
Fulla de Salix eleagnos Scop.
Fulla de Salix spp.
Borró de Salix caprea L.
Tija de Salix purpurea L.
Borró de Salix spp.
Fulla de Salix eleagnos Scop.
Fulla de Salix eleagnos Scop.
Flor de Salix sp.
Fulla de Populus sp.
Fulla d’Acer monspessulanum L.
Fulla d’Acer campestre L.
Fulla d’Acer opalus Mill.
Fulla d’Acer campestre L.
Fulla d’Acer monspessulanum L.
Flor de Verbascum sp.
Tija de Tamarix sp.
Tija de Tamarix sp.
Borró de Daphne sp.
Borró de Ulmus minor Mill.
Fulla de Ulmus minor Mill.
220 Butlletí ICHN 85 (4), 2021
ET AL.
A. MARTÍNEZ-ROMERO
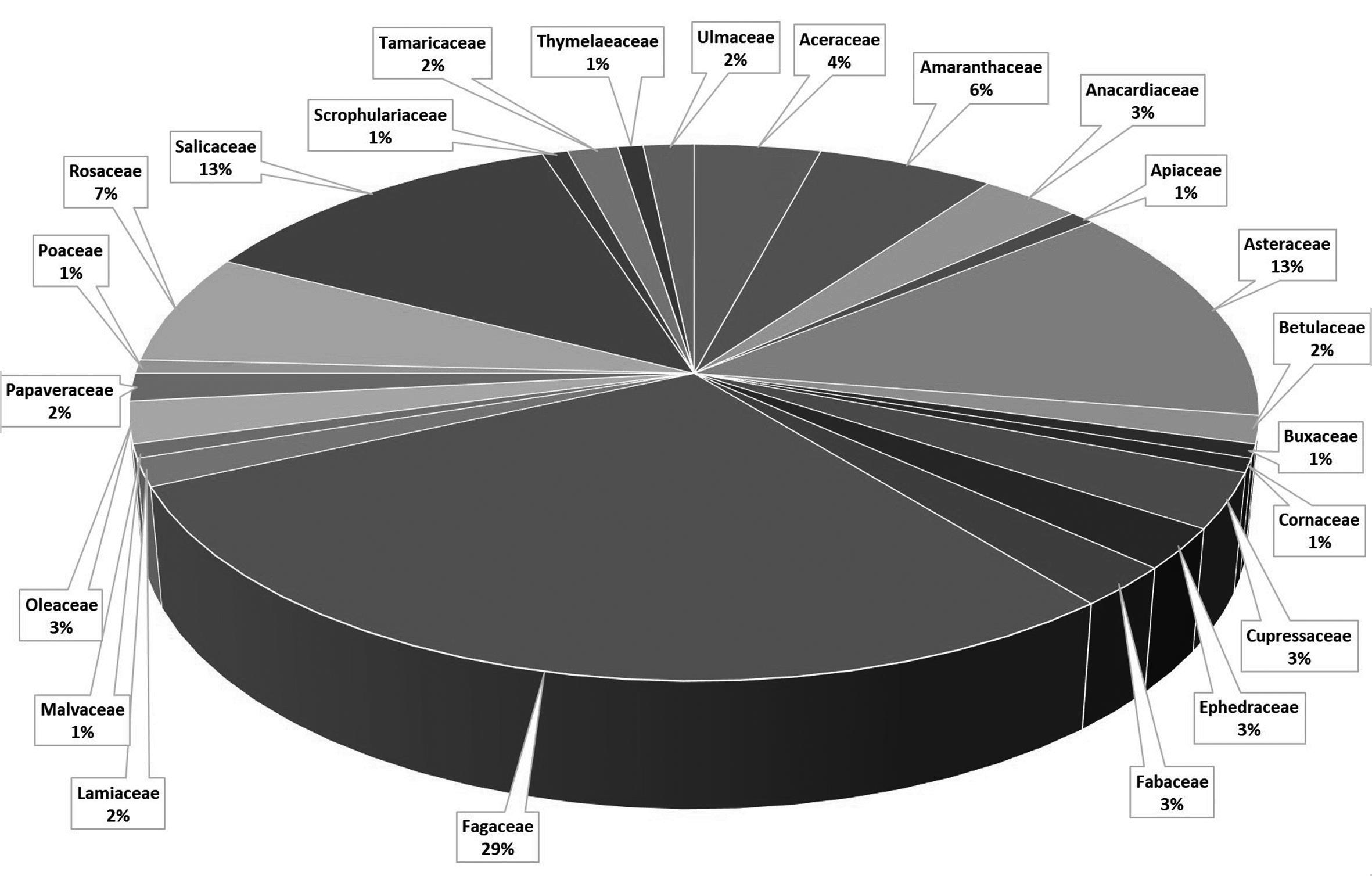
hedricola Cockerell, 1902 (Fig. 3d), Rabdophaga degeerii (Bremi, 1847) (Fig. 3e) i Stefaniella cecconii (Kieffer, 1909) (Fig. 3f). Per a Catalunya, la coŀlecció d’Antoni Ribes aporta les primeres cites de 13 espècies: Asphondylia conglomerata Stefani, 1900 (Fig. 3g), Lipara lucens Meigen, 1830 (Fig. 5a), Oedaspis fissa Löw, 1862 (Fig. 5b) Probruggmanniella phillyreae (Tavares, 1907) (Fig. 3i), Psectrosema tamaricis (Stefani, 1902) (Fig. 4a), Rabdophaga clavifex (Kieffer, 1891) (Fig. 4f), R. rosaria (Loew, 1850) (Fig. 4b), Rhopalomyia ambrosinae Gagne, 2004 (Fig. 4c), R. producticeps Kieffer, 1912 (Fig. 4d), R. tubifex (Bouché, 1847) (Fig. 4e), Stefaniella brevipalpis Kieffer, 1898 (Fig. 4g), Stefaniola bilobata (Kieffer, 1913) (Fig. 4h) i Xerephedromyia ustjurtensis Fedotova, 1992 (Fig. 4i).
D’hemípters s’han identificat 13 espècies pertanyents a 10 gèneres de la família Aphididae i 1 gènere de la família Psyllidae. Hi ha formes molt peculiars entre les gales d’hemípters: enduriment de fruits hipertrofiats, inflament de borrons, malformacions al pecíol o recargolaments i plecs de les fulles (pseudogales). Hi ha combinacions entre recargolaments (Fig. 6a) i enduriments que donen lloc a formes semicirculars (Fig. 6b). Totes les espècies es troben incloses en els registres de fauna peninsular, però a Catalunya s’esmenten per primer cop les espècies Hayhurstia atriplicis (Linnaeus, 1761) (Fig. 6c) i Pemphigus immunis Buckton, 1896 (Fig. 6d).
Pel que fa als himenòpters s’han identificat 43 espècies pertanyents a 18 gèneres de 3 famílies, en la que predominen els Cynipidae sobre els Tenthredinidae i els Eurytomidae.
Les gales induïdes per les espècies d’aquest ordre d’insectes són les més complexes. La major part d’espècies produeixen gales en les fulles, tot i que també es troben sovint en borrons (Fig. 7a) i més rarament a les tiges i els fruits. Són més freqüents les gales globulars, eŀlíptiques i uniloculars en les fulles (Fig. 7b), mentre que a les tiges sovintegen els engruiximents multicamerals (Fig. 7e). Són primeres cites per a la península Ibèrica: Pontania kriechbaumeri (Konow 1901) (Fig. 7c) i Euura viminalis (Linnaeus 1758) (Fig. 7d), i per a Catalunya: Diastrophus mayri Reinhard, 1876 (Fig. 7e).
Finalment els lepidòpters, l’ordre més reduït en la coŀlecció, queden representats amb 3 espècies identificades de 2 gèneres, pertanyents a les famílies Gelechiidae i Tortricidae. La morfologia de les gales és similar; totes elles comparteixen òrgan vegetal (tija) essent la gala un engrossiment fusiforme amb un únic forat d’emergència (Fig. 8). Totes elles es troben ja citades anteriorment a la península Ibèrica.
Pel que fa als hostatgers vegetals, aquests estan representats per 23 famílies (Taula 2), de les que destaquen fagàcies, asteràcies i salicàcies (Fig. 9). Les fagàcies són els hostatgers més diversificats entre les espècies cecidògenes ja que han estat comptabilitzats 35 espècies d’organismes cecidògens. Els òrgans més afectats han estat les fulles i els borrons.
Considerant el nombre de gales total recomptat en la coŀlecció, les famílies vegetals amb més gales induïdes en les mostres estudiades han estat les fagàcies (6.603 gales), seguides per les amarantàcies (3.728), asteràcies (3.119) i papaveràcies (1.943).
221 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA
GALES I ORGANISMES CECIDÒGENS DE PONENT DE LA COĿLECCIÓ ANTONI RIBES
Figura 9. Percentatge d’espècies d’organismes cecidògens per a cada família vegetal.
A. MARTÍNEZ-ROMERO ET AL.
GEA, FLORA ET FAUNA
Discussió
Han restat per identificar 21 mostres corresponents a 14 espècies diferents.
Pel que fa a la totalitat dels diferents ordres analitzats, es pot considerar que l’alta proporció de dípters obtinguda està justificada per la coneguda importància i estudis relacionats a aquest ordre com a principals agents cecidògens. Segons Pujade-Villar (2019), «La família Cecidomyiidae és possiblement la més diversa en nombre d’espècies cecidògenes dins dels insectes», i segons Skuhravá (1996), «Els cecidòmids són una de les famílies de dípters amb més diversitat dins de l’àmbit mundial». Així que, de les 49 espècies de dípters presents en la coŀlecció 44 d’elles (més d’un 89 %) són cecidòmids.
De manera similar, la proporció baixa dels lepidòpters entre les mostres de la coŀlecció, estaria justificada al comparar-la amb l’anterior ordre, tenint en compte que hi ha moltes menys espècies cecidògenes. En la coŀlecció cal destacar l’espècie Parapodia sinaica (Frauenfeld, 1859) de la família Galechiidae, que ha estat citada recentment per primer cop a la regió de Murcia (Salas-Remón et al., 2015) i de la que s’han estudiat 14 mostres amb un total de 976 gales. Això posa en evidència que segurament no és una espècie rara sinó que la seva absència es deguda a la manca de mostreig.
En el cas dels àcars, hem constatat que el nombre espècies no citades a la península Ibèrica és molt superior a les d’altres ordres, a més, aquestes espècies no citades fins ara representen més de la meitat d’espècies d’àcars determinats. Això és degut a que hi ha pocs estudis sobre les espècies d’àcars a la península Ibèrica i menys de les espècies cecidògenes, ja que la seva taxonomia és complexa i pateix modificacions continues. En el panorama actual, es coneix que la gran majoria dels àcars cecidògens pertanyen a la família Eriophyiidae, que formen gales en plantes molt diverses, però principalment de la família de les salicàcies, fagàcies, rosàcies i aceràcies.
El recompte de gales estudiades associades a espècies d’hemípters es baix pel que fa a les mostres i al número de localitats on s’han obtingut aquestes. De igual manera les estructures induïdes per les espècies de l’ordre Hemiptera identificades a la coŀlecció tenen característiques molt marcades en quan a les formes peculiars, colors vius i mida; a més, es produeixen en òrgans molt exposats dels seus hostatgers vegetals per la qual cosa es poden detectar molt fàcilment, el que hauria facilitat la tasca de localització i recoŀlecció. No obstant això, el seu recompte es menor en relació als altres ordres en proporció (Fig. 1a-b). Aquestes variacions en el nombre, poden ser degudes a que la inducció de gales per part d’aquest grup és menys freqüent.
A la web «Microhimenòpters de Ponent» es descriuen les principals famílies d’himenòpters que actuen com a agents cecidògens en el subordre Apocrita. Algunes de les mostres de la coŀlecció havien estat utilitzades per Ribes per completar les descripcions i fotografies en diferents seccions de la família Cynipidae, però hi ha altres seccions que no es troben actualitzades tot i tenir mostres i fotografies que l’autor podia haver utilitzat; la mort sobtada li ho va impedir. És el
cas, per exemple, de Plagiotrochus burnayi Kieffer, 1901 i Diastrophus mayri Reinhard, 1876. A la web (http://ponent. atspace.org/fauna/ins/index.htm) no s’hi troba la superfamília Tenthredinoidea, i no hi es present degut a que és un macrohymenoptera; malgrat això l’Antoni Ribes havia coŀlectat dues espècies del gènere Pontania i una del gènere Euura
En comparació amb l’estudi de la coŀlecció Vilarrúbia dipositada al Museu de Ciències Naturals de Barcelona (Blanes-Dalmau et al., 2017), pel que fa a la proporció de les espècies cecidògenes per a cada ordre i a les famílies d’hostes vegetals presents els resultats són molt similars. Aquesta relativa homogeneïtat, després de la gran quantitat d’esforç de mostreig en ambdues coŀleccions, ha de ser deguda a que és més fàcil coŀlectar espècies que fan gales en determinades plantes que d’altres, d’aquí la coincidència de resultats malgrat els diferents interessos de coŀlecta d’ambdues coŀleccions; l’Antoni Ribes coŀlectà principalment a Ponent estudiant relacions tròfiques entre insectes i plantes, mentre que l’Antoni Vilarrúbia coŀlectà principalment a Tona buscant biodiversitat per enriquir la coŀlecció de gales del Museu de Zoologia de Barcelona.
Agraïments
A la família Ribes, en especial atenció a la Maite Ribes Escolà, per haver cedit la coŀlecció de gales i mostres vegetals al darrer autor de la qual ha sorgit aquest estudi. A Neus Brañas (Myrmex SL, MCNB) per documentar tota la coŀlecció Ribes i a Jordi Agulló i Katia Cezón pel seu suport en la confecció del llistat que permet divulgar les dades al GBIF.
Bibliografia
Anònim. 2017. Ponent (regió) [en línia]. Disponible en: <https:// ca.wikipedia.org/wiki/Ponent_(regió)> [Data de consulta: 16 de Març 2019]
Anònim. 2019. Biodiversidad Virtual. Insectarium Virtual [en línia]. Disponible en: <https://www.biodiversidadvirtual.org/ insectarium/> [Data de consulta: 15 de Juliol 2018]
Anònim. 2020a. GBIF: The Global Biodiversity Information Facility [en línia]. Disponible en: <https://www.gbif.org/es/> [Data de Consulta: 27 de Gener de 2020]
Anònim. 2020b. Banc de dades de Biodiversitat de Catalunya. Generalitat de Catalunya i Universitat de Barcelona [en línia]. Disponible en: <http://biodiver.bio.ub.es/biocat/homepage.htlm> [Data de consulta: 16 de Gener 2020]
Askew, R. R. & Blasco-Zumeta, J. 1998. Insects associated with galls of a new species of Eurytomidae (Hymenoptera: Chalcidoidea) on Ephedra nebrodensis in Spain. Journal of Natural History, 32: 805-821.
Bellido, D., Ros-Farré, P. & Pujade-Villar, J. 2003. Coŀlecció Villarrúbia I: Gaŀles dipositades al Museu de Zoologia de Barcelona. Sessió Conjunta d’Entomologia de la ICHN-SCL, XII:102-138.
Blanes-Dalmau, M., Caballero-López, B., & Pujade-Villar J. 2017. Estudi de les gales de la Coŀlecció Vilarrúbia dipositada al Museu de Ciències Naturals de Barcelona. Butlletí de la Institució catalana d’Història natural, 81: 137-173.
222 Butlletí ICHN 85 (4), 2021
Carles-Tolrá, M., (coord.) et al. 2002. Catálogo de los Díptera de España, Portugal y Andorra (Insecta). En: Monografías S.E.A.
Vol. 8. Sociedad Entomológica Aragonesa. Zaragoza. 323 p.
Cobo, A., Ribes, A. & Pujade-Villar, J. 2016. Parasitoides Calcidoideos (Hymenoptera: Chalcidoidea) asociados a Isocolus melikai Pujade-Villar, 2014 (Hymenoptera: Cynipidae). Boletín de la Sociedad Entomológica Aragonesa, 59: 153–156.
Cogolludo J. 1921. Contribución al conocimiento de las zoocecidias de España. Museo Nacional de Ciencias Naturales (CSIC) Madrid, Serie Botánica, 16: 1-117.
Dauphin, P. & Aniotsbehere, J. C. 1993. Les galles de France. En: Memoires de la S.L.B., tome 2. Societe Linneenne de Bordeaux. Bordeaux. 316 p.
Ellis, W. N. 2018. Plant Parasite of Europe [en línia]. Disponible en: <https://bladmineerders.nl/> [Data de consulta: 20 de Juliol 2018]
Folliot, R. 1977. Les insectes cécidogènes et la cécidogenèse. En: Traité de Zoologie. Grasse, P. P. (Ed.). Volume 8, Fasc. V, B. Masson. Paris, P. 389-429.
Fusu, L. & Ribes, A. 2017. Description of the first Palaearctic species of Tineobius Ashmead, 1896 with DNA data, a checklist of world species, and nomenclatural changes in Eupelmidae (Hymenoptera, Chalcidoidea). European Journal of Taxonomy, 263: 1–19. DOI: 10.5852/ejt.2017.263
Hearn, J., Blaxter, M., Schönrogge, K., Nieves-Aldrey, J. L., PujadeVillar, J., Huguet, E., Crezenm, J. M., Shorthouse, J. D. & Stone, G. N. 2019. Genomic dissection of an extended phenotype: Oak galling by a cynipid gall wasp. PLoS Genet, 15(11): e1008398. DOI: 10.1371/journal.pgen.1008398
Jong, Y. de, Verbeek, M., Michelsen, V., Place Bjørn P. de, Los, W., Steeman, F., Bailly, N., Basire, C. Chylarecki, P., Stloukal, E., Hagedorn, G., Wetzel, F. T., Glöckler, F., Kroupa, A., Korb, G., Hoffmann, A., Häuser, C., Kohlbecker, A., Müller, A., Güntsch, A., Stoev, P. & Penev, L. 2014. Fauna Europaea - all European animal species on the web. Biodiversity Data Journal 2: e4034. DOI: 10.3897/BDJ.2.e4034. Disponible en: <https://fauna-eu. org/> [Data de consulta: 14 de Febrer 2019]
Luisier, A. 1932. In memoriam le R. P. J. da Silva Tavares, S. J. Brotéria, 1932: 1-26.
Margalef, R. 1974. Ecología. Omega. Barcelona. 951 p (2ª edición).
Medianero, E., Parra, L. A., Sánchez, I. & Nieves-Aldrey, J. L. 2007. Agallas inducidas por insectos en especies de Artemisia (Asteraceae) en España, con especial referencia a la comunidad de Madrid. Boletín Sociedad Entomológica Aragonesa, 41: 291302.
Nieto-Nafría, J. M., Mier-Durante, M. P., Binazzi, A., & Pérez Hidalgo, N. 2002. Hemíptera, Aphididae II. En: Fauna Ibérica, vol. 19. Ramos, M.A. et al (Eds.). Museo Nacional de Ciencias Naturales. CSIC. Madrid. 336 p.
Nieves-Aldrey, J. L. 1998. Insectos que inducen la formación de agallas en las plantas: una fascinante interacción ecológica y evolutiva. Boletín Sociedad Entomológica Aragonesa, 23: 3-12.
Nieves-Aldrey, J. L. 2001. Hymenoptera, Cynipidae. En: Fauna Ibérica, vol. 16. Museo Nacional de Ciencias Naturales. CSIC. Madrid. 640 p.
Noyes, J. S. 2019. Universal Chalicoidea Database. World Wide Web electronic publication [en línia]. Disponible en: <http://www.nhm. ac.uk/chalcidoids> [Data de consulta: 18 de Juliol 2018]
Price, P. W. Fernandes, G. W. & Waring, G. L., 1987. Adaptive nature of insect gall. Environmental Entomology, 16: 15-24.
Price, P. W., Waring, G. L. & Fernandes, G. W. 1986. Hypotheses on the adaptive nature of galls. Proceedings of the Entomological Society of Washington, 88: 361-363.
Pujade-Villar, J. 1986. Estudi preliminar: els cecidis i la cecidología. Ed. Eumo. Coŀlecció l’entorn, 9: 27-39.
Pujade-Villar, J. 2013. Las agallas de los encinos: un ecosistema en miniatura que hace posible estudios multidisciplinares. Entomología mexicana, 12: 1-20.
Pujade-Villar, J. 2014. IN MEMORIAM, Antoni Ribes Escolà (Lleida, 1968-2014). Butlletí de la Institució Catalana d’Història Natural, 78: 143-145.
Pujade-Villar, J. 2015. Nota necrològica: Antoni Ribes Escolà (Lleida, 1968 – Torres de Segre, 2014). Boletín Asociación española de Entomología, 39 (1-2): 191-196.
Ribes, A. 2011a. Some Chalcidoidea (Hymenoptera) from Lleida new to the Spanish fauna. Boletín de la Sociedad Entomológica Aragonesa, 48: 337-343.
Ribes, A. 2011b. A new species of Hyssopus Girault from Spain and France (Hymenoptera: Chalcidoidea: Eulophidae). Boletín de la Sociedad Entomológica Aragonesa, 48: 237-241.
Ribes, A. 2012. Microhimenòpters de ponent [en línia]. Disponible en: <http://ponent.atspace.org/fauna/ins/index.htm> [Data de consulta: 6 de Maig 2018]
Ribes Escolà, A. & Askew, R. R. 2009. Chalcidoidea (Hymenoptera) reared from fruits of Juniperus phoenicea, with descriptions of three new species. Boletín Sociedad Entomológica Aragonesa, 45: 109-121.
Salas-Remón, P., Llimona, X., Lozano, T., Alcaraz, F. & PujadeVillar, J. 2015. Aportación al conocimiento de los cecidios encontrados en la vegetación de Murcia y localidades próximas del SE de España (Arthropoda: Insecta y Acari; Bacterii). Anales de Biología, 37: 43-68.
Skuhravá, M., Skuhravý, V. & Blasco-Zumeta, J. 1993. Gall midges (Diptera: Cecidomyiidae) of Aragón (Spain). A review of species found in the period 1890-1990 with new records for the Monegros region. Revista aragonesa de Entomología, Zapateri, 3.
Skuhravá, M., Skuhravý, V., Blasco-Zumeta, J. & Pujade- Villar, J. 2006. Gall midges (Diptera: Cecidomyiidae) of the Iberian Peninsula 2. Zoogeographical analysis of the gall midge fauna. Boletín de la Asociación Española de Entomología, 30 (1-2): 93-159.
Skuhravá, M., Skuhravý, V., Blasco-Zumeta, J. & Pujade-Villar, J. 1996. Gall midges (Diptera: Cecidomyiidae) of the Iberian Peninsula. Boletín de la Asociación española de Entomología, 20 (1-2): 41-61.
Vilarrúbia, A., 1936. Les zoocecidies de les plantes de Catalunya. Treballs del Museu de Ciències Naturals de Barcelona, Sèrie Entomològica IX, 10: 1-106.
Xue, X. F., Wang, Z., Song, Z. W. & Hong, X. Y. 2009. Eriophyoid mites on Fagaceae with descriptions of seven new Genera and eleven new species (Acari: Eriophyoidea). Zootaxa, 2253: 1-95.
223 Butlletí ICHN 85 (4), 2021 GEA, FLORA ET FAUNA
GALES I ORGANISMES CECIDÒGENS DE PONENT DE LA COĿLECCIÓ ANTONI RIBES
NOTA BREU
Sobre la distribución de especies de la familia Ciidae Leach, 1819 (Coleoptera) en la Península Ibérica no contempladas en el nuevo Catálogo Paleártico
On the distribution of species of the family Ciidae Leach, 1819 (Coleoptera) in the Iberian Peninsula not included in the new Catalogue Palaearctic
Amador Viñolas* & Josep Muñoz-Batet*
* Museu de Ciències Naturals de Barcelona. Laboratori de Natura. Coŀlecció d’artròpodes. Passeig Picasso, s/n. 08003 Barcelona. A/e: av.rodama@gmail.com
Rebut: 26.11.2021. Acceptat: 10.12.2021. Publicat: 30.12.2021
Viñolas & Muñoz-Batet (2015), junto con la descripción de Cis onyosi Viñolas & Muñoz-Batet, 2015 indicaron en una tabla las especies del género Cis Latreille, 1796 citadas por diferentes autores del área peninsular con posterioridad al Catálogo Paleártico de Jelínek (2008) en lo que se refiere al apartado correspondiente a la familia Ciidae Leach, 1819 del volumen 5 dicho catálogo.
La sorpresa, al revisar el nuevo Catálogo Paleártico (Tenebrionoidea. Revised and Updated Second Edition. Volumen 5. (Królik, 2020)), ha sido no ver en él reflejadas todas las especies citadas como nuevas, por diferentes autores, para la Península y Francia durante estos doce años, ni tan solo las citadas en el trabajo anteriormente mencionado de Viñolas & Muñoz-Batet (2015) con excepción de la nueva especie Cis onyosi. Solamente indica como nueva para España a Sulcacis bidentulus (Rosnhauer, 1847) con especímenes recolectados por K. Rudzisńki en Torrente de Cinca (Huesca) y depositados en su colección.
Es incongruente la mala información sobre la distribución de las especies que da el nuevo Catálogo, ya que, como se puede comprobar, no se ha realizado una búsqueda bibliográfica exhaustiva, y la información que aporta es falsa, pudiendo crear confusión y errores a los autores que lo consulten. Era de esperar que en un volumen de esta magnitud y, además, de tan elevado importe, la información que aportara fuera la correcta y actualizada.
A continuación se detalla la información sobre las especies citadas bibliográficamente del área peninsular de la familia Ciidae y que no están reflejadas en el nuevo Catálogo Paleártico (Królik, 2020).
Género Cis Latreille, 1796
Cis festivus Panzer, 1793
Pérez Moreno & Moreno Grijalba (2009) citan la especie del Parque Natural Sierra de Cebollera (La Rioja) y Diéguez Fernández (2012) indica su presencia en el Montseny (Barcelona).
Cis fissicollis Mellié, 1848
Diéguez Fernéndez (2012, 2013) cita la especie de Vidrà (Girona) y de los Montes de Vitoria (Araba).
Cis glabratus Mellié, 1848
Diéguez Fernández (2014b) cita la especie del Parc Natural de la Serra de Collserola (Barcelona), con dos ejemplares obtenidos por emergencia de un Fomitopsidaceae, Fomitopsis pinicola (Sw.) P. Karst sobre un Pinus L. muerto.
Cis laminatus Mellié, 1848
Viñolas et al. (2013) citan, por primera vez, la especie de la Península Ibérica, con una numerosa serie de ejemplares recolectados con trampas de interceptación de vuelo en Vimbodí i Poblet (Tarragona). Viñolas (2020) la cita por segunda vez del área peninsular con ejemplares recolectados mediante trampas de interceptación de vuelo en Blanes (Girona).
Cis onyosi Viñolas & Muñoz-Batet, 2015
Especie descrita de Sant Bartomeu del Grau (Barcelona) y citada posteriormente por Viñolas (2020) de Mura (Barcelona) y Blanes (Girona) con especímenes recolectados mediante trampas de interceptación de vuelo. Rose (2017) lo cita por primera vez de Francia, con ejemplares recolectados en Sanginet (Landes) y Cestas (Gironde), mediante trampas de interceptación de vuelo y por emergencia de un Fomitopsidaceae, Fomitopsis pinicola (Sw.) P. Karst.
Cis pygmaeus pygmaeus (Marsham, 1802)
Pérez Moreno & Moreno Grijalba (2009) citan la especie del Parque Natural Sierra de Cebollera (La Rioja), Pérez Moreno (2010) también indica su presencia en Almarza de Cameros y Nieva de Cameros (La Rioja) con la captura de dos ejemplares recolectados mediante trampa de tubo y Diéguez Fernández (2012) lo cita de Banyoles (Girona).
225 Butlletí ICHN 85 (4), 2021
NOTA BREU
edition)
Butlletí de la Institució Catalana d’Història Natural, 85 (4): 225-226. 2021 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print
10.2436/20.1502.01.95
DOI:
Cis rugulosus Mellié 1848
Pérez Moreno & Moreno Grijalba (2009) indican su presencia en el Parque Natural Sierra de Cebollera (La Rioja), Diéguez Fernández (2012, 2014a) lo cita de Molins de Rey y Montseny (Barcelona), Llívia (Girona), Siresa (Huesca), Montagut (Lleida), Samos (Lugo), Cantalojas (Guadalajara) y Valls (Tarragona). Vorst (2013) indica su presencia, con una serie de ejemplares recolectados en un viejo hongo sin determinar, en Enirio-Aralarko (Gipuzkoa).
Género Hadraule C.G. Thomson, 1859
Hadraule elongatula (Gyllenhal, 1827)
Viñolas et al. (2014) citan, por primera vez la especie de la Península Ibérica, a partir de un ejemplar recolectado mediante trampa de luz UV en Tabernas (Almería).
Género Rhopalontus Mellié, 1847
Rhopalontus baudueri Abeille de Perrin, 1874
Viñolas et al. (2015) lo citan, por primera vez de la Península Ibérica, con un ejemplar recolectado por emergencia de un Polyporaceae del género Fomes (Fr.) Fr. procedente de Masquefa (Barcelona). Muñoz-Batet et al. (2021) con un ejemplar recolectado mediante polytrap indican su presencia en la Jonquera (Girona).
Bibliografía
Diéguez Fernández, J. M. 2012. Aportaciones al conocimiento de la corología ibérica de algunas especies de coleópteros saproxílicos micetófagos (Coleoptera: Ciidae, Mycetophagidae, Leiodidae). Heteropterus Revista de Entomología, 12 (1): 65-77.
Diéguez Fernández, J. M. 2013. Coleoptera. Familia Ciidae. Algunas citas de cíidos de España. Arquivos Entomolóxicos, 8: 103-106.
Diéguez Fernández, J. M. 2014a. Registros interesantes de coleópteros para España (Insecta: Coleoptera). 3ª nota. Arquivos Entomolóxicos, 10: 119-124.
Diéguez Fernández, J. M. 2014b. Catálogo de los Coleoptera de la Sierra de Collserola (Barcelona, NE de España): primeros resul-
tados. Arquivos Entomolóxicos, 10: 235-264.
Jelínek, J. 2008. Ciidae, P. 55-62. In: Löbl, I. & Smetana, A. (ed.): Catalogue of Palaeartic Coleoptera. Vol. 5. Apollo Books. Stenstrup. 670 p.
Królik, R. 2020. Family Ciidae. P. 33-43. In: Iwan, D. & Löbl, I. (Eds.). Tenebrionoidea. Revised and Updated Second Edition. Volumen 5. Brill. Leiden. 945 p.
Muñoz-Batet, J., Soler, J. & Viñolas, A. 2021. Osmoderma eremita (Scopoli, 1763) al Paratge Natural d’Interès Nacional de l’Albera (Coleoptera: Scarabaeidae: Cetoniinae) amb notes sobre altres coleòpters saproxílics interessants. Butlletí de la Institució Catalana d’Història Natural, 85 (3): 87-90.
Pérez Moreno, I. 2010. Nuevas aportaciones al conocimiento de la fauna de coleópteros saproxílicos (Coleoptera) del Sistema Ibérico Septentrional, I: Robledales del valle medio del Iregua (Sierra de Cameros, La Rioja, España). Boletín de la Sociedad Entomológica Aragonesa, 46 : 321-334.
Pérez Moreno, I. & Moreno Grijalba, F. 2009. Los coleópteros saproxílicos del Parque Natural Sierra de Cebollera (La Rioja). Ciencias de la Tierra, 28. Instituto de Estudios Riojanos. Logroño. 180 p.
Rose, O. 2017. Cis onyosi Viñolas & Muñoz-Batet, espèce nouvelle pour la faune de France (Coleoptera Ciidae). L’Entomologiste, 73 (2): 83-84.
Viñolas, A. 2020. Nuevas aportaciones al conocimiento de la fauna coleopterológica de Cataluña, Península Ibérica. Nota 9a. (Coleoptera). Arquivos Entomolóxicos, 22: 113-129.
Viñolas, A. & Muñoz-Batet, J. 2015. Una nueva especie del género Cis Latreille, 1796 de la comarca de Osona, Cataluña, España (Coleoptera: Tenebrionoidea: Ciidae). Arquivos Entomolóxicos, 13 (2014): 123-130.
Viñolas, A., Muñoz-Batet, J. & Soler, J. 2016. Noves o interessants localitzacions d’espècies de coleòpters per a la península Ibèrica i illes Canàries (Coleoptera). Butlletí de la Institució Catalana d’Història Natural, 80: 101-112.
Viñolas, A., Muñoz, J., Mencuccini, M. & Benvenuti, F. 2013. Nuevos datos sobre Rushia parreyssi (Mulsant, 1856), Melandrydae Leach, 1815 y otros coleópteros interesantes de la sierra de Prades, Tarragona (Coleoptera). Orsis, 27: 29-51.
Viñolas, A., Muñoz Batet, J., Bentanachs, J. & Abós, L. 2014. Nuevos registros de coleópteros interesantes de Cataluña y Almería (Península Ibérica) (Coleoptera). Arquivos Entomolóxicos, 10: 25-38.
Vorst, O. 2013. On some Gipuzkoan Coleoptera, including several species new to the Iberian Peninsula. Heteropterus, Revista de Entomología, 13 (2): 147-173.
226 Butlletí ICHN 85 (4), 2021
A. VIÑOLAS & J. MUÑOZ-BATET
NOTA BREU
Butlletí de la Institució Catalana d’Història Natural, 85 (4): 227-230. 2021
NOTA BREU
ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition)
L’home part de la biosfera: 50 anys d’un article seminal de Ramón Margalef
Man as part of the Biosphere: 50 years of an oustanding paper from Ramon Margalef
Narcís Prat Fornells*
* Universitat de Barcelona. Facultat de Biologia. Grup de recerca FEHM (Freshwater Ecology, Hydrology and Management). Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals. Avda. Diagonal, 643. 08028 Barcelona. A/e: nprat@ub.edu
Rebut: 03.11.2021. Acceptat: 10.12.2021. Publicat: 30.12.2021
Ramón Margalef (1919-2004) fa mig segle va publicar un estudi en el que proposava avaluar l’impacte de la humanitat sobre el planeta amb dos paràmetres, la taxa de creixement demogràfic (r) i la taxa de creixement en l’ús de l’energia (f). Els dos creixements eren exponencials i Margalef trobava diferències entre els països desenvolupats (r = 0,015; f = 0,039) i poc desenvolupats (r = 0,035; f = 0,015). Aquest resultat el va utilitzar moltes vegades al llarg dels seus escrits. Lluny de quedar-se en l’anàlisi de les dades, Margalef feia profundes reflexions de les conseqüències d’aquest creixement (dit demofòric). Hem fet un petit càlcul de com ambdós paràmetres han evolucionat al llarg del període 2010-2016 i veiem com en aquests moments es poden donar quatres situacions (r positiva/negativa; f positiva/negativa) que són característiques de diferents països. La mitjana mundial pel període analitzat fou de r=0,011 i f= -0,009. Les reflexions de Margalef són de total actualitat, profundes i crítiques i mostren la gran personalitat de l’ecòleg de la biosfera.
Margalef, el creixement demofòric i les seves conseqüències
Ara fa 50 anys el Professor Ramon Margalef publicava als Treballs de la Societat Catalana de Biologia una reflexió sobre els efectes de l’home sobre la biosfera (Margalef, 1971) amb el títol: «L’home part de la biosfera i objecte d’estudi de l’Ecologia». Deia: «La població humana creix i consumeix energia i tota mena de recursos a velocitat accelerada. La conseqüència és que la Terra se’ns va quedant petita». Margalef feia tot un seguit de reflexions que han resultat gairebé profètiques (tot i que ell no creia en les profecies) com «...l’explotació de la natura porta un problema.... que té a veure amb la desigualtat entre els homes i les dones» o una altra: «Quan augmenta el flux d’energia..... augmenta també la seva variància. Encara que pugui disminuir la possibilitat de morir de fam, es fa més palès el control d’unes nacions per les altres, o d’uns grups humans per uns altres». I es preguntava: «Pot renunciar l’home a fer ús d’aquesta capacitat de control?».
En el seu article Margalef feia servir dos paràmetres per caracteritzar l’impacte de l’home en la biosfera: la demografia i el consum energètic. Més gent vol dir més impacte sobre la naturalesa. L’impacte de l’energia usada per l’home el va descriure més endavant Margalef de forma molt sintètica
en dos capítols de llibre (Margalef, 1976, 1989). En aquest treballs, Margalef mostrava com el desenvolupament de les societats (des dels caçadors recoŀlectors fins a les societats tecnològiques) depenia del seu consum d’energia externa (o exosomàtica). Si el dividia per l’energia endosomàtica, li donava una ràtio individual que anava de 2 pels països més pobres a més de 100 pels Estats Units (EEUU).
Margalef, en el seu treball de 1971, definia dos coeficients, r = taxa instantània d’augment net de la població (N) i f = taxa instantània en l’ús individual de l’energia (E), i l’acceleració de l’acció de l’home a la biosfera com la suma d’ambdós (r+f). Els dos coeficients feien créixer exponencialment la població o l’energia, (Margalef, 1971: figura 2). Els països en procés de desenvolupament presentaven uns valors r = 0,035 i f =0,015, mentre que en els països desenvolupats els valors d’aquests coeficients eren a l’inrevés. La suma final era la mateixa (0,05). Conclusió, ambdós grups de països tenen impactes similars. Margalef feia un símil amb un cotxe que premia l’accelerador a fons. Quan algú deia que es veia una paret al fons de la carretera, els dos conductors responien que encara era molt lluny i que ja hi hauria temps de parar. S’ignorava l’impacte ambiental o l’esgotament dels recursos per mantenir un creixement econòmic sostingut al llarg del temps i mantenir els creixements exponencials (també dels diners, o sia increment del PIB mundial).
Margalef va seguir explicant aquesta idea en diferents treballs posteriors, per exemple en el seu llibre Our Biosphere, (1997): «...els consums d’energia i altres recursos no tenen límits. Segueixen l’efecte de Mateu o la llei de Pareto, i es troben en l’origen de les preocupants diferències entre el que, de manera eufemística, anomenem països desenvolupats i els països en desenvolupament. Amb més tristesa que orgull, reprodueixo a continuació una petita taula (Taula 1) del meu llibre d’Ecologia (1974), encara vàlida i eloqüent per si sola».
Cinquanta anys desprès, els efectes del model de desenvolupament econòmic dels humans, s’estan fent notar. Estem a l’Antropocé (Crutzen, 2000). L’informe de l’IPCC (2021) és molt clar i és difícil avui negar que l’home no sigui el causant de l’increment dels gasos d’efecte hivernacle. Un dels canvis que Margalef assenyalava és la transformació de la majoria de paisatges de la Terra, que han canviat d’una matriu natural
227 Butlletí ICHN 85 (4), 2021
NOTA BREU
DOI: 10.2436/20.1502.01.96
Taula 1. Valors de r i f calculats per Margalef el 1971 i reproduïts en el llibre «Ecologia» (1974) i a «Our Biosphere» (1997).
amb algunes taques de la petjada de l’home (pobles i petites carreteres) a un sistema de ciutats relligades entre elles per grans autopistes i que encerclen les poques taques verdes que queden entre elles Margalef (1985, 2003). És el que ell anomenava «canvis en la topologia del Paisatge». Tello (2013) va mostrar que els paisatges agroforestals en mosaic tenien una major biodiversitat i eren més productius que els monocultius extensius, tal com deia Margalef.
Al cap de 50 anys d’aquell treball ens podem preguntar si aquella idea de Margalef encara té validesa. A la taula 2 mostrem els valors calculats de (r) i de (f) per a una sèrie de països (que comprenen el 80 % de la població mundial) i el període 2010-2016, amb la finalitat de buscar algun patró similar al que deia Margalef. Les dades procedeixen de diferents fonts estadístiques públiques (ONU, BP...). Ens podríem preguntar ara si usar només l’energia i la demografia era suficient per sintetitzar tots els canvis ambientals. Margalef el 1971 advertia: «...repeteixo, aquesta predicció no es pot estendre a més d’una o dues desenes d’anys».
El creixement demofòric als inicis del segle XXI
El creixement demogràfic de la humanitat ha continuat essent exponencial al llarg dels darrers 100 anys. En l’article de Margalef, la població de la terra era la meitat de l’actual, i, per tant, és de suposar que els valors de (r) han seguit augmentant. A la taula 2 podem veure que la mitjana mundial de la taxa instantània de creixement (r) pel període 2010-2016 era de 0,011, la taxa instantània de creixement en energia (f) era negativa (f=-0,002) i el valor de r+f resultava de 0,0091. Valors que són inferiors als de Margaglef a la Taula 1. O sigui que el planeta hauria baixat la seva acceleració però encara no estaria frenant (r+f mitjà mundial és positiu). Recordem que aquest fou un període de crisi econòmica, pel que la (f) mundial en conjunt és negativa (Taula 2). Segons l’ONU (2021), el creixement poblacional aviat serà negatiu i, amb les tendències actuals, el 2100 s’assolirà el màxim de població (11000 milions), l’evolució dels valors de (f) estan per veure.
Per veure com variaven amb un cert detall els valors de r i f pel període 2010-2015, hem seleccionat 19 països, que inclouen els més poblats i una representació de tots els continents (Taula 2). Els hem agrupat segons quatre situacions possibles: 1) Decreixement de r i f; 2) Decreixement de r i augment de f; 3) Creixement de r i disminució de f; 4) Increment de f i r a la vegada.
En primer lloc tenim països que decreixen tant en població com en consum d’energia primària (Rússia i Japó). La suma de r+f en els dos casos és negativa. Mentre que en el cas de
Taula 2. Valors de diferents paràmetres demogràfics, energètics i econòmics de diferents països del món (valors entre 2010 i 2016). S’han seleccionat els països amb més població ja que són els principals emissors de carboni a l’atmosfera (TnCO2/cap) en global. Elaboració pròpia a partir de dades estadístiques de fonts diverses. Estan ordenats en funció dels valors positius o negatius de: r = taxa instantània d’augment de la població. f= taxa instantània d’augment del consum d’energia. En primer lloc països amb r i f negativa, en segon lloc països amb r negativa i f positiva, en tercer lloc països amb r positiva i f negativa i finalment en països amb r i f positives. Endo/Exo = Relació entre energia endosomàtica i exosomàtica. Ha/ cap = Petjada Ecològica. A part dels 19 països també s’indica la mitjana mundial a la darrera fila que inclou tots els països del món.
228 Butlletí ICHN 85 (4), 2021 NOTA BREU N. PRAT FORNELLS
r f r+f Països desenvolupats 0,015 0,039 0,054 Països en desenvolupament 0,035 0,015 0,050
Fills/pers r f r+f Exo/Endo TnCO2/cap Ha/cap PIB/càpita Rússia EU 1,66 -0,0005 -0,0040 -0,0045 48,74 19,55 3,70 24451 Japó Àsia 1,40 -0,0022 -0,0177 -0,0199 37,29 11,73 4,90 40743 Alemanya EU 1,39 -0,0006 0,0005 -0,0001 41,67 11,80 1,60 48014 Regne Unit EU 1,70 0,0053 -0,0246 -0,0193 31,10 9,09 5,30 41756 Sudàfrica Àfrica 2,40 0,0083 -0,0228 -0,0145 23,52 4,58 2,10 13996 Espanya EU 1,32 0,0001 -0,0120 -0,0119 30,57 7,56 1,20 34906 Mèxic NA 2,29 0,0124 -0,0236 -0,0112 14,89 5,09 3,40 16988 Egipte Àfrica 3,38 0,0196 -0,0227 -0,0031 9,49 3,10 1,10 10193 USA NA 1,89 0,0073 -0,0099 -0,0026 73,53 19,43 9,40 56116 Etiòpia Àfrica 4,20 0,0288 -0,0156 0,0132 3,29 1,81 0,80 1630 Nigèria Àfrica 5,40 0,0263 -0,0010 0,0253 1,32 1,65 1,44 6004 Brasil SA 1,82 0,0080 0,0055 0,0135 14,53 14,15 2,40 15391 Pakistan Àsia 3,72 0,0203 0,0053 0,0256 4,17 1,88 0,80 5010 Colòmbia SA 1,93 0,0085 0,0258 0,0343 9,08 3,53 1,90 13829 Xina Àsia 1,55 0,0043 0,0336 0,0379 22,75 9,04 2,10 14450 Índia Àsia 2,48 0,0118 0,0293 0,0411 5,47 2,24 0,90 6100 Indonesia Oceania 2,50 0,0112 0,0350 0,0462 7,78 2,96 1,10 11057 Filipines Oceania 3,04 0,0151 0,0367 0,0518 3,80 1,61 0,90 7387 Bangladesh Àsia 2,23 0,0118 0,0674 0,0792 1,95 1,11 0,60 3397 Món 4,50 0,0111 -0,0020 0,0091 18,33 7,12 2,70 15672
Japó els dos descensos semblen estructurals i que continuaran en el temps, en el cas de Rússia el descens en el consum d’energia podria ser un fet puntual derivat de la crisi econòmica. Per bé que les taxes individuals disminueixin, els valors totals d’emissions de CO2 o la petjada ecològica dels dos països eren molt alts, el que indica que l’impacte ambiental actual dels dos països és rellevant. Rússia té un valor de Tn CO2/càpita molt elevat, el que indica la poca eficiència del seu sistema de generació d’energia.
En segon lloc podem veure com Alemanya és el cas d’un país on hi havia un petit decreixement demogràfic i un petit creixement del consum d’energia. El valor final de r+f és negatiu (-0,0595) com passa a molts països desenvolupats. Alemanya ha començat a frenar el creixement demofòric si ens atenem a les polítiques que està emprenent destinades a disminuir les emissions de CO2. El seu impacte global és, però, tan important com el de Japó, i tant el rati Exo/Endo com les Tn de CO2/càpita són molt elevades (Taula 2).
Tot seguit tenim països que tenen creixement demogràfic positiu i un decreixement energètic. Alguns tenen una r+f final petita (per exemple Espanya o Gran Bretanya. Interessant és el cas dels Estats Units i Nigèria, dos països on en el període estudiat, hi havia un creixement demogràfic (molt més elevat a Nigèria) amb un decreixement de l’ús de l’energia. Mentre Nigèria te un valor positiu de r+f (0,0261), els EEUU el tenen negatiu (-0,0026) (Taula 2). Observi´s però que l’impacte era molt diferent, tant si ho valorem en termes d’emissions de CO2 com en termes de petjada ecològica (Taula 2). A molts països llatinoamericans hi ha una frenada de r i creixements de f, però la seva incidència a nivell mundial és poca (Taula 2).
Finalment, veiem com hi ha uns quants països, dels més poblats del món, que tenen unes taxes de creixement positives en ambdós índexs. Crida l’atenció la Índia amb una acceleració de 0,411 i un nivell d’emissions de carboni similar al de Brasil, el que implica que serà un dels màxims emissors en el futur ja que incrementarà la seva petjada ecològica, ara molt baixa. El mateix s’aplica al Pakistan, amb unes taxes creixents també (0,0256) tot i que amb un impacte encara baix. La situació de Brasil és també de creixement de tots dos índexs i té una emissió de carboni a l’atmosfera molt gran (pel metà emès per la seva ramaderia).
Quin serà el creixement del futur?
Les projeccions de la ONU donen com a resultat un creixement de la població a finals del segle XXI de fins a 11000 milions d’humans a partir del qual la població s’estabilitzaria i despès començaria a baixar. La població creixerà sobretot a l’Àsia i a alguns països de l’Africa, com Nigèria. Aquest països es preveu que doblin el nombre d’habitants en un període de temps relativament curt. Però el creixement vindrà sobretot d’Àsia i Àfrica, de països ja molt poblats i amb taxes de fills/ dona encara molt altes, mentre Europa i Amèrica ja tenen taxes de fills per dona per sota de 1,5 (Taula 2). El creixement demogràfic és molt important i, si no es limita la població, les necessitats de cultivar per alimentar-la faran que hi hagi una crisi encara més gran de la Biodiversitat (Crist et al., 2017)
Hi ha països que, per la seva situació econòmica i per les limitacions que imposen a la immigració, tenen ja una disminució de la població i la tindran encara més gran en el futur. L’exemple paradigmàtic és el Japó. Diferents països seguiran el passos del Japó, i alguns factors poden encara reduir-la més. Per exemple, amb la COVID-19, la taxa de naixements a Espanya va disminuir el 20 % i ja era de les més baixes del món (1,32). Altres autors han estudiat les pautes de creixement de la població i han conclòs que, degut als canvis en l’actitud de les dones davant la maternitat induïts per diversos factors (per exemple les sèries de televisió), la població mundial no creixerà tant i que a finals del segle XXI serà d’uns 7 mil milions d’habitants, desprès d’haver crescut fins a 8,5 (Bricker & Ibbitson, 2019). O sigui que la r va de baixa i hi anirà cada vegada més, no està clar a quin ritme ni el moment en que s’estabilitzarà la població.
Pel que fa a l’energia, què podem predir?. Sembla que molts dels països aposten per una transició energètica, que no sempre inclou una disminució de l’ús de l’energia sinó un canvi a energies dites «més netes», de manera que hi ha fins i tot qui qüestiona si el «creixement verd» és possible (Hickel & Kallis, 2020). No queda clar, doncs, si hi haurà una disminució no sols de la f sinó del consum total d’energia de cada país, que és l’important. Ja hem dit abans que les prediccions no es poden fer a gaire llarg termini, totes les que es fan ara tenen un gran grau d’incertesa (variància en deia Margalef); la humanitat s’enfronta a escenaris demogràfics i de consum d’energia molt dispars i problemàtics, que inclouen l’escassetat d’energia i recursos. Els canvis futurs depenen molt de factors socials i polítics que de vegades poden fer caminar el món en direccions que no estaven previstes.
És incomplerta l’aproximació que feia Margalef fa 51 anys?
Fa 50 anys no existia ni el concepte de petjada ecològica, ni es coneixien prou bé les emissions de carboni a l’atmosfera o altres indicadors de canvi global que es fan servir ara com a descriptors dels canvis ambientals, com els de la Taula 2. Faria servir Margalef ara només l’energia i la demografia o buscaria altres indicadors? En el seu article de 1971, Margalef anava més enllà de la seva formula demofòrica i va explorar també l’impacte ambiental. Deia: «Més enllà de la qüestió demogràfica tenim l’explotació o la conservació de la natura, base de la nostra subsistència«. Parlava també de mobilitat i com això augmenta el consum d’energia externa. També de poŀlució o de les desigualtats produïdes per l’explotació de la natura, i de com l’ecologia... «..resulta un si és no és incòmoda quan no es limita a recomanar directament que hom no embruti massa els rius o com seria de convenient de no talar un bosc bonic», paraules altra volta ben actuals. I sentencia: «cada vegada estem més lligats els uns als altres i cada vegada la interacció entre home i natura es fa a una escala més amplia, és a dir, cada vegada seria més difícil superar els nostres disbarats coŀlectius». Sort que no era profeta.
Quan el món sembla encaminar-se a una aposta per fer servir sobretot energies renovables i anar abandonant els combustibles fòssils, potser podríem pensar que en el futur les emissions de carboni a l’atmosfera seran menors (amb o
229 Butlletí ICHN 85 (4), 2021 NOTA BREU L'HOME PART DE LA BIOSFERA: 50 ANYS D'UN ARTICLE SEMINAL DE RAMÓN MARGALEF
sense descens de l’energia usada). També, que la disminució d’aquestes emissions pot fer que es reverteixi l’increment de temperatura de l’atmosfera i tots els efectes associats (per tant potser no caldria reduir la f de Margalef). Es tractaria de mantenir la despesa d’energia individual (però renovable) alhora que el creixement econòmic mesurat com a increment del PIB, o sigui no caldria decreixement energètic i es podria seguir amb el creixement econòmic. Serà això possible?. La relació entre PIB i despesa d’energia (Kw-h/€) és també un valor interessant. Hi ha exemples de països amb baixa eficiència, com Turkmenistan, que podrien millorar, però Suïssa o Singapur, amb valors Kw-h/€ ja baixos, ja no poden disminuir gaire més aquesta eficiència i en canvi tenen altes emissions per càpita. N’hi haurà prou amb millorar l’eficiència o cal disminuir el consum energètic?. Tot fa preveure que sense la disminució del consum total d’energia (sigui de l’origen que sigui) no es revertirà el canvi ambiental que l’home està produint en la Biosfera.
Les emissions de CO2, o la petjada ecològica (les hectàrees de planeta que es necessiten per persona), o el dia en que a cada país se li acaba la superfície necessària per absorbir el seu impacte, són indicadors interessants (Taula 2). Tots mostren que no n’hi ha prou en baixar l‘acceleració (r+f) ni tampoc en millorar el valor de Kw-h/€. Disminuir l’acceleració que proporciona la (r+f) de l’equació de Margalef no és suficient i cal baixar tant la població com la despesa d’energia si volem un planeta sostenible, o sigui que cal un decreixement de tots els indicadors. El cotxe elèctric, l’ús de noves fonts d’energia o la crisi dels materials seguirà provocant un impacte ambiental enemic de la Biodiversitat i del funcionament de la Biosfera tal com el coneixem ara. Voler incrementar de forma ràpida la producció d’energies renovables, pot tenir altres impactes ambientals (que fan que molta gent s’oposi a nous parcs eòlics o solars), i per tant que els objectius de producció d’energies renovables no siguin assolits. En tot cas, tot i que l’emissió de gasos d’efecte hivernacle disminueixi, sembla que el consum d’energia seguirà alt i per tant també l’impacte ambiental. I seguirem invertint la «topologia del paisatge» (més urbanització i carreteres) el que, segons Margalef, és tant o més preocupant que el canvi climàtic. No sembla que les solucions actuals siguin les adequades per evitar que la Biosfera arribi al seu «punt crític» i ja no torni a ser mai més la que nosaltres vàrem conèixer.
A modus d’epíleg
En resum, l’aproximació de Margalef crec que encara és molt vàlida i que disminuir la població i gastar menys energia, sigui la que sigui, és el camí cap a la sostenibilitat. Les paraules de Margalef fa 50 anys són encara ben vàlides i ens recorden altra vegada la gran personalitat científica i humana d’algú que va ser un mestre per a molts, i un exemple com a persona. Li havia sentit a dir moltes vegades, quan anava a fer alguna xerrada a una escola o una llar d’avis, que anava
a fer «apostolat». Els que venim darrera també hem mirat de fer una mica d’apòstols, no sé si alguna vegada ho hem fet com Sant Pere i altres com Sant Pau, en tot cas hem intentat seguir els passos del nostre mestre per entendre, conservar i restaurar la Nostra Biosfera, en el meu cas sobretot els ecosistemes aquàtics.
Bibliografia
Bricker, D. & Ibbitson, J. 2019. El planeta vacío. Penguin Random House. Barcelona. 315 p. Crist, E., Camilo, M. & Engelman, R. 2017. The interaction of human population, food production and biodiversity protection. Science, 356: 260-264.
Crutzen, P. J. & Stoermer, E. F. 2000. The ‘Anthropocene. Global Change Newsletter, 41: 17-18.
IPCC, 2021. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [MassonDelmotte, V., P. Zhai, A. Pirani, S. L. Connors, C. Péan, S. Berger, N. Caud, Y. Chen, L. Goldfarb, M. I. Gomis, M. Huang, K. Leitzell, E. Lonnoy, J. B. R. Matthews, T. K. Maycock, T. Waterfield, O. Yelekçi, R. Yu and B. Zhou (eds.)]. Cambridge University Press. In Press.
Hickel, J. & Kallis, G. 2020. Is green growth possible?. New Political Economy, 25: 469-486.
IPBES 2019. Global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. E. S. Brondizio, J. Settele, S. Díaz, and H. T. Ngo (editors). IPBES secretariat, Bonn, Germany. 1148 pages. [Accessible a: https:// doi.org/10.5281/zenodo.3831673].
Margalef, R. 1971. L’Home part de la biosfera i objecte i estudi de l’Ecologia. Treballs Societat Catalana de Biologia, 30: 13-25.
Margalef, R. 1976. Bases ecològiques per a una gestió de la natura P. 25-64. In: Folch, R. (ed.). Natura, ús o abús? Llibre Blanc de la Gestió de la natura als Països catalans. Institució catalana d’Història natural/Barcino. Barcelona. 442 p.
Margalef, R. 1985. L’Ecologia. Servei del Medi ambient, Diputació de Barcelona. Barcelona. 126 pp.
Margalef, R. 1989. Introducció al coneixement de la biosfera. P. 15-25. In: J. Terradas, N. Prat. A. Escarré & R. Margalef (eds.) Història natural dels Països catalans, 14. Sistemes naturals. Folch, R. (Dir.). 500 p.
Margalef, R. 1997. Our Biosphere. Excellence in ecology. 10. Ecology Institute. Oldendorf/Luhe. 176 p.
Margalef, R. 2003. De com la civilització modifica l’entorn i accelera la dinàmica d’una evolució global amb inversió de la topologia original dels espais continentals humanitzats. In: Marrasé, C., Llebot, J.E. (Eds.). Un tast de canvi global. Treballs de la Societat Catalana de Biologia: 54: 9-12.
ONU 2021. Desafíos globales: Población. Disponible en: https:// www.un.org/es/global-issues/population [Data de Revisió: 1 desembre 2021]
Tello, E. 2013. La transformació històrica del paisatge entre l’economia, l’ecologia i la història: podem posar a prova la hipòtesi de Margalef?. Treballs de la Societat Catalana de Geografia, 75: 195-221.
230 Butlletí ICHN 85 (4), 2021
NOTA BREU
N. PRAT FORNELLS
NORMES DE PUBLICACIÓ DEL «BUTLLETÍ DE LA INSTITUCIÓ CATALANA D’HISTÒRIA NATURAL»
Objectiu de la revista
El Butlletí de la Institució Catalana d’Història Natural publica principalment articles inèdits i notes curtes de qualsevol camp de la història natural (revisions i descripcions relatives a la gea, la flora i la fauna; aspectes funcionals dels sistemes naturals; treballs sobre gestió del patrimoni natural, etc.), però també articles d’opinió i aportacions relatives a l’ofici del naturalista, ressenyes de llibres i glosses de naturalistes i científics destacats.
Tramesa dels manuscrits
Només podran ser publicats els treballs i les notes quan l’autor o un dels autors sigui soci de la ICHN.
Els treballs seran enviats per correu electrònic (butlleti.ichn@ iec.cat) en format RTF, Word (doc, docx) o pdf al Redactor en Cap. En el cas de que es faci per altres vies el Comitè Editorial no es fa responsable dels retards que se’n derivin.
Els articles podran ser escrits en català, castellà, anglès, francès o qualsevol altra llengua culta d’alfabet llatí, que garanteixi una àmplia difusió, seguint la normativa que s’exposa a continuació. El número total de pàgines, incloent les taules i figures, no podrà passar de 40 en format DIN-A4; en cas de manuscrits amb un nombre superior de pàgines, el comitè editorial es reserva la decisió de publicar-los.
La submissió d’un manuscrit implicarà que el treball és original i que no ha estat enviat a cap altra revista. No s’acceptaran articles ja publicats o en premsa.
En el cas que no s’acompleixi aquesta normativa, el manuscrit serà retornat a l’autor.
Organització dels treballs
Els treballs estaran organitzats de la següent forma (exceptuant casos especials):
• Títol
• Autors
Nom(s) i cognom(s) de l’autor(s), seguit a sota de la direcció(ns) professional(s) i correu electrònic(s). Si hi ha dos o més autors, aquests seran reconeguts amb superíndex.
• Autor per a la correspondència (només quan hi ha més d’un autor)
Nom complet i correu electrònic
• Resums
Si el treball és en català o anglès hi haurà un resum en català i un altre en anglès (màxim 250 paraules). En els altres casos hi haurà, a més dels resum esmentats, un resum en l’idioma del treball. El títol del treball encapçalarà en negreta el(s) resum(s) que no siguin escrits en l’idioma del treball
• Paraules clau (un màxim de 12), es coŀlocaran a sota de cada resum
• Introducció
• Materials i mètodes
• Resultats i discussió (separats o combinats)
• Conclusions (optatiu)
• Agraïments (si cal)
• Bibliografia
Estarà organitzada per ordre alfabètic. Els noms de les revistes no han de ser abreujats. No han d’aparèixer referències relacionades amb els noms dels autors de noms científics (veure més endavant la forma de citar-les).
• Figures i peus de figura, les taules i capçaleres de taula estaran integrats en el text o coŀlocats desprès de la bibliografia, en pàgina a part.
PUBLICATION RULES OF THE «BUTLLETÍ DE LA INSTITUCIÓ CATALANA D’HISTÒRIA NATURAL»
Scope of the journal
The Butlletí de la Institució Catalana d’Història Natural mainly publishes unpublished articles and short notes on any field of natural history (reviews and descriptions relating to Gaia, flora and fauna, functional aspects of natural systems, works on natural heritage management, etc.), but also opinion articles and contributions about the work of naturalists, book reviews and commentaries on the works of leading scientists and naturalists.
Submission of manuscripts
Only papers or notes by authors who are members of the ICHN will be published.
The manuscripts must be sent by e-mail (butlleti.ichn@iec.cat) in RTF format, Word (doc, docx) or pdf to the Editor in Chief. If manuscripts are sent by other means, the Editorial Board is not responsible for any delays that might occur.
The articles should be written in Catalan, Spanish, English, French or any other language that employs a Latin alphabet to ensure wide diffusion. The rules concerning submissions are set forth below. The total number of pages (including tables and figures) should not be greater than 40 DIN-A4; if there are more pages, the editorial board reserves the right to decide whether to publish them or not.
The submission of a manuscript implies that the work is original and has not been submitted to any other journal. The journal will not accept any article that has already been published or is in press.
If this rule is not fulfilled, the manuscript will be returned to the author.
Organization of the work
The work should be organized as follows (except in special cases):
• Title
• Authors
Name(s) and surname(s) of author(s), professional address(es) and e-mail(s). If there are two or more authors, author references will be represented in superscript.
• Corresponding author (only one if there is more than one author)
Full name and e-mail
• Abstract
If the work is in Catalan or English, abstracts should be written in Catalan and in English (maximum 250 words). In other cases, there should also be an abstract in the original language of the work. The title of the manuscript in bold will be given as a headline to the abstract(s) that are not written in the language used in the manuscript.
• Key words (max. 12), below the abstract.
• Introduction
• Material and Methods
• Results and Discussion (separate or combined)
• Conclusions (optional)
• Acknowledgements (if necessary)
• Bibliography
Organized in alphabetical order. The names of journals should not be abbreviated. References to the names of the authors of scientific names are not necessary (see below how to cite them).
• Figures and figure captions, tables and table captions should be integrated into the text or placed after the bibliography on a separate page.
231 Butlletí ICHN 85 (1-4), 2021
Les notes estaran organitzades de la següent forma:
• Títol (el primer títol amb l’idioma de la nota i el segon en anglès, si l’anglès és l’idioma de la nota el segon títol serà en català).
• Text (sense els subapartats presents en els treballs) amb les figures (si n’hi ha) incloses en el text.
• Agraïments (si cal)
• Bibliografia (veure l’apartat anterior)
• Autors (veure l’apartat anterior)
Les notes mai podran superar les 5 pàgines, figures incloses.
Format principal dels treballs i notes:
1) Seran escrits en DIN-A4, amb els marges esquerre i dret del text: 2,5 cm; marge superior: 3,0 cm, inferior: 2,5 cm.
2) Distància entre línies: 1,5.
3) El text definitiu, previ a l’acceptació definitiva del manuscrit, ha d’estar escrit preferentment amb RFT, tot i que també s’accepten formats Word
4) Font del títol: Times New Roman o Times, grandària de la lletra 14 pt, en negreta
5) Font del text: Times New Roman o Times, grandària de la lletra 10 pt. Tots els títols i subtítols han d’estar alineats a l’esquerra i en negreta
6) No activar la divisió de paraules al text.
7) Els paràgrafs han d’estar justificats i sagnats a 0,8 cm. Utilitzat sangries; no utilitzar espais ni tabulacions.
8) Els noms científics de gènere, subgènere, espècie, subespècie varietat i els sintaxons (associacions vegetals) aniran en cursiva. Els noms dels taxons superiors (família, ordre...) aniran sense cursiva, amb la primera lletra en majúscula si el terme s’escriu en llatí.
9) Els noms dels autors de les espècies esmentades s’han d’escriure amb la mateixa lletra del text (lletra normal), sense abreujar. Només s’esmenta l’autor i any la primera vegada que s’anomena el taxó.
10) Utilitzar n. sp., n. gen., n. fam, etc, cada vegada que nous tàxons es mencionin, i n. comb. cada vegada que una nova combinació se citi.
11) Les referències que se citin al text s’han d’escriure amb la mateixa lletra del text. Exemples: Bunyol (2001), Bunyol (2001, 2002), (Bunyol, 2000; Torrat, 2002), (Bunyol & Torrat 2003, 2005), Bunyol (2001: 1; 2003: 4), Bunyol (2001: Fig. 2), Bunyol et al. (2003) (Bunyol et al., 2006) segons convingui.
12) No numerar les pàgines
13) Les figures (dibuixos i fotos) han de ser referenciades en el text com «Fig. X» (si n’hi ha una) o «Figs. X-XX» (en el cas de que n’hi hagi més d’una) amb un punt desprès de la «g» o «s» respectivament (si la redacció és en anglès aleshores sense punt desprès de la «g» o «s»). Les taules com «Taula X» o «Taules X-XX»
14) Si una figura conté més d’una imatge, aquestes s’hauran d’identificar amb lletres minúscules (a, b, c, ...) a la mateixa figura, de manera que en el text quedi referenciada la «Fig. 1a» o les «Figs. 1a-b», segons correspongui.
Taules i figures
Les iŀlustracions originals s’enviaran a l’editor quan el manuscrit hagi estat revisat, en fitxers independents del text juntament amb el manuscrit revisat.
- Els dibuixos en gama de grisos, les figures i/o plànols hauran de tenir una resolució de 300 dpi en format TIF o JPEG d’alta qualitat.
Cada figura pot incloure una escala mètrica, sense multiplicador.
Bibliografia
Les referències seran sagnades a model francès (a partir de la segona línia) a 1cm
Notes should be organized as follows:
• Title (The first title in the note language, the second in English. If English is the language of the note, the second title should be in Catalan)
• Text (with no subsections) and figures (if any) incorporated into the text.
• Acknowledgements (if necessary)
• Bibliography (see the previous section)
• Authors (see the previous section)
Notes should never exceed 5 pages, including figures.
Basic layout of manuscripts and notes:
1) Written on A4 paper, with left- and right-hand margins to the text: 2.5 cm; top margin: 3.0 cm; bottom margin: 2.5 cm.
2) Distance between lines: 1.5.
3) The final text, prior to final acceptance of the manuscript, should be written preferably in RFT, although a Word format is also acceptable.
4) Source title: Times New Roman or Times font size 14 pt., bold.
5) Text Font: Times New Roman or Times font size 10pt. All headings and subheadings should be left aligned and in bold.
6) The hyphenation text option should not be used.
7) Paragraphs should be justified and indented by 0.8 cm. If indentation is used, do not use spaces or tabs.
8) The scientific names of genera, sub-genera, species, subspecies, varieties and syntaxa (plant associations) should be written in italics. The names of higher taxa (family, order, etc. ...) should not be in italics and should have the first letter capitalized if the word is written in Latin.
9) The names of the authors of the species mentioned must be written in the same text font (regular font), and not abbreviated. The descriptor of the species and the year should be given the first time the name of the taxon is cited.
10) Use n. sp., n. gen., n. fam., etc., each time a new taxon is mentioned and n. comb. each time a new combination is cited.
11) References cited in the text should be written in the same style as the text. Examples: Bunyol (2001), Bunyol (2001, 2002), (Bunyol, 2000; Torrat, 2002), (Bunyol & Torrat 2003, 2005), Bunyol (2001: 1; 2003: 4), Bunyol (2001: Fig. 2), Bunyol et al. (2003) (Bunyol et al., 2006).
12) Pages should not be numbered.
13) Figures (drawings and photographs) should be referenced in the text as «Fig. X» (if any) or «Figs X-XX» (if there is more than one) with a full-stop after the «g» or «s» (if the text is in English there is no need for a full-stop after the «g» or «s»). Tables should be mentioned as follows: «Table X» or «X-XX tables».
14) Multiple images in a figure should be identified with lower-case letters (a, b, c, etc. ...) in the figure and referenced in the text as «Fig. 1a» or «Figs 1a-b», as appropriate.
Tables and figures
Once the manuscript has been reviewed, the original artwork must be sent to the editor in separate text files along with the revised manuscript.
- Drawings in shades of grey; figures and/or plans should have a resolution of 300 dpi and be in either JPEG or TIF high-quality format.
Each figure may include a metric scale without a multiplier.
Bibliography
References will be indented 1 cm as per the French model (from the second line onwards).
232 Butlletí ICHN 85 (1-4), 2021
Se citarà de la següent forma segons el cas:
Article de revista
(AUTOR Any. Títol. Nom complet de la revista en cursiva, número: pàgines)
- un autor:
Viñolas, A. 2002. Nova aportació al coneixement dels anòbids de la península Ibèrica (Coleoptera: Anobiidae). Butlletí de la Institució Catalana d’Història Natural, 70: 73-77.
- més d’un autor:
Pujade-Villar, J., Hanson, P., & Melika, G. 2012. A new genus of oak gallwasp, Coffeikokkos Pujade-Villar & Melika, gen. n., with a description of a new species from Costa Rica (Hymenoptera, Cynipidae). ZooKeys, 168: 19–29.
Llibre complet
(AUTOR Any. Títol en cursiva. Editorial o entitat responsable de la publicació. Lloc de la impressió. Número de pàgines).
Bolòs, A. 1950. Vegetación de las comarcas barcelonesas, descripción geobotánica y catálogo florístico. Instituto Español de Estudios Mediterráneos. Barcelona. 579 p.
Capítol de llibre
(AUTOR Any. Títol del capítol en cursiva. Pàgines del capítol (P. XX-XX). In: Editor (ed.). Títol del llibre. Editorial. Ciutat. País. Número de pàgines del llibre.)
Montoya-Lerma, J. & Ferro, C. 1999. Flebótomos (Diptera: Psychodidae) de Colombia. P. 211-245. In: Amat, G.; Andrade-C., G.; Fernández, F. (eds.). Insectos de Colombia. Volumen II. Academia Colombiana de Ciencias Exactas, Físicas y Naturales. Colección Jorge Alvarez Lleras. No. 13. Editora Guadalupe Ltda. Bogotá. Colombia. 492 p.
Citacions d’ internet
(AUTOR/EDITOR. Any. Títol. “Disponible a:” URL”, [data de consulta escrit com dia/mes/any, sense comes]
Font, X. 2013. Mòdul Flora i Vegetació. Banc de Dades de Biodiversitat de Catalunya. Generalitat de Catalunya i Universitat de Barcelona. Disponible en: http://biodiver.bio.ub.es/biocat/homepage.html [Data de consulta: 5 juny 2012].
Revisió dels manuscrits
Els treballs seran revisats per dos assessors escollits per la Comissió de Publicacions d’acord amb la temàtica de l’article. Un cop el manuscrit hagi estat revisat es comunicarà a l’autor, si es dóna el cas, les modificacions necessàries perquè sigui acceptat definitivament. L’autor podrà fer-hi les esmenes que consideri oportunes i la Comissió de Publicacions en decidirà l’acceptació final o el rebuig si no es modifica. En cas de desavinença dels autors o en el cas de que el treball no sigui acceptat, si no es modifica (de forma justificada), es podrà demanar la retirada de l’article.
Publicació
Un cop el treball hagi estat definitivament acceptat es publicarà online en format pdf al més aviat possible a l’apartat Publicacions, del web de la ICHN, ja paginats i amb el seu corresponent ISSN (online). A final d’any, el volum querarà tancat i serà exclusivament on-line.
Les fotografies seran publicades a color.
Els autors rebran els pdf corresponents. No es faran separates en paper.
References will be cited as follows:
Journal articles
(AUTHOR Year. Title. Complete name of the journal in italics, number: pages)
- one author:
Viñolas, A. 2002. Nova aportació al coneixement dels anòbids de la península Ibèrica (Coleoptera: Anobiidae). Butlletí de la Institució Catalana d’Història Natural, 70: 73-77.
- more than one author:
Pujade-Villar, J., Hanson, P., & Melika, G. 2012. A new genus of oak gallwasp, Coffeikokkos Pujade-Villar & Melika, gen. n., with a description of a new species from Costa Rica (Hymenoptera, Cynipidae). ZooKeys, 168: 19–29.
Complete book
(AUTHOR Year. Title in italics. Editorial or entity responsible for the publication. Place of printing. Number of pages).
Bolòs, A. 1950. Vegetación de las comarcas barcelonesas, descripción geobotánica y catálogo florístico. Instituto Español de Estudios Mediterráneos. Barcelona. 579 p.
Book chapter
(AUTHOR Year. Title of the chapter in italics. Pages of the chapter (P. XX-XX). In: Editor (ed.). Book title. Editorial. City. Country. Number of book pages.
Montoya-Lerma, J. & Ferro, C. 1999. Flebótomos (Diptera: Psychodidae) de Colombia. P. 211-245. In: Amat, G.; Andrade-C., G.; Fernández, F. (eds.). Insectos de Colombia. Volumen II. Academia Colombiana de Ciencias Exactas, Físicas y Naturales. Colección Jorge Alvarez Lleras. No. 13. Editora Guadalupe Ltda. Bogotá. Colombia. 492 p.
Internet citations
(AUTHOR/EDITOR. Year. Title. Available at: ‘URL’, [Consulted on day/month/year, no commas].
Font, X. 2013. Mòdul Flora i Vegetació. Banc de Dades de Biodiversitat de Catalunya. Generalitat de Catalunya i Universitat de Barcelona. Disponible en: http://biodiver.bio.ub.es/biocat/homepage.html [Data de consulta: 5 juny 2012].
Review of manuscripts
Papers will be reviewed by two reviewers chosen by the Publication Committee in accordance with the theme of the article.
Once the manuscript has been reviewed, the author will be informed, if applicable, of the modifications needed for definitive acceptance. The author should make the necessary amendments and the Publication Committee will decide whether the modified manuscript is to be accepted or rejected. In case of disagreement or if the work is not accepted if not modified (with justification), a request to removal the article may be made.
Publication
Once the work has been definitively accepted, it will be published online in pdf format as soon as possible in the Publications section of the ICHN website, already paginated and with its corresponding ISSN (online). At the end of the year, the volume will be closed and will be exclusively online.
Photographs will be published in color.
Authors will receive the corresponding pdf. There will be no reprints.
233 Butlletí ICHN 85 (1-4), 2021
GEA, FLORA ET FAUNA
Artur LLuent & Antonio Gómez-BoLeA
Distribució de macrolíquens a Catalunya amb noves dades corològiques sobre líquens protegits
JorDi Bou mAnoBens & LLuís ViLAr sAis
JorDi ComAs Dos
Pere AYmeriCH
Notes florístiques de les conques altes dels rius Segre i Llobregat. V Floristic data of the upper Segre and Llobregat basins. V 41
Pere PoNS, raFaeL CarboNeLL FoNt, Martí FraNCh, JoSeP M. baS, DaNieL eSPeJo Fraga, FerraN FoNteLLeS, DAViD FunosAs, mArC FuseLLAs, roGer PuiG-Gironès, CArLes toBeLLA & mArC FrAnCH Diversitat, distribució i fenologia de les cigales (Hemiptera: Cicadidae) a Catalunya (ne Península iberica)
Diversity, distribution and phenology of cicadas (Hemiptera: Cicadidae) in Catalonia (ne iberian Peninsula)
eDuArD ViVes & JorDi ComAs
un nou gènere i espècie de molopina hipogeu del Pirineu Català (Alt urgell, Lleida) (Coleoptera: Carabidae: Pterostichini)
A new genus and new species of hypogeum molopina from Catalan Pyrenean area (L’Alt urgell, Lleida)
AmADor ViñoLAs, José iGnACio reCALDe irurzun & JoseP muñoz-BAtet revisión de la familia salpingidae
JoseP
JorDi ViLa, oLga MorozoVa, aNNa FeDoSoVa, tàNia JiMéNez-PaLoMar & XaVier LLiMoNa
al coneixement de la flora vascular del sector oriental del massís del Montseny (Catalunya, Ne de la península ibèrica)
Contributions to the knowledge of the vascular flora of the eastern sector of the Montseny massif (Catalonia, Ne iberian Peninsula)
Aitor mArtínez-romero, Antoni riBes esCoLà (†), BertA CABALLero-LóPez & JuLi PuJADe-ViLLAr gales i organismes cecidògens de Ponent de la coŀlecció antoni ribes
Galls and cecidogenic organisms of Ponent from the Antoni ribes collection 209
NOTES BREUS
ALBert tArrAGó
Actualització de la informació sobre la població de Saponaria glutinosa (Caryophyllaceae) a l’Alt urgell (Catalunya)
new information about Saponaria glutinosa (Caryophyllaceae) in Alt urgell (Catalonia)
3
Distribution of macrolichens in Catalonia with new chorological data on protected lichens
L’associació
trixaginis-Scirpoidetum holoschoeni a Catalunya the Bartsia trixaginis-Scirpoidetum holoschoeni association in Catalonia 19
Bartsia
noves espècies del
Parvospeonomus Bellés & escolà, 1977 (Coleoptera: Leiodidae:
two new species of the genus Parvospeonomus Bellés & escolà, 1977 (Coleoptera: Leiodidae: Leptodirini) 29
gènere
Leptodirini)
59
(Coleoptera: Carabidae: Pterostichini) 77
Leach, 1815 en el área iberobalear (Coleoptera) review of the family salpingidae Leach, 1815 from the iberian-Balearic area (Coleoptera) 91
romero & Pere nAVArro-rosinés sobre la presència d’Hydropunctaria scabra a la península ibèrica, i notes sobre altres líquens aquàtics de Les Planes de son i mata de València About the presence of Hydropunctaria scabra in the iberian Peninsula, and notes of other aquatic lichens of Les Planes de son i mata de Valencia 119 AmADor ViñoLAs & JoseP muñoz-BAtet nuevos datos sistemáticos y de distribución de Xyletinus (Calypterus) bucephalus s. lat. (Coleoptera: Ptinidae: Xyletininae) new systematic and distribution data for Xyletinus (Calypterus) bucephalus s. lat. (Coleoptera: Ptinidae: Xyletininae) 131
PuJADe-ViLLAr & PAuL HAnson Neuroterus titou n. sp. from Costa rica (Cynipidae: Cynipini) Neuroterus titou n. sp. de Costa rica (Cynipidae: Cynipini) 139 Pere AYmeriCH & LLorenç sáez aportacions a la flora aŀlòctona catalana Notes about alien flora in Catalonia 151
CristiAn
JuLi
Gesti PeriCH & LLuís
aportacions al coneixement de la flora de les guilleries orientals i àrees properes (nord-est de Catalunya) – iii Contributions to the knowledge of the flora of the eastern guilleries and nearby areas (northeastern Catalonia) – iii 163
ViLAr sAis
Principals resultats de
russo-catalana als Pirineus catalans occidentals: prats alpins i subalpins i boscos subalpins main results of the russian-Catalan mycological survey, in the western Catalan Pyrenees: alpine and subalpine belt 171 Antoni mAYorAL Arqué sobre la gairebé absència de Rosmarinus officinalis (Lamiaceae) al Pla d’urgell i algunes anomalies fenològiques interessants About the near absence of Rosmarinus officinalis (Lamiaceae) in Pla d’urgell and some interesting phenological anomalies 191
BArnoLA eCHenique,
&
aportacions
la campanya micològica
Pere
JoseP mAriA PAnAreDA CLoPés
JoseP Gesti PeriCH
199
íNDeX
35
JuLi PuJaDe-ViLLar, aMaDor ViñoLaS, JoSeP Muñoz-batet, DeNiS J. brotherS & CeLSo o azeVeDo
Sclerodermus versus Scleroderma (Hymenoptera: Bethylidae)
Sclerodermus
JAn tomàs, ADrià mirALLes-núñez & ADrián neCoeCHeA
La xinxa de l’eucaliptus, Thaumastocoris peregrinus Carpintero & Dellapé, nou heteròpter exòtic per a Catalunya (Hemiptera: thaumastocoridae)
the bronze bug, Thaumastocoris peregrinus Carpintero & Dellapé, new exotic true bug for Catalonia (Hemiptera: thaumastocoridae)
mArtA GouLA, rAFAeL CArBoneLL, eLenA LóPez GALLeGo & JAn tomàs
Alloeorhynchus (Alloeorhynchus) flavipes (Fieber, 1836), espècie reconfirmada per a la fauna ibèrica (hemiptera: heteroptera; Nabidae)
Alloeorhynchus (Alloeorhynchus) flavipes (Fieber, 1836), reconfirmed species to the iberian fauna (hemiptera: heteroptera: Nabidae)
enriC BALLesteros & Boris WeitzmAnn on the presence of a small population of Sargassum trichocarpum J. agardh (Phaeophyceae: Fucales) in Catalonia (northwestern mediterranean) sobre la presència d’una petita població de Sargassum trichocarpum J. agardh (Phaeophyceae: Fucales) a Catalunya (mediterrània nord-occidental)
JoseP muñoz-BAtet, JoAquim soLer & AmADor ViñoLAs
Osmoderma eremita (Scopoli, 1763) al Paratge Natural d’interès Nacional de l’albera (Coleoptera: Scarabaeidae:
37
55
73
83
JuLi PuJaDe-ViLLar, JuaN M. VaNegaS-riCo & NoeL Mata-CaSaNoVa
vs. Scleroderma (Hymenoptera: Bethylidae)
Cetoniinae) amb notes sobre altres coleòpters saproxílics interessants Osmoderma eremita (Scopoli, 1763) at the «Paratge Natural d’interès Nacional de l’albera» (Coleoptera: Scarabaeidae: Cetoniinae) with notes on other interesting saproxylic beetles 87 LLuís ViLAr & LLorenç sáez Erigeron blakei Cabrera (asteraceae), an alien species confirmed for the iberian Peninsula Erigeron blakei Cabrera (asteraceae), espècie aŀlòctona confirmada a lapenínsula ibèrica 111 Genís PuiG & LLorenç sáez nova població de l’espècie amenaçada Asphodelus ramosus subsp. ramosus (Asphodelaceae) a Catalunya A new population of the threatened species Asphodelus ramosus subsp. ramosus (Asphodelaceae) in Catalonia 115 Pere AYmeriCH & ALBert tArrAGó noves dades poblacionals sobre Daphne alpina (thymelaeaceae) a Catalunya new populational data about Daphne alpina (thymelaeaceae) in Catalonia 125
Primer hospedador conocido de Aegilips clarimontis Kieffer, 1907(Figitidae: Anacharitinae) First known host of Aegilips clarimontis Kieffer, 1907 (Figitidae: Anacharitinae) 127 iGnAsi soriAno sobre la variabilitat de Pedicularis pyrenaica i la presència de Pedicularis mixta (orobanchaceae) als Pirineus orientals on the variability of Pedicularis pyrenaica and the presence of Pedicularis mixta (orobanchaceae) in the eastern Pyrenees 147 JorGe meDeros, ADrià mirALLes-núñez & eDuArD mArquès-morA Primeros registros de Nigrotipula nigra nigra (Linnaeus, 1758) para la Península ibérica (Diptera: tipulidae) First records of Nigrotipula nigra nigra (Linnaeus, 1758) (Diptera: tipulidae) for the iberian Peninsula 187 AmADor ViñoLAs & JoseP muñoz-BAtet sobre la distribución de especies de la familia Ciidae Leach, 1819 (Coleoptera) en la Península ibérica no contempladas en el nuevo Catálogo Paleártico on the distribution of species of the family Ciidae Leach, 1819 (Coleoptera) in the iberian Peninsula not included in the new Catalogue Palaearctic 225 nArCís PrAt ForneLLs L’home part de la biosfera: 50 anys d’un article seminal de ramón margalef man as part of the Biosphere: 50 years of an oustanding paper from ramon margalef 227
